1. Introduction
The cultivation of oil palm serves as a pivotal economic catalyst within the Indonesian context, making substantial contributions to the national economy by serving as a leading plantation commodity and a significant generator of foreign exchange. As attested by Alkusma et al. [
1], the relentless expansion of plantations and the burgeoning oil palm industry correlates with a heightened generation of waste materials, notably including substantial quantities of oil palm empty fruit bunches (OPEFB)—a form of solid waste. As reported by Yanti and Hutasuhut [
2], in the year 2018, up to 6.7 million tons of OPEFB were disposed of, raising environmental challenges due to pollution and landfill congestion.
OPEFB is classified as lignocellulosic biomass waste, representing a reservoir of substantial untapped potential. Its constitution includes cellulose (ranging from 24% to 65%), hemicellulose (comprising 21% to 34%), and lignin (constituting 14% to 31%) [
3]. Particularly noteworthy is its significant hemicellulose content, a complex polysaccharide polymer that includes glucose, mannose, arabinose, and xylose units, thereby conferring structural support to the cellular walls. Importantly, up to 82% of the hemicellulose in OPEFB can be converted into xylose, thereby serving as a pivotal precursor for xylitol production [
4].
Xylitol is a sugar alcohol used in several sectors, including the food, pharmaceutical, and health industries. It is commonly used in the manufacturing of confectioneries, chewing gum, carbonated beverages, and oral hygiene items. In a study conducted by Muryanto et al. [
5], the residue obtained after the extraction of xylitol, rich in cellulose, has the potential for conversion into bioethanol, a kind of liquid fuel energy. Additionally, residues of xylitol and ethanol production may contain partially degraded lignin, as well as incompletely converted hemicellulose and cellulose, serving as substrates for enzymatic reactions. Enzymes like cellulase, xylanase, and laccase are often derived from the metabolic processes of microorganisms [
6].
In the pretreatment phase, the primary objective is to eliminate lignin, reduce cellulose crystallinity, and convert OPEFB into pulp to enhance the feasibility of hydrolysis and fermentation. This is achieved by subjecting the polysaccharide crystals to a heating process at a temperature of 120 °C [
7]. During hydrolysis, hemicellulose is degraded, facilitating the extraction of fibers, and polysaccharide chains are broken down into monosaccharides [
8]. The hydrolysate is then used in the fermentation phase, where xylitol is produced by the yeast
Meyerozyma caribbica. This particular yeast strain is recognized for its resilience to inhibitory substances, making it an ecologically sustainable and cost-effective choice for this process. The achievement of effective xylitol bioconversion through fermentation is contingent upon many parameters, including temperature, pH, aeration conditions, substrate concentration, and the existence of other molecules apart from xylose, such as glucose. In addition, the xylose solution undergoes a conversion process, resulting in the formation of the xylitol solution [
9]. The adequate preparation of the medium is crucial for achieving good fermentation of xylitol. The use of different compositions of the medium has a significant impact on the resulting products.
On the other hand, the solid waste produced during the process of xylitol manufacture from OPEFB is significant due to its substantial cellulose content, making it a promising feedstock for bioethanol production within a biorefinery framework. Enzymatic hydrolysis can degrade cellulose polymers into glucose, enabling further processing of residual solid waste. Compared to acid hydrolysis, enzymatic hydrolysis generates more glucose, resulting in more significant bioethanol production [
10].
While previous studies have mostly focused on bioethanol production, this study adopts a novel perspective by examining bioethanol generation using varying quantities of cost-effective and readily available fermentation media [
11]. The objective is to maximize the extraction of ethanol from a cost-effective substrate. Based on the findings of Mardawati et al. [
4], it is imperative that the materials used in the process satisfy the fundamental nutritional demands necessary for the proliferation of microorganisms. These needs include key components such as carbon, nitrogen, non-metallic constituents (specifically sulfur and phosphorus), metallic elements including calcium, zinc, sodium, potassium, copper, manganese, magnesium, and iron, as well as vitamins, water, and energy.
This study employs a two-stage enzymatic hydrolysis technique, focusing on using OPEFB waste for bioethanol synthesis and xylitol production. Both steps of hydrolysis require the use of hemicellulase enzymes to catalyze the breakdown of hemicellulose into xylose, serving as a substrate for xylitol fermentation. The cellulose-rich xylitol residue is subjected to further hydrolysis to produce glucose hydrolysate that may be used for ethanol production. The resulting solid waste from the process of hydrolysate separation is used as a substrate for lignocellulosic enzyme synthesis. This study demonstrates the use of the OPEFB biorefinery for the combined synthesis of xylitol and bioethanol, with subsequent fermentation using different medium compositions.
Enzyme synthesis can be accomplished through fermentation by microbial organisms, often bacteria or fungi [
12]. It has been shown that fungi exhibit a greater hydrolysis rate in comparison to yeast and bacteria. They are capable of producing a diverse range of hydrolytic and oxidative enzymes that have the ability to break down the components of lignocellulosic materials. The selection of
Aspergillus niger, a filamentous fungus known for its production of cellulase, xylanase, and laccase enzymes, was based on its favorable performance and cost-effectiveness compared to other choices [
13]. The development of fungi is influenced by several parameters, including substrate concentration, nutrient supply, aeration, pH, temperature, incubation conditions, moisture content, and fermentation duration [
14].
Microorganisms play an active role in fermentation and naturally reproduce in environments conducive to their development [
15]. The present study used the submerged fermentation technique to produce the three enzymes. The methodology used a liquid substrate, with the addition and replacement of nutrients conducted in a continuous manner within the submerged fermentation medium. This environment is particularly appropriate for microorganisms, including fungi, that prefer elevated levels of moisture [
16].
The primary aim of this research was to evaluate the use of a fermentation medium in conjunction with a two-stage enzymatic hydrolysis procedure for the production of xylitol and bioethanol. Furthermore, an additional objective for this research was to assess the enzymatic activity of cellulase, xylanase, and laccase enzymes produced by Aspergillus niger during cultivation on an integrated OPEFB substrate. The enzyme evaluation was conducted by taking into account key variables, including activity, protein content, specific activity, and enzyme mass.
2. Materials and Methods
2.1. Materials Preparation and Source
All components of the OPEFB parts were cleaned under running water and subjected to drying at 60 °C in an oven for a period of 24 h. The dried material was subsequently fragmented into finer particles using a disc mill and sorted via a sieve with a mesh size ranging from 60 to 80 mesh or 0.25 to 0.177 mm size. The prepared materials were stored in air-tight bags at ambient temperature until they were used.
Enzymatic hydrolysis was performed using the Cellic HTec2 and Cellic CTec2 enzymes sourced from Novozymes in Copenhagen, Denmark. The activity of these enzymes was 75 IU/mL and 130 FPU/mL, respectively. In this study, Meyerozyma caribbica Y67 from the Indonesia Culture Collection (InaCC)-BRIN, Indonesia, was used to ferment xylitol. While Candida sp. Unpad CC Y26 was used to make ethanol derived from environmentally exposed OPEFB. Additional chemicals were purchased from Sigma Aldrich in St. Louis, Missouri, and Merck in Singapore. All chemicals were of laboratory grade and used immediately upon receipt. All solutions were prepared with 18.2 MΩ cm Milli-Q water (Millipore, St. Louis, MO, USA).
2.2. Pretreatment of Oil Palm Empty Fruit Bunches
A preliminary study was undertaken to identify the optimal pretreatment method for OPEFB. The experimental investigation involved the evaluation of different pretreatment methods, specifically acid and alkaline catalysts, with variations in the duration of the pretreatment process for each catalyst. The OPEFB flour was dissolved in either 4% NaOH or 4% H
2SO
4 solutions after being weighed on an analytical scale. Referring to the study by Sari et al. [
17], the materials were mixed in an Erlenmeyer flask using a solid-to-liquid ratio of 1:20 (b/v). This means that 20 g of OPEFB flour was added to 400 mL of solution. The mixture was processed in an autoclave at 121 °C for periods of 15, 30, and 60 min. The optimal pretreatment methods would be selected based on lignin reduction, hemicellulose, and cellulose retention, as well as process duration.
2.3. Enzymatic Hydrolysis of Pretreated Oil Palm Empty Fruit Bunches Using Cellic Htech Enzyme
The pretreated OPEFB powder was sterilized by immersing 10 g in 100 mL of pH 5 acetate buffer in an Erlenmeyer flask and autoclaving it for 15 min at 121 °C. Hydrolysis was carried out in a shaker incubator (N-Biotek, Seoul, South Korea) at 60 °C and 150 rpm for 96 h using 38.5 I.U./g of biomass Cellic HTec. Following hydrolysis, the liquid fraction was separated through centrifugation at 6000 rpm for 20 min. Sampling was performed every 24 h. The solid residue was oven-dried at 105 °C oven for 24 h. The dried solid residue was employed as a substrate for the synthesis of ethanol.
Equations for calculating theoretical xylose and glucose concentrations (g/L) in the feedstocks are defined in Equations (1) and (2), respectively, as follows:
Theoretical xylose concentration (g/L):
Theoretical glucose concentration (g/L):
2.4. Xylitol Fermentation of Oil Palm Empty Fruit Bunches Hydrolysate by M. caribbica
Inoculum preparation started by sterilizing 50 mL of potato dextrose broth (PDB) at 121 °C in an autoclave for 15 min. Following sterilization, the sterile medium solution was inoculated with a single loopful of Meyerozyma caribbica cells and incubated at 30 °C for 48 h, with a shaking speed of 200 rpm in a shaker incubator. Subsequently, 5% of this cultivated inoculum was mixed with 200 mL of fresh PDB medium and incubated again in a shaker incubator at 30 °C for 48 h at 200 rpm. The mixture was then centrifuged at 6000 rpm for 12 min to separate the cell pellet from the medium. The cell pellet was subsequently homogenized in 50 mL of the desired fermentation medium.
This study examined three different xylitol fermentation media. The first medium, labeled Medium A, has been employed in prior research for xylitol fermentation, enriched with a variety of minerals and vitamins [
18]. The composition of this medium was 9.438 g/L (NH
4)
2SO
4, 2.5 g/L KH
2PO
4, 0.05 g/L CaCl
2·2H
2O, 0.5 g/L MgSO
4·7H
2O, 0.5 g/L citric acid, 0.035 g/L FeSO
4·7H
2O, 0.0092 g/L MnSO
4·7H
2O, 0.011 g/L ZnSO
4·7H
2O, 0.001 g/L CuSO
4·7H
2O, 0.002 g/L CoCl
2·6H
2O, 0.0013 g/L Na
2CoO
4·2H
2O, 0.002 g/L H
3BO
3, 0.0035 g/L KI, 0.0005 g/L Al
2(SO
4)
3, 0.1 g/L myo-inositol, 0.02 g/L calcium-pantothenate, 0.005 g/L thiamine hydrochloride, 0.005 g/L pyridoxal hydrochloride, 0.005 g/L nicotine acid, 0.001 g/L aminobenzoic acid, and 0.0001 g/L d-biotin. The second medium, labeled Medium B, was formulated with 5 g/L yeast extract, 2 g/L KH
2PO
4, 2 g/L (NH
4)
2HPO
4, and 0.3 g/L MgSO
4·7H
2O. The third medium, referred to as Medium C, is an AB-mix, which is a commercial hydroponic fertilizer that contains macroelements, such as N, Ca, K, Mg, S, and P, as well as microelements, including Fe, Mn, Cu, B, Zn, and Mo.
2.5. Ethanol Production Using Semi-Simultaneous Saccharification and Fermentation (Semi-SSF) Method
The Semi-SSF technique was used to produce bioethanol from OPEFB residue. A 250 mL Erlenmeyer flask with a working volume of 100 mL was used. The OPEFB residue and cellulase enzyme concentrations in the total citrate buffer solution at pH 5 were 10% (w/v) and 65 FPU/gram biomass, respectively. Before enzyme addition, both biomass and buffer were sterilized. The saccharification process was conducted in the incubator shaker at 50 °C for 96 h at a shaking speed of 130 rpm.
For inoculum preparation, 50 mL of potato dextrose broth (PDB) medium was sterilized and mixed with one loopful of Candida sp. Unpad CC Y26 cells. The inoculum solution was incubated at 30 °C and 200 rpm for 48 h in a shaker incubator. Then, 5% of the prepared inoculum was mixed with 200 mL of fresh PDB medium and incubated under the same conditions. The cell pellet was separated from the media by centrifuging the inoculum at 6000 rpm for 12 min.
After 96 h of the hydrolysis period, neither filtration nor enzyme inactivation was carried out on the hydrolysate. A cell pellet of Candida sp. Unpad CC Y26 and a two-fold concentrated fermentation medium were then added to the hydrolysis Erlenmeyer flask. Fermentation was conducted at 30 °C, 150 rpm for an additional 96 h in an incubator shaker.
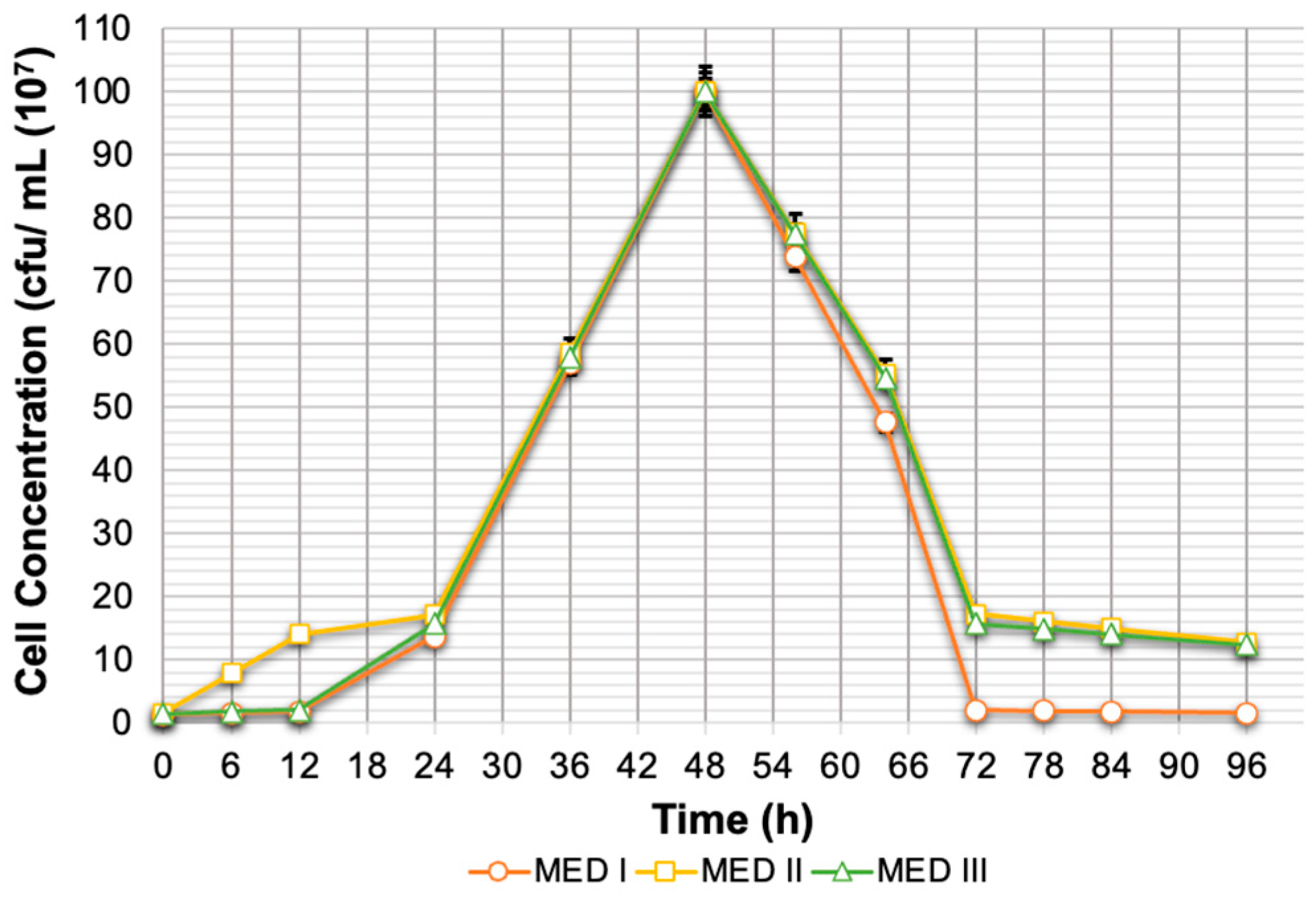
This section encompasses an investigation of three distinct types of xylitol fermentation media. The first medium (Medium 1) compositions were 1.8 g/L yeast extract, 0.9 g/L (NH4)2HPO4, and 0.6 g/L (NH2)2CO. The second medium (Medium 2) compositions were 4 g/L peptone, 3.6 g/L yeast extract, 2 g/L KH2PO4, 1 g/L MgSO4·7H2O, and 3 g/L (NH4)2SO4. The third medium (Medium 3) compositions were 5 g/L peptone, 3 g/L yeast extract, 1 g/L KH2PO4, 0.5 g/L MgSO4·7H2O, 0.11 g/L CaCl2, and 0.05 g/L ZnSO4.
2.6. Extracellular Enzymes Production by Aspergillus niger Unpad CC C42
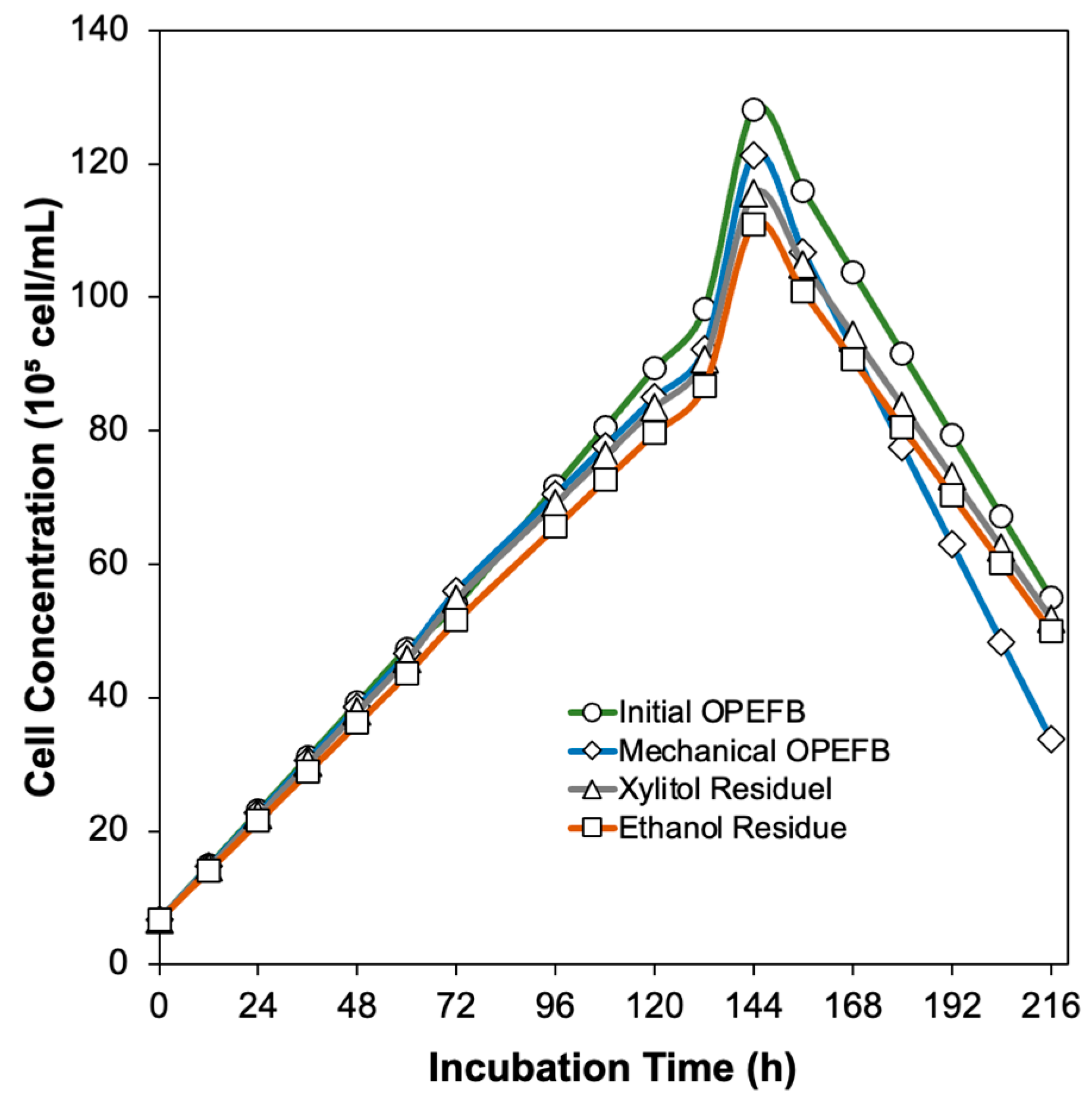
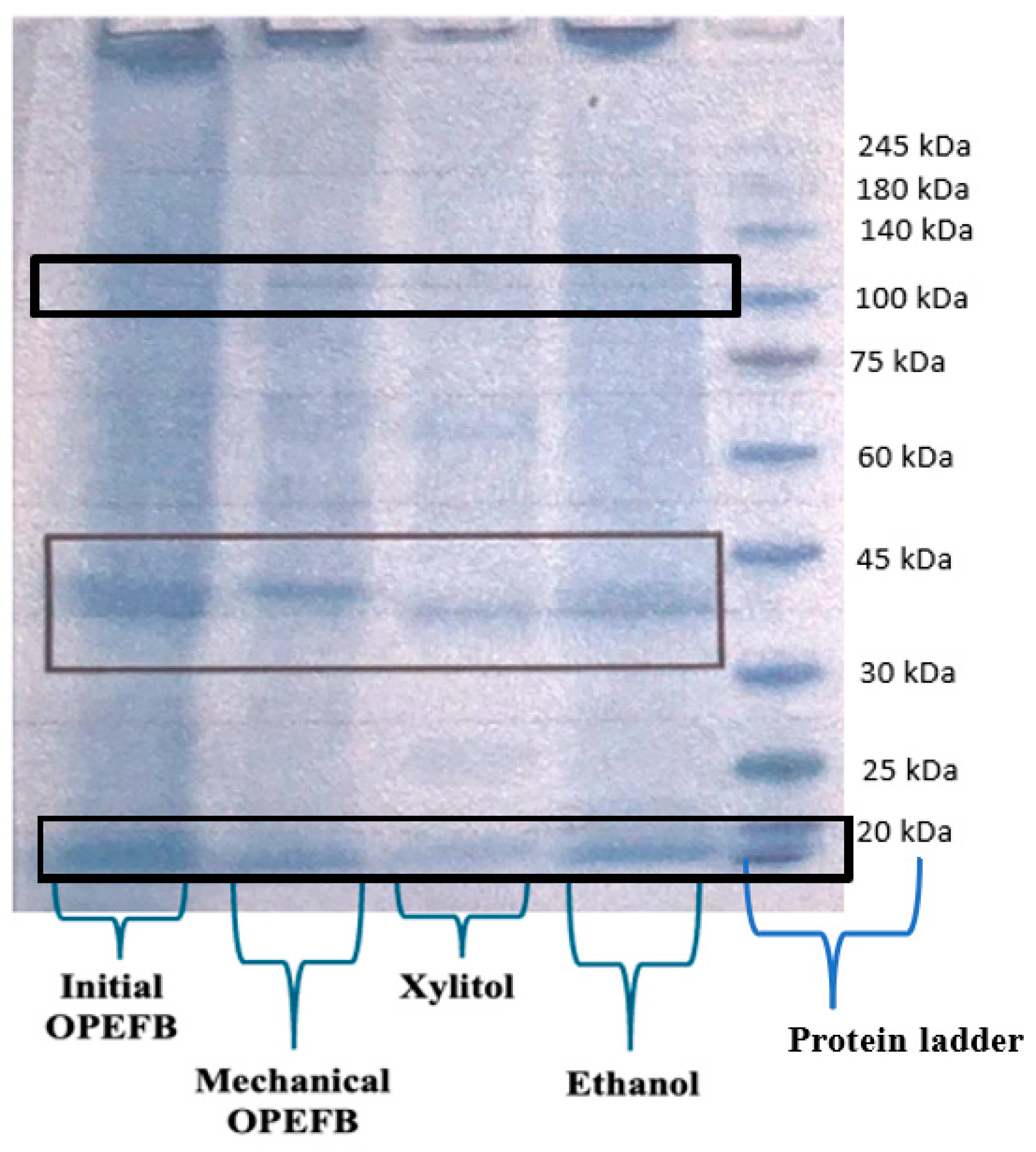
The study investigated variations of OPEFB substrates, including untreated and mechanically treated OPEFB, xylitol production residue, and ethanol fermentation residue. For enzyme production, 2.4 g of PDB and 100 mL of distilled water were homogenized using a magnetic stirrer. A concentration of 2.5 g of OPEFB biomass at a concentration of 2.5% was added and sterilized in an autoclave at 121 °C for 15 min. To enhance extracellular enzyme production, 1% Tween 80 was added to the fermentation flask. Fungal spore density was measured using a hemocytometer according to Avin’s procedure [
19]. Aseptically, a spore density of 6.8 × 10
7 cells/mL was inoculated into the fermentation flasks and incubated at 30 °C and 200 rpm for 9 days using an incubator shaker. Sampling was performed every three days, with the samples centrifuged at 6000 rpm for 20 min to separate the fungal biomass and extracellular enzymes in the supernatant. Subsequently, the supernatant was subjected to various tests to evaluate the enzymatic activity, protein concentration, specific activity, and molecular weight of xylanase, cellulase, and laccase enzymes.
2.7. Analysis Method
The lignocellulose composition was determined using the Van Soest method, as described by Hindrichsen et al. [
20]. This method enables the simultaneous quantification of cellulose, hemicellulose, and lignin concentrations in the biomass. For analyzing the hydrolysis and fermentation samples, high-performance liquid chromatography (HPLC) was conducted using a Waters 1515 pump, a 2707 autosampler, and UV and refractive index (RI) detectors with an Aminex HPX-87H column (Milford, MA, USA). The mobile phase used was 5 mM H
2SO
4 with a flow rate of 0.6 mL/min. The growth of
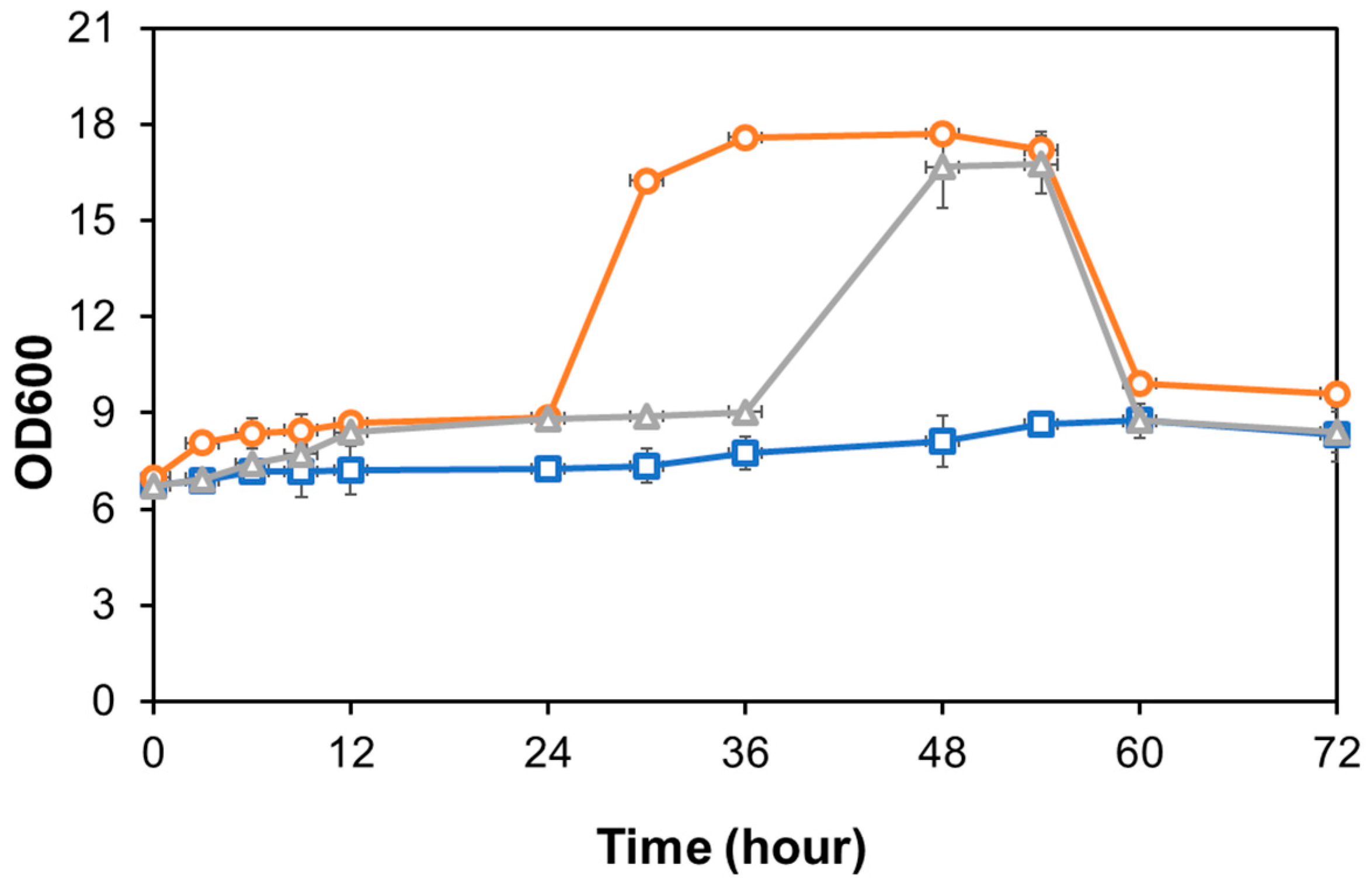
M. caribbica was monitored by measuring optical density at 600 nm and converted to dry cell weight (DCW) using a calibration curve. Meanwhile, to evaluate the growth of
Candida sp. Unpad CC Y26 cells, the total plate count (TPC) method was employed due to the precipitation of yeast cells during fermentation, and the calibration curve was also used to convert colony-forming units (CFUs) to dry cell weight (DCW). The growth of
Aspergillus niger cells can be measured based on their spore density using a hemocytometer. A scanning electron microscope (JEOL, JSM-6330F; Tokyo, Japan) was used to examine the surface of the OPEFB before and after various treatments.
2.8. Enzyme Activity Test
Enzyme activities of cellulase and xylanase were measured using the Jampala et al. [
21] method. Each test tube contained 0.5 mL of crude enzyme and 1 mL of 0.05 M citrate buffer pH 4.8. For cellulase and xylanase assays, 0.5 mL of 1% carboxymethyl cellulose (CMC) and 1% xylan were used as substrates, respectively. After 30 min incubation in a 50 °C water bath, 2 mL of DNS reagent was added, followed by heating in a 90 °C water bath for 10 min, then chilled. The color change was seen, and the absorbance was measured at 540 nm using a spectrophotometer. Enzyme activities were calculated based on standard glucose (
y = 92.347
x − 0.0633, R
2 = 0.9973) and xylose (
y = 40.118
x + 0.444, R
2 = 0.995) equations. The glucose and xylose standard curve (0–10 g/L) for cellulase activity from Adney and Baker [
22] was utilized.
The laccase activity was spectrophotometrically determined as per the method of Yuliana et al. [
23]. A mixture of 60 μL of 0.5 M acetate buffer at pH 5, 20 μL of 1 mM 2.2′-azino-bis(3-ethylbenzothiazoline-6-sulfonate) (ABTS), and 50 μL of the crude enzyme was incubated in a microplate at 37 °C for 30 min, with absorbance measured at 420 nm. Enzyme units were defined as the laccase activity that oxidized 1 μmol of ABTS per minute.
Specific activity refers to the number of enzyme activity units per milligram of protein under optimum conditions. This is commonly used to indicate the amount of enzyme involved in a specific enzymatic reaction. To determine the specific activity of xylanase, the enzyme activity units and xylanase protein concentration were measured [
24]. One unit of xylanase activity was defined as the amount of xylanase activity that produced 1 mg of xylose under optimum conditions. A higher specific activity value indicated a better ability of the enzyme to utilize the substrate.
2.9. Protein Content
The Bicinchoninic Acid (BCA) protein assay kit was used for protein quantification, following the Thermoscientific protocol [
25]. The protein testing was performed by adding 200 μL of BCA Working Reagent to each column of the microplate samples to be tested. The BCA Working Reagent was prepared by mixing BCA Reagent B with A in a ratio of 1:50. Subsequently, a total of 200 μL of BCA Working Reagent was added to each well containing 25 μL of the enzyme sample. After shaking for 30 s and incubating at 37 °C for 30 min, the absorbance was measured at 562 nm using a spectrophotometer. Protein concentration was derived from a BSA (0–200 g/L) standard curve (
y = 0.0008
x + 0.0064 (R
2 = 0.946).
2.10. Enzyme Molecular Weight
SDS-PAGE was performed using the Laemmli (1970) method with 15% separating and 4% stacking gels. After mixing 15 µL of the sample with 10 µL of loading dye, the mixture was heated at 95 °C for 10 min. Five microliters of samples were loaded into each well. Electrophoresis was carried out using Mini Protean Tetra Cell units (Bio-Rad Laboratories, Inc., Richmond, CA, USA) for approximately 60 min at 110 V. Proteins were stained with Simply Blue and destained with Aquades after separation. A broad-range molecular weight standard marker (10–245 kDa) was used to determine the molecular weight (MW) of the enzymes.
2.11. Statistical Analysis
Student t-test was used to determine the statistical significance of all measurements. The data were presented as mean ± standard deviation. A p-value of less than 0.05 was considered statistically significant.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}