
Effect of Alkaline and Mechanical Pretreatment of Wheat Straw on Enrichment Cultures from Pachnoda marginata Larva Gut
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Medium, Wheat Straw Pretreatment, and Experiment Setup
2.2. Analytical Methods
2.3. Microbial Community Analysis
2.4. Scanning Electron Microscopy (SEM)
2.5. Statistical Analysis
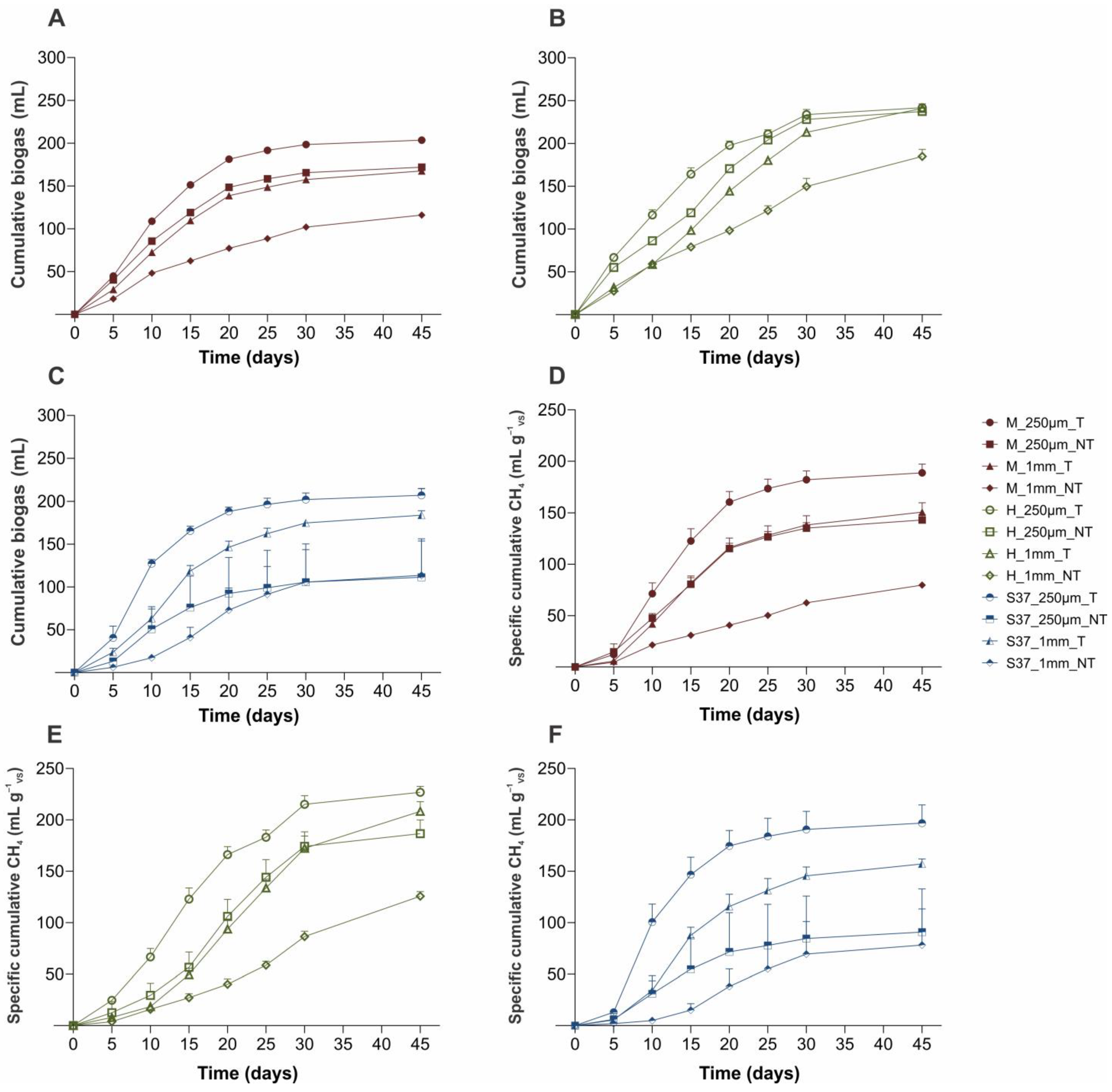
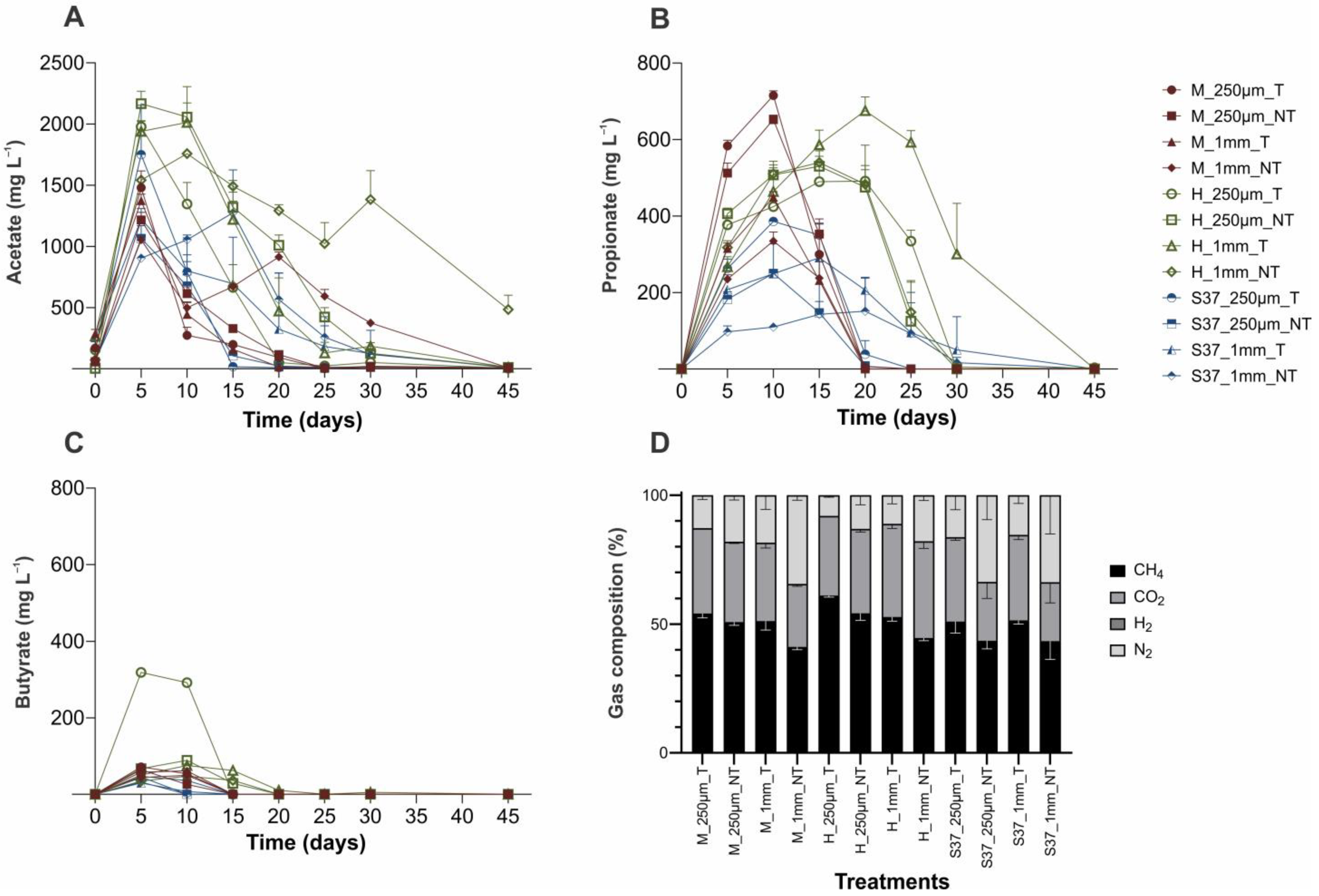
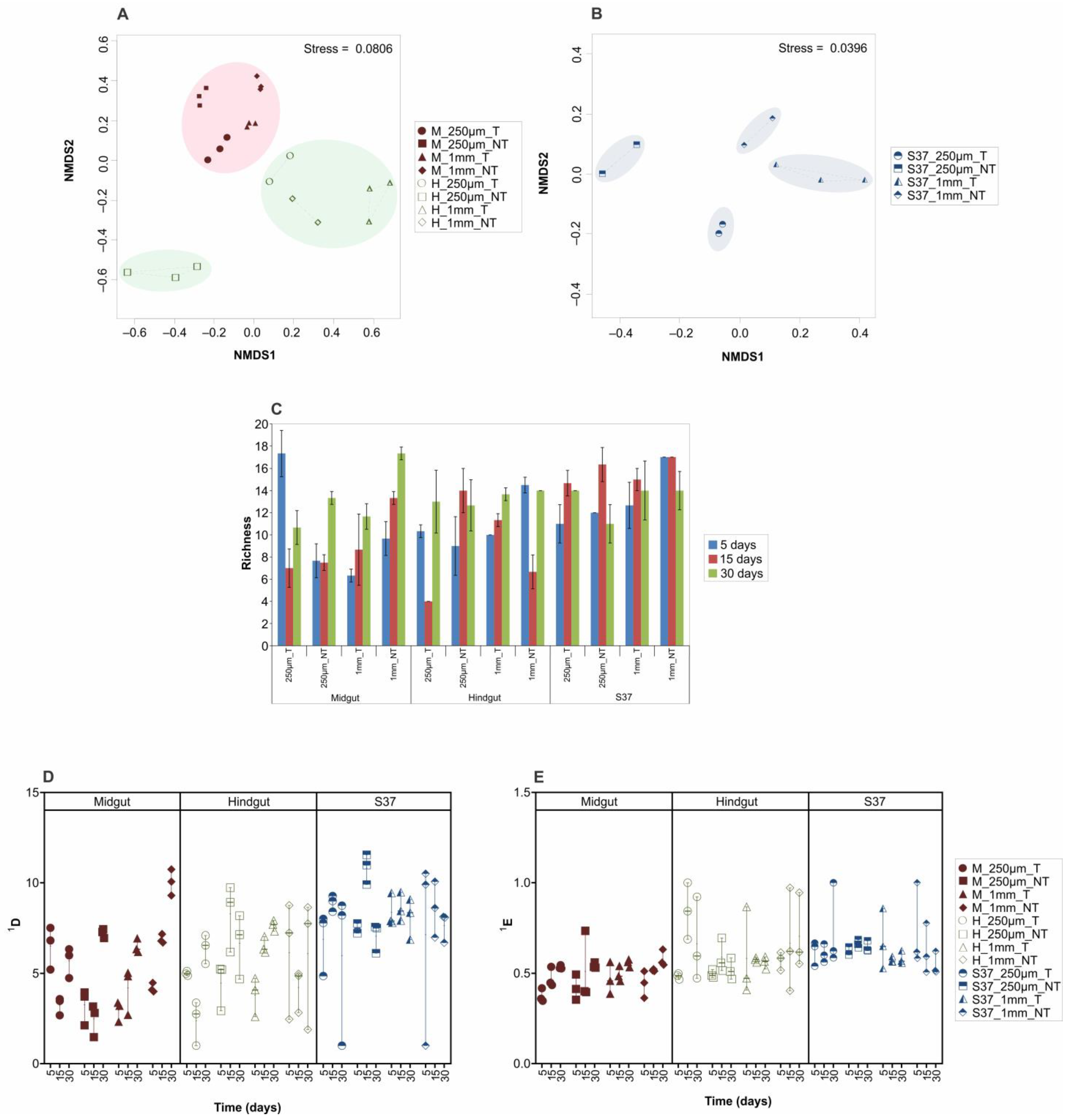
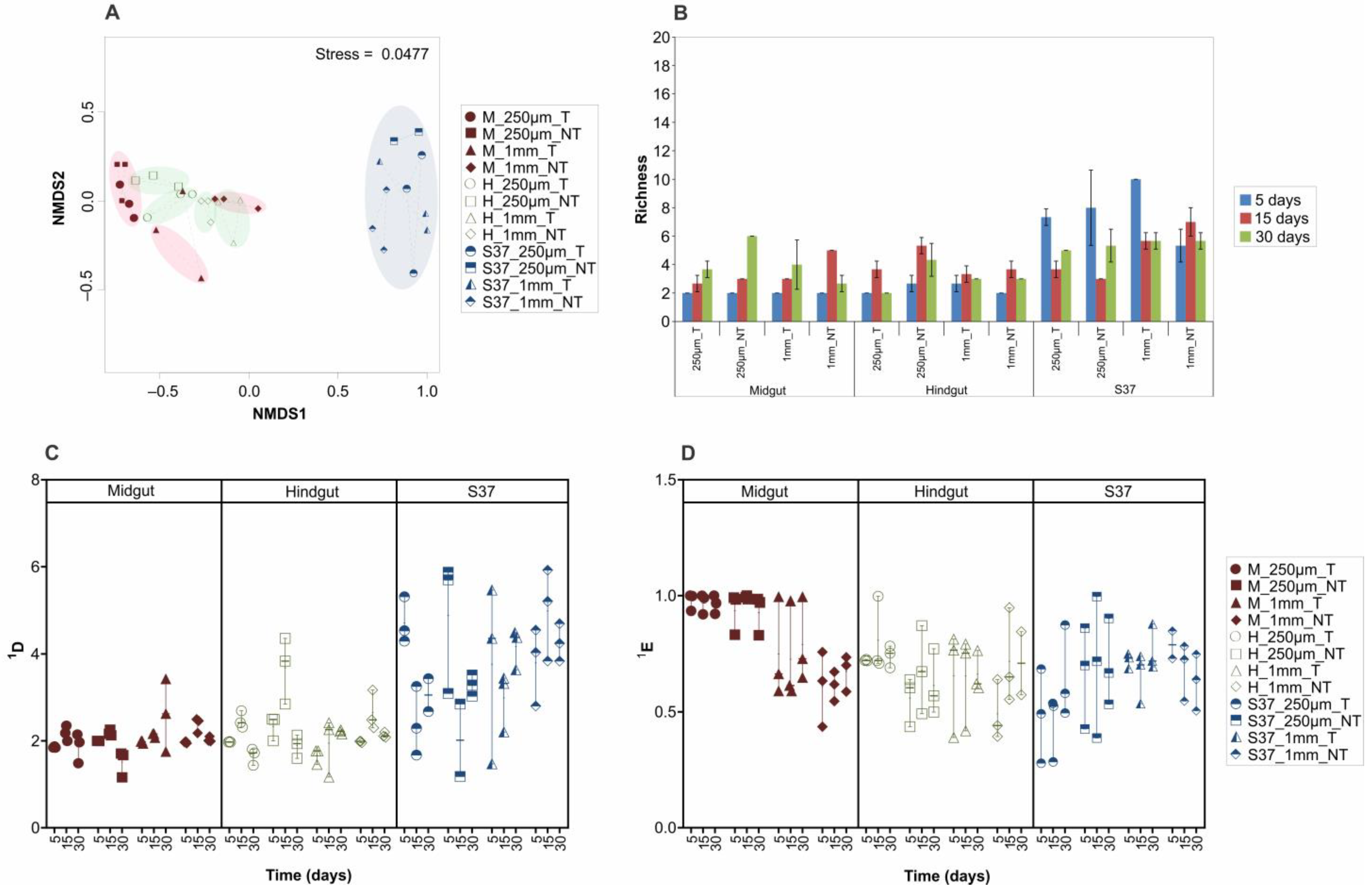
3. Results and Discussion
3.1. Physical-Chemical Parameters
3.2. Scanning Electron Microscopy
3.3. Microbial Community
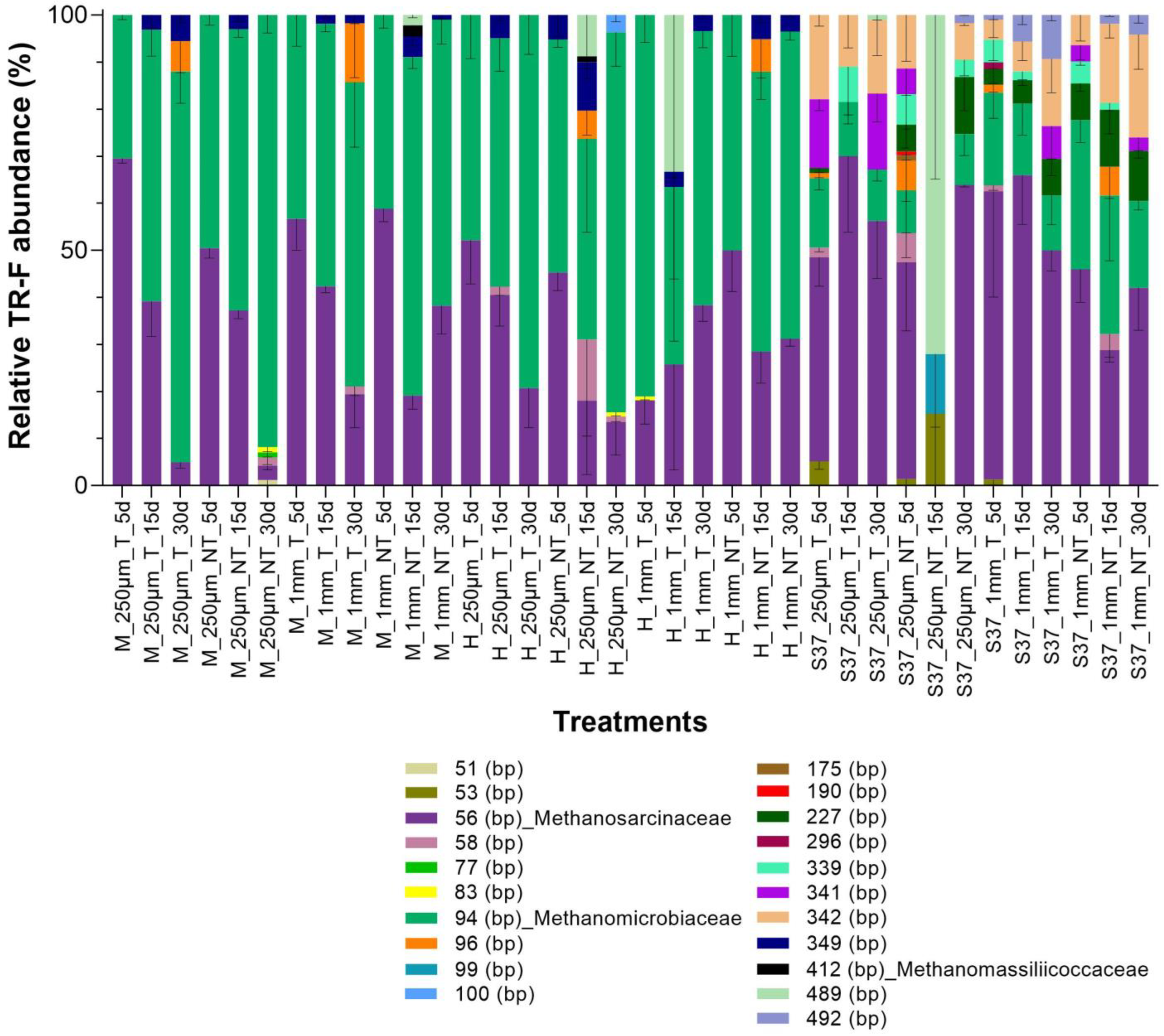
3.4. Methanogenic Community Structure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dumond, L.; Lam, P.Y.; Van Erven, G.; Kabel, M.; Mounet, F.; Grima-Pettenati, J.; Tobimatsu, Y.; Hernandez-Raquet, G. Termite Gut Microbiota Contribution to Wheat Straw Delignification in Anaerobic Bioreactors. ACS Sustain. Chem. Eng. 2021, 9, 2191–2202. [Google Scholar] [CrossRef]
- Saini, J.K.; Saini, R.; Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. 3 Biotech 2015, 5, 337–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noike, T.; Endo, G.; Chang, J.; Yaguchi, J.; Matsumoto, J. Characteristics of Carbohydrate Degradation and the Rate-limiting Step in Anaerobic Digestion. Biotechnol. Bioeng. 1985, 27, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Vavilin, V.A. A description of hydrolysis kinetics in anaerobic degradation of particulate organic matter. Bioresour. Technol. J. 1996, 56, 229–237. [Google Scholar] [CrossRef]
- Ahorsu, R.; Medina, F.; Constantí, M. Significance and Challenges of Biomass as a Suitable Feedstock for Bioenergy and Biochemical Production: A Review. Energies 2018, 11, 3366. [Google Scholar] [CrossRef] [Green Version]
- Mosier, N.; Wyman, C.; Dale, B.; Elander, R.; Lee, Y.Y.; Holtzapple, M.; Ladisch, M. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 2005, 96, 673–686. [Google Scholar] [CrossRef]
- Awogbemi, O.; Kallon, D.V. Von Case Studies in Chemical and Environmental Engineering Pretreatment techniques for agricultural waste. Case Stud. Chem. Environ. Eng. 2022, 6, 100229. [Google Scholar] [CrossRef]
- Khan, M.U.; Usman, M.; Ashraf, M.A.; Dutta, N.; Luo, G.; Zhang, S. A review of recent advancements in pretreatment techniques of lignocellulosic materials for biogas production: Opportunities and Limitations. Chem. Eng. J. Adv. 2022, 10, 100263. [Google Scholar] [CrossRef]
- Mohammad Rahmani, A.; Gahlot, P.; Moustakas, K.; Kazmi, A.A.; Shekhar Prasad Ojha, C.; Tyagi, V.K. Pretreatment methods to enhance solubilization and anaerobic biodegradability of lignocellulosic biomass (wheat straw): Progress and challenges. Fuel 2022, 319, 123726. [Google Scholar] [CrossRef]
- Kumar, R.; Kim, T.H.; Basak, B.; Patil, S.M.; Kim, H.H.; Ahn, Y.; Yadav, K.K.; Cabral-Pinto, M.M.S.; Jeon, B.H. Emerging approaches in lignocellulosic biomass pretreatment and anaerobic bioprocesses for sustainable biofuels production. J. Clean. Prod. 2022, 333, 130180. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, Z.F.; Zhang, C.C.; Nan, J.; Ren, N.Q.; Lee, D.J.; Chen, C. Advances in pretreatment of lignocellulosic biomass for bioenergy production: Challenges and perspectives. Bioresour. Technol. 2022, 343, 126123. [Google Scholar] [CrossRef]
- Show, B.K.; Banerjee, S.; Banerjee, A.; GhoshThakur, R.; Hazra, A.K.; Mandal, N.C.; Ross, A.B.; Balachandran, S.; Chaudhury, S. Insect gut bacteria: A promising tool for enhanced biogas production. Rev. Environ. Sci. Biotechnol. 2022, 21, 1–25. [Google Scholar] [CrossRef]
- Chen, H. “Biotechnology of Lignocellulose.” Theory and Practice; Springer: Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; New York, NY, USA; London, UK, 2014. [Google Scholar] [CrossRef]
- Monlau, F.; Barakat, A.; Trably, E.; Dumas, C.; Steyer, J.P.; Carrère, H. Lignocellulosic materials into biohydrogen and biomethane: Impact of structural features and pretreatment. Crit. Rev. Environ. Sci. Technol. 2013, 43, 260–322. [Google Scholar] [CrossRef]
- Kumar, S.; Gandhi, P.; Yadav, M.; Paritosh, K.; Pareek, N.; Vivekanand, V. Weak alkaline treatment of wheat and pearl millet straw for enhanced biogas production and its economic analysis. Renew. Energy 2019, 139, 753–764. [Google Scholar] [CrossRef]
- Durot, N.; Gaudard, F.; Kurek, B. The unmasking of lignin structures in wheat straw by alkali. Phytochemistry 2003, 63, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Bali, G.; Meng, X.; Deneff, J.I.; Sun, Q.; Ragauskas, A.J. The Effect of Alkaline Pretreatment Methods on Cellulose Structure and Accessibility. ChemSusChem 2015, 8, 275–279. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Xu, F.; Li, Y. Pretreatment of lignocellulosic biomass for enhanced biogas production. Prog. Energy Combust. Sci. 2014, 42, 35–53. [Google Scholar] [CrossRef]
- Moset, V.; de Almeida Neves Xavier, C.; Feng, L.; Wahid, R.; Møller, H.B. Combined low thermal alkali addition and mechanical pre-treatment to improve biogas yield from wheat straw. J. Clean. Prod. 2018, 172, 1391–1398. [Google Scholar] [CrossRef]
- Rani, P.; Bansal, M.; Pathak, V.V. Experimental and kinetic studies for improvement of biogas production from KOH pretreated wheat straw. Curr. Res. Green Sustain. Chem. 2022, 5, 100283. [Google Scholar] [CrossRef]
- Liu, X.; Zicari, S.M.; Liu, G.; Li, Y.; Zhang, R. Pretreatment of wheat straw with potassium hydroxide for increasing enzymatic and microbial degradability. Bioresour. Technol. 2015, 185, 150–157. [Google Scholar] [CrossRef]
- Jaffar, M.; Pang, Y.; Yuan, H.; Zou, D.; Liu, Y.; Zhu, B.; Korai, R.M.; Li, X. Wheat straw pretreatment with KOH for enhancing biomethane production and fertilizer value in anaerobic digestion. Chin. J. Chem. Eng. 2016, 24, 404–409. [Google Scholar] [CrossRef]
- Cern, W.; Rabaey, K.; Vervaeren, H. Low temperature calcium hydroxide treatment enhances anaerobic methane production from (extruded) biomass. Bioresour. Technol. 2015, 176, 181–188. [Google Scholar] [CrossRef]
- Fernandes, T.V.; Bos, G.J.K.; Zeeman, G.; Sanders, J.P.M.; Lier, J.B. Van Effects of thermo-chemical pre-treatment on anaerobic biodegradability and hydrolysis of lignocellulosic biomass. Bioresour. Technol. 2009, 100, 2575–2579. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Holtzapple, M.T. Lime pretreatment and enzymatic hydrolysis of corn stover. Bioresour. Technol. 2005, 96, 1994–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for Pretreatment of Lignocellulosic Biomass for Efficient Hydrolysis and Biofuel Production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Gu, W.; Bousfield, D.W.; Tripp, C.P. Formation of calcium carbonate particles by direct contact of Ca(OH)2 powders with supercritical CO2. J. Mater. Chem. 2006, 16, 3312–3317. [Google Scholar] [CrossRef]
- Ukrainczyk, M.; Kontrec, J.; Babić-Ivančić, V.; Brečević, L.; Kralj, D. Experimental design approach to calcium carbonate precipitation in a semicontinuous process. Powder Technol. 2007, 171, 192–199. [Google Scholar] [CrossRef]
- Li, X.; Peng, Y.; Ren, N.; Li, B.; Chai, T. Effect of temperature on short chain fatty acids (SCFAs) accumulation and microbiological transformation in sludge alkaline fermentation with Ca(OH)2 adjustment. Water Res. 2014, 61, 34–45. [Google Scholar] [CrossRef]
- Xu, J.; Du, Y.; Jiang, Z.; She, A. Effects of calcium source on biochemical properties of microbial CaCo3 precipitation. Front. Microbiol. 2015, 6, 1366. [Google Scholar] [CrossRef]
- Dieudonné, A.; Battimelli, A.; Philippe, J.; Hélène, S.; Renaud, C.; Godon, J.J. Mechanical Pretreatment of Various Types of Biomass from Animals: What Potential Applications to Anaerobic Digestion? BioEnergy Res. 2022. [Google Scholar] [CrossRef]
- Andersen, L.F.; Parsin, S.; Lüdtke, O.; Kaltschmitt, M. Biogas production from straw—The challenge feedstock pretreatment. Biomass Convers. Biorefin. 2022, 12, 379–402. [Google Scholar] [CrossRef]
- Dumas, C.; Silva Ghizzi Damasceno, G.; Abdellatif, B.; Carrère, H.; Steyer, J.P.; Rouau, X. Effects of grinding processes on anaerobic digestion of wheat straw. Ind. Crops Prod. 2015, 74, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Ozbayram, E.G.; Kleinsteuber, S.; Nikolausz, M. Biotechnological utilization of animal gut microbiota for valorization of lignocellulosic biomass. Appl. Microbiol. Biotechnol. 2019, 104, 489–508. [Google Scholar] [CrossRef]
- Lazuka, A.; Auer, L.; Donohue, M.O.; Raquet, G.H. Anaerobic lignocellulolytic microbial consortium derived from termite gut: Enrichment, lignocellulose degradation and community dynamics. Biotechnol. Biofuels 2018, 11, 284. [Google Scholar] [CrossRef]
- Lemke, T.; Stingl, U.; Egert, M.; Friedrich, M.W.; Brune, A. Physicochemical Conditions and Microbial Activities in the Highly Alkaline Gut of the Humus-Feeding Larva of Pachnoda ephippiata (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 2003, 69, 6650–6658. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, B.G.; Logroño, W.; Nunes da Rocha, U.; Harms, H.; Nikolausz, M. Enrichment of Anaerobic Microbial Communities from Midgut and Hindgut of Sun Beetle Larvae (Pachnoda marginata) on Wheat Straw: Effect of Inoculum Preparation. Microorganisms 2022, 10, 761. [Google Scholar] [CrossRef]
- Cazemier, A.E.; Verdoes, J.C.; Reubsaet, F.A.G.; Hackstein, J.H.P.; van der Drift, C.; Op den Camp, H.J.M. Promicromonospora pachnodae sp. nov., a member of the (hemi)cellulolytic hindgut flora of larvae of the scarab beetle Pachnoda marginata. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2003, 83, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Golwala, S.; Kothari, D.K. Maximizing Biogas Yield Through Inoculum and Optimizing Various Operating Parameters: A Review. SSRN Electron. J. 2021, 1–8. [Google Scholar] [CrossRef]
- Tsapekos, P.; Kougias, P.G.; Vasileiou, S.A.; Lyberatos, G.; Angelidaki, I. Effect of micro-aeration and inoculum type on the biodegradation of lignocellulosic substrate. Bioresour. Technol. 2017, 225, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.A.; Sheikh, Z. Effect of inoculum type and organic loading on biogas production of sunflower meal and wheat straw. Sustain. Environ. Res. 2019, 29, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Yun, J.; Zhang, H.; Yu, Z. Full-scale anaerobic reactor samples would be more suitable than lab-scale anaerobic reactor and natural samples to inoculate the wheat straw batch anaerobic digesters. Bioresour. Technol. 2019, 293, 122040. [Google Scholar] [CrossRef] [PubMed]
- Andrew, G. Hashimoto Effect of Inoculum/Substrate Ratio on Methane Yield and Production Rate from Straw. Biol. Wastes 1989, 28, 247–255. [Google Scholar]
- Lawal, A.A.; Dzivama, A.U.; Wasinda, M.K. Effect of inoculum to substrate ratio on biogas production of sheep paunch manure. Res. Agric. Eng. 2016, 62, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, M.; Andres, Y.; Blel, W.; Hassan, R.; Ahmed, A. Effect of inoculum VS, organic loads and I/S on the biochemical methane potential of sludge, buckwheat husk and straw. Desalin. Water Treat. 2019, 157, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Chen, X.; Liu, Z.; Zhou, X.; Zhang, Y. Effect of inoculum sources on the anaerobic digestion of rice straw. Bioresour. Technol. 2014, 158, 149–155. [Google Scholar] [CrossRef]
- Jensen, M.B.; De Jonge, N.; Dolriis, M.D.; Kragelund, C.; Møller, H.B.; Ditlev, L.; Ottosen, M.; Nielsen, J.L. Cellulolytic and Xylanolytic Microbial Communities Associated With Lignocellulose-Rich Wheat Straw. Front. Microbiol. 2021, 12, 645174. [Google Scholar] [CrossRef]
- Veluchamy, C.; Kalamdhad, A.S. Enhanced methane production and its kinetics model of thermally pretreated lignocellulose waste material. Bioresour. Technol. 2017, 241, 1–9. [Google Scholar] [CrossRef]
- Kang, Y.R.; Su, Y.; Wang, J.; Chu, Y.X.; Tian, G.; He, R. Effects of different pretreatment methods on biogas production and microbial community in anaerobic digestion of wheat straw. Environ. Sci. Pollut. Res. 2021, 28, 51772–51785. [Google Scholar] [CrossRef]
- Ozbayram, E.G.; Kleinsteuber, S.; Nikolausz, M.; Ince, B.; Ince, O. Enrichment of lignocellulose-degrading microbial communities from natural and engineered methanogenic environments. Appl. Microbiol. Biotechnol. 2018, 102, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Rajeswari, G.; Jacob, S.; Chandel, A.K.; Kumar, V. Unlocking the potential of insect and ruminant host symbionts for recycling of lignocellulosic carbon with a biorefinery approach: A review. Microb. Cell Fact. 2021, 20, 107. [Google Scholar] [CrossRef] [PubMed]
- Gales, A.; Chatellard, L.; Abadie, M.; Bonnafous, A.; Auer, L. Screening of Phytophagous and Xylophagous Insects Guts Microbiota Abilities to Degrade Lignocellulose in Bioreactor. Front. Microbiol. 2018, 9, 2222. [Google Scholar] [CrossRef]
- Mohammed, W.S.; Ziganshina, E.E.; Shagimardanova, E.I.; Gogoleva, N.E.; Ziganshin, A.M. Comparison of intestinal bacterial and fungal communities across various xylophagous beetle larvae (Coleoptera: Cerambycidae). Nat. Sci. Rep. 2018, 8, 10073. [Google Scholar] [CrossRef] [Green Version]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Auer, L.; Lazuka, A.; Sillam-dussès, D.; Miambi, E.; Donohue, M.O.; Hernandez-raquet, G. Uncovering the Potential of Termite Gut Microbiome for Lignocellulose Bioconversion in Anaerobic Batch Bioreactors. Front. Microbiol. 2017, 8, 2623. [Google Scholar] [CrossRef] [Green Version]
- Porsch, K.; Wirth, B.; Tóth, E.M.; Schattenberg, F.; Nikolausz, M. Characterization of wheat straw-degrading anaerobic alkali-tolerant mixed cultures from soda lake sediments by molecular and cultivation techniques. Microb. Biotechnol. 2015, 8, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Grieve, A.D.; Gurd, G.W.; Maass, O. The solubility of lime in water and the specific conductivities of its saturated solutions. Can. J. Res. 1933, 8, 577–582. [Google Scholar] [CrossRef]
- Logroño, W.; Popp, D.; Kleinsteuber, S.; Sträuber, H.; Harms, H. Microbial Resource Management for Ex Situ Biomethanation of Hydrogen at Alkaline pH. Microorganisms 2020, 8, 614. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Regan, J.M. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 2008, 74, 6663–6671. [Google Scholar] [CrossRef]
- Bühligen, F.; Lucas, R.; Nikolausz, M.; Kleinsteuber, S. A T-RFLP database for the rapid profiling of methanogenic communities in anaerobic digesters. Anaerobe 2016, 39, 114–116. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.; Harms, H.; Johst, K.; Frank, K.; Kleinsteuber, S. A critical evaluation of ecological indices for the comparative analysis of microbial communities based on molecular datasets. FEMS Microbiol. Ecol. 2017, 93, fiw209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memon, M.J.; Memon, A.R. Wheat straw optimization via its efficient pretreatment for improved biogas production. Civ. Eng. J. 2020, 6, 1056–1063. [Google Scholar] [CrossRef]
- Cazemier, A.E.; Den Camp, H.J.M.O.; Hackstein, J.H.P.; Vogels, G.D. Fibre digestion in arthropods. Comp. Biochem. Physiol. A Physiol. 1997, 118, 101–109. [Google Scholar] [CrossRef]
- Momoh, O.L.Y.; Ouki, S. Development of a novel fractal-like kinetic model for elucidating the effect of particle size on the mechanism of hydrolysis and biogas yield from ligno-cellulosic biomass. Renew. Energy 2018, 118, 71–83. [Google Scholar] [CrossRef]
- Sträuber, H.; Bühligen, F.; Kleinsteuber, S.; Nikolausz, M.; Porsch, K. Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms. Bioengineering 2015, 2, 66–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, R.; Takeuchi, H.; Hasegawa, T.; Vijay, V.K. Experimental evaluation of substrate’s particle size of wheat and rice straw biomass on methane production yield. Agric. Eng. Int. CIGR J. 2015, 17, 93–104. [Google Scholar]
- Vasmara, C.; Cianchetta, S.; Marchetti, R.; Galletti, S. Biogas production from wheat straw pre-treated with hydrolytic enzymes or sodium hydroxide. Environ. Eng. Manag. J. 2017, 16, 1827–1835. [Google Scholar] [CrossRef]
- Vasmara, C.; Cianchetta, S.; Marchetti, R.; Galletti, S. Biogas production from wheat straw pre-treated with ligninolytic fungi and co-digestion with pig slurry. Environ. Eng. Manag. J. 2015, 14, 1751–1760. [Google Scholar] [CrossRef]
- Sambusiti, C.; Monlau, F.; Ficara, E.; Carrère, H.; Malpei, F. A comparison of different pre-treatments to increase methane production from two agricultural substrates. Appl. Energy 2013, 104, 62–70. [Google Scholar] [CrossRef]
- Zerback, T.; Schumacher, B.; Weinrich, S.; Hülsemann, B.; Nelles, M. Hydrothermal Pretreatment of Wheat Straw—Evaluating the Effect of Substrate Disintegration on the Digestibility in Anaerobic Digestion. Processes 2022, 10, 1048. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, C. Anaerobic codigestion of pretreated wheat straw with cattle manure and analysis of the microbial community. Bioresour. Technol. 2015, 186, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Pavlostathis, S.G.; Gossett, J.M. Alkaline Treatment of Wheat Straw for Increasing Anaerobic Biodegradability. Biotechnol. Bioeng. 1985, 27, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Zhang, J.; Zhang, J. Roles of calcium-containing alkali materials on dark fermentation and anaerobic digestion: A systematic review. Int. J. Hydrog. Energy 2021, 46, 38645–38662. [Google Scholar] [CrossRef]
- Reilly, M.; Dinsdale, R.; Guwy, A. Enhanced biomethane potential from wheat straw by low temperature alkaline calcium hydroxide pre-treatment. Bioresour. Technol. 2015, 189, 258–265. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Li, H.; Radwan, A.A.; Sheng, K.; Chen, X. Effect of hydrothermal and Ca(OH)2 pretreatments on anaerobic digestion of sugarcane bagasse for biogas production. Bioresour. Technol. 2018, 259, 54–60. [Google Scholar] [CrossRef]
- Guan, R.; Li, X.; Chufo, A.; Yuan, H.; Liu, Y.; Zou, D.; Zuo, X.; Gu, J. Enhancing anaerobic digestion performance and degradation of lignocellulosic components of rice straw by combined biological and chemical pretreatment. Sci. Total Environ. 2018, 637–638, 9–17. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, Y.; Zhou, X. Effect of Ca(OH)2 pretreatment on extruded rice straw anaerobic digestion. Bioresour. Technol. 2015, 196, 116–122. [Google Scholar] [CrossRef]
- Taherdanak, M.; Zilouei, H. Improving biogas production from wheat plant using alkaline pretreatment. Fuel 2014, 115, 714–719. [Google Scholar] [CrossRef]
- Chandra, R.; Takeuchi, H.; Hasegawa, T.; Kumar, R. Improving biodegradability and biogas production of wheat straw substrates using sodium hydroxide and hydrothermal pretreatments. Energy 2012, 43, 273–282. [Google Scholar] [CrossRef]
- Janke, L.; Weinrich, S.; Leite, A.F.; Sträuber, H. Pre-treatment of filter cake for anaerobic digestion in sugarcane biorefineries: Assessment of batch versus semi-continuous experiments. Renew. Energy 2019, 143, 1416–1426. [Google Scholar] [CrossRef]
- Ghosh, P.; Shah, G.; Sahota, S.; Singh, L.; Vijay, V.K. Biogas Production from Waste: Technical Overview, Progress, and Challenges. In Bioreactors; Elsevier: Amsterdam, The Netherlands, 2020; pp. 89–104. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, R.; Liu, G.; Chen, C.; He, Y.; Liu, X. Comparison of methane production potential, biodegradability, and kinetics of different organic substrates. Bioresour. Technol. 2013, 149, 565–569. [Google Scholar] [CrossRef]
- Gao, J.; Li, J.; Wachemo, A.C.; Yuan, H.; Zuo, X.; Li, X. Mass conversion pathway during anaerobic digestion of wheat straw. RSC Adv. 2020, 10, 27720–27727. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Z.; Zou, H.; Yang, H.; Sun, T.; Li, M. Digestive performance of sludge with different crop straws in mesophilic anaerobic digestion. Bioresour. Technol. 2019, 289, 121595. [Google Scholar] [CrossRef] [PubMed]
- De la Rubia, M.A.; Fernández-Cegrí, V.; Raposo, F.; Borja, R. Influence of particle size and chemical composition on the performance and kinetics of anaerobic digestion process of sunflower oil cake in batch mode. Biochem. Eng. J. 2011, 58–59, 162–167. [Google Scholar] [CrossRef]
- Asghar, U.; Irfan, M.; Iram, M.; Huma, Z.; Nelofer, R.; Nadeem, M.; Syed, Q. Effect of alkaline pretreatment on delignification of wheat straw. Nat. Prod. Res. 2014, 29, 125–131. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Flemming, H. The perfect slime. Colloids Surf. B Biointerfaces 2011, 86, 251–259. [Google Scholar] [CrossRef]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Maddela, N.R.; Zhou, Z.; Yu, Z.; Zhao, S.; Menga, F. Functional Determinants of Extracellular Polymeric Substances in Membrane Biofouling: Experimental Evidence from Pure-Cultured Sludge Bacteria. Appl. Environ. Microbiol. 2018, 84, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, M.; Kim, K.S. Studies on the interactions of Ca2+ and Mg2+ with EPS and their role in determining the physicochemical characteristics of granular sludges in SBR system. Process Biochem. 2015, 50, 966–972. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, L.; Li, M.; Xu, Z.; Zhu, W. Effects of Ca and Mg levels on colony formation and EPS content of cultured M. aeruginosa. Procedia Environ. Sci. 2011, 10, 1452–1458. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Qian, D.; Geng, Z.; Dai, K.; Zhang, W.; Van Loosdrecht, M.C.M.; Zeng, R.J. Enhanced Methane Recovery from Waste-Activated Sludge by Alginate-Degrading Consortia: The Overlooked Role of Alginate in Extracellular Polymeric Substances. Environ. Sci. Technol. Lett. 2020, 8, 86–91. [Google Scholar] [CrossRef]
- Boone, D.R.; Whitman, W.B.; Koga, Y. Methanosarcinaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; ISBN 9781118960608. [Google Scholar]
- Dridi, B.; Fardeau, M.; Ollivier, B.; Raoult, D.; Drancourt, M. Methanomassiliicoccus luminyensis gen. nov., sp. nov., a methanogenic archaeon isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2012, 62, 1902–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles, G.; Nair, R.B.; Kleinsteuber, S.; Nikolausz, M.; Horváth, I.S. Biogas Production: Microbiological Aspects. In Biogas; Springer: Cham, Switzerland, 2018; pp. 163–198. [Google Scholar] [CrossRef]
- Ma, S.; Jiang, F.; Huang, Y.; Zhang, Y.; Wang, S.; Fan, H.; Liu, B.; Li, Q.; Yin, L.; Wang, H.; et al. A microbial gene catalog of anaerobic digestion from full-scale biogas plants. Gigascience 2021, 10, giaa164. [Google Scholar] [CrossRef]
- Paul, S.S.; Deb, S.M.; Dey, A.; Somvanshi, S.P.S.; Singh, D.; Rathore, R.; Stiverson, J. 16S rDNA analysis of archaea indicates dominance of Methanobacterium and high abundance of Methanomassiliicoccaceae in rumen of Nili-Ravi buffalo. Anaerobe 2015, 35, 3–10. [Google Scholar] [CrossRef]
- Ince, O.; Akyol, Ç.; Ozbayram, E.G.; Tutal, B.; Ince, B. Enhancing Methane Production from Anaerobic Co-Digestion of Cow Manure and Barley: Link between Process Parameters and Microbial Community Dynamics. Environ. Prog. Sustain. Energy 2020, 39, 13292. [Google Scholar] [CrossRef]
- Duan, N.; Kougias, P.G.; Campanaro, S.; Treu, L.; Angelidaki, I. Evolution of the microbial community structure in biogas reactors inoculated with seeds from different origin. Sci. Total Environ. 2021, 773, 144981. [Google Scholar] [CrossRef]
- Hori, T.; Haruta, S.; Sasaki, D.; Hanajima, D.; Ueno, Y.; Ogata, A.; Ishii, M.; Igarashi, Y. Reorganization of the bacterial and archaeal populations associated with organic loading conditions in a thermophilic anaerobic digester. J. Biosci. Bioeng. 2015, 119, 337–344. [Google Scholar] [CrossRef]
- Moestedt, J.; Müller, B.; Westerholm, M.; Schnürer, A. Ammonia threshold for inhibition of anaerobic digestion of thin stillage and the importance of organic loading rate. Microb. Biotechnol. 2016, 9, 180–194. [Google Scholar] [CrossRef]
- Campanaro, S.; Treu, L.; Kougias, P.G.; De Francisci, D.; Valle, G. Metagenomic analysis and functional characterization of the biogas microbiome using high throughput shotgun sequencing and a novel binning strategy. Biotechnol. Biofuels 2016, 9, 26. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeder, B.G.; İstanbullu, H.B.; Schmidt, M.; Logroño, W.; Harms, H.; Nikolausz, M. Effect of Alkaline and Mechanical Pretreatment of Wheat Straw on Enrichment Cultures from Pachnoda marginata Larva Gut. Fermentation 2023, 9, 60. https://doi.org/10.3390/fermentation9010060
Schroeder BG, İstanbullu HB, Schmidt M, Logroño W, Harms H, Nikolausz M. Effect of Alkaline and Mechanical Pretreatment of Wheat Straw on Enrichment Cultures from Pachnoda marginata Larva Gut. Fermentation. 2023; 9(1):60. https://doi.org/10.3390/fermentation9010060
Chicago/Turabian StyleSchroeder, Bruna Grosch, Havva Betül İstanbullu, Matthias Schmidt, Washington Logroño, Hauke Harms, and Marcell Nikolausz. 2023. "Effect of Alkaline and Mechanical Pretreatment of Wheat Straw on Enrichment Cultures from Pachnoda marginata Larva Gut" Fermentation 9, no. 1: 60. https://doi.org/10.3390/fermentation9010060