

Optimization of Solid-Phase Lactobacillus Fermentation Conditions to Increase γ-Aminobutyric Acid (GABA) Content in Selected Substrates
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Lactic Acid Bacteria Fermentation Conditions
2.3. Analysis of Glutamic Acid and GABA Content
2.4. DPPH Readical Scavenging Acitivity
2.5. ABTS Readical Scavenging Acitivity
2.6. Statistical Analysis
3. Results and Discussions
3.1. Identification and Qualification of Glutamic Acid and GABA
3.2. Changes in GABA Content According to Fermentation Conditions
3.2.1. Effect of Fermentation Strains on GABA Content
3.2.2. Effect of Fermentation Temperature on GABA Content
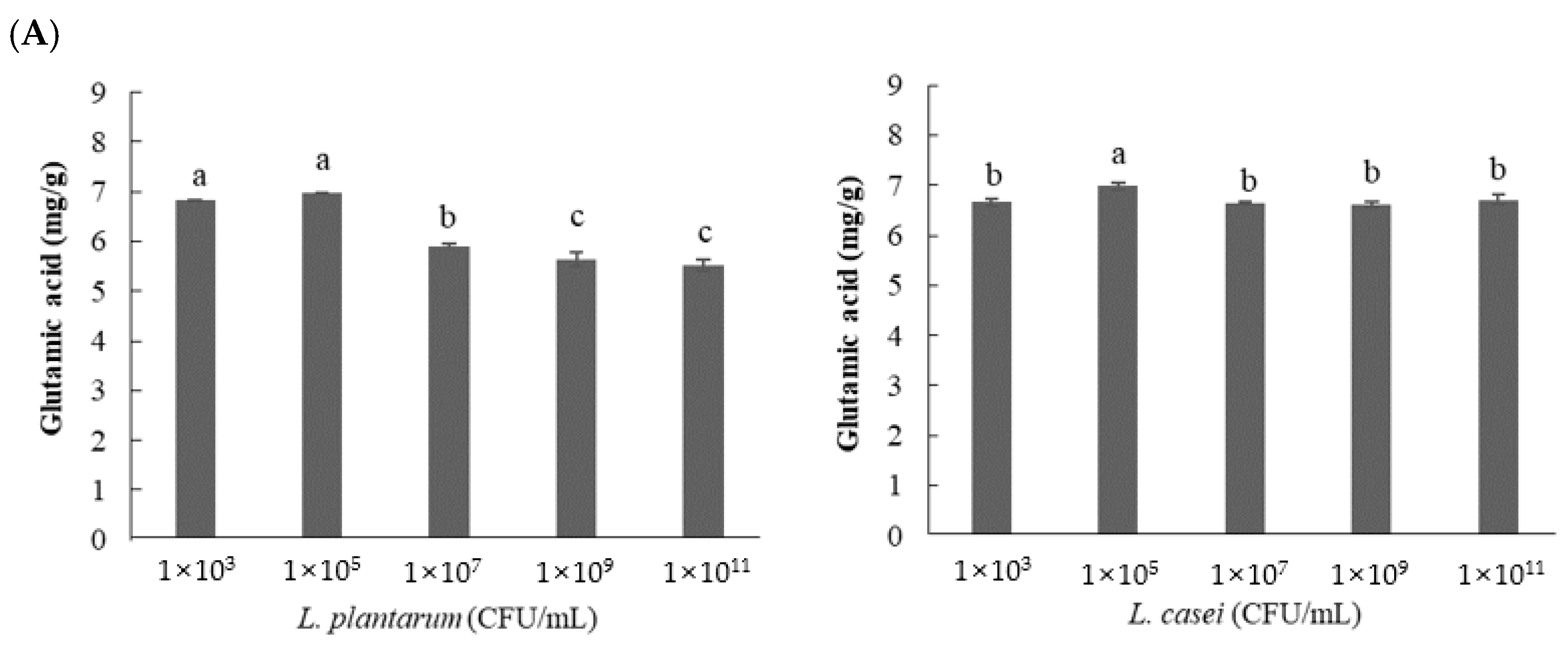
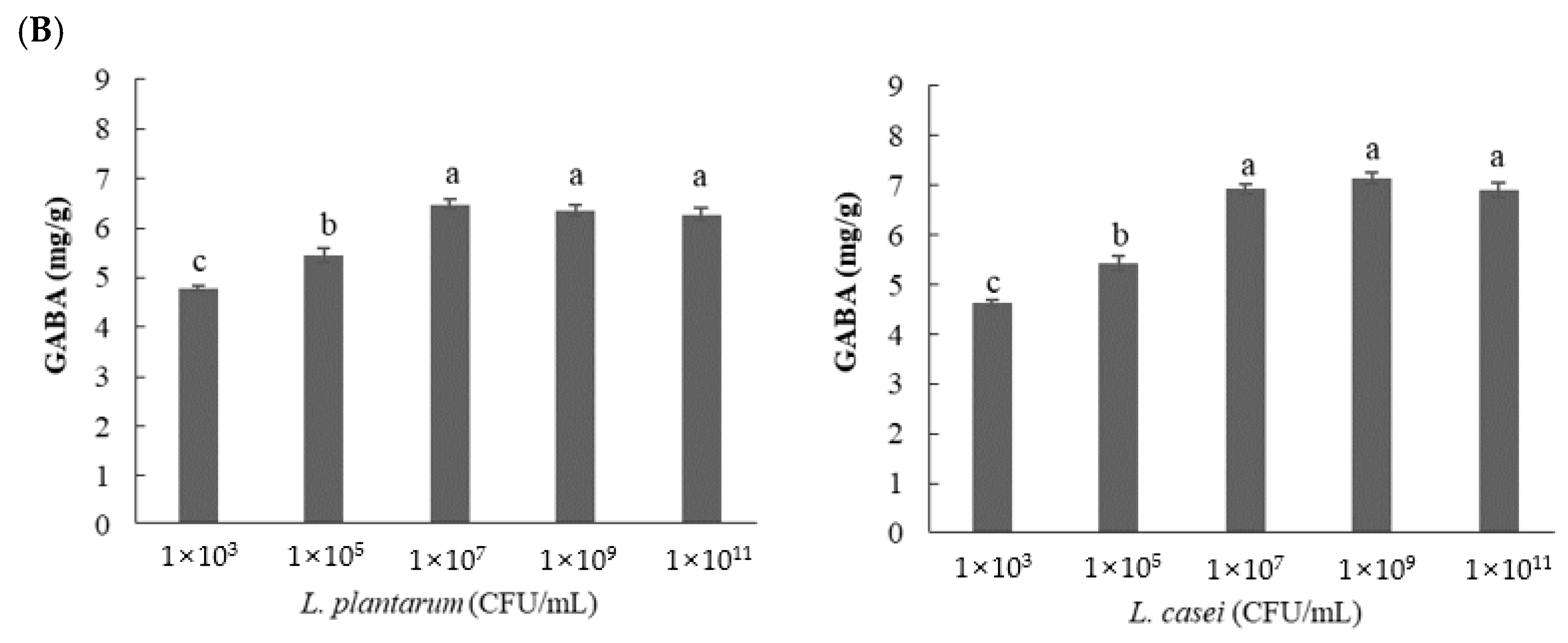
3.2.3. Effect of Inoculated Cell Number on GABA Content
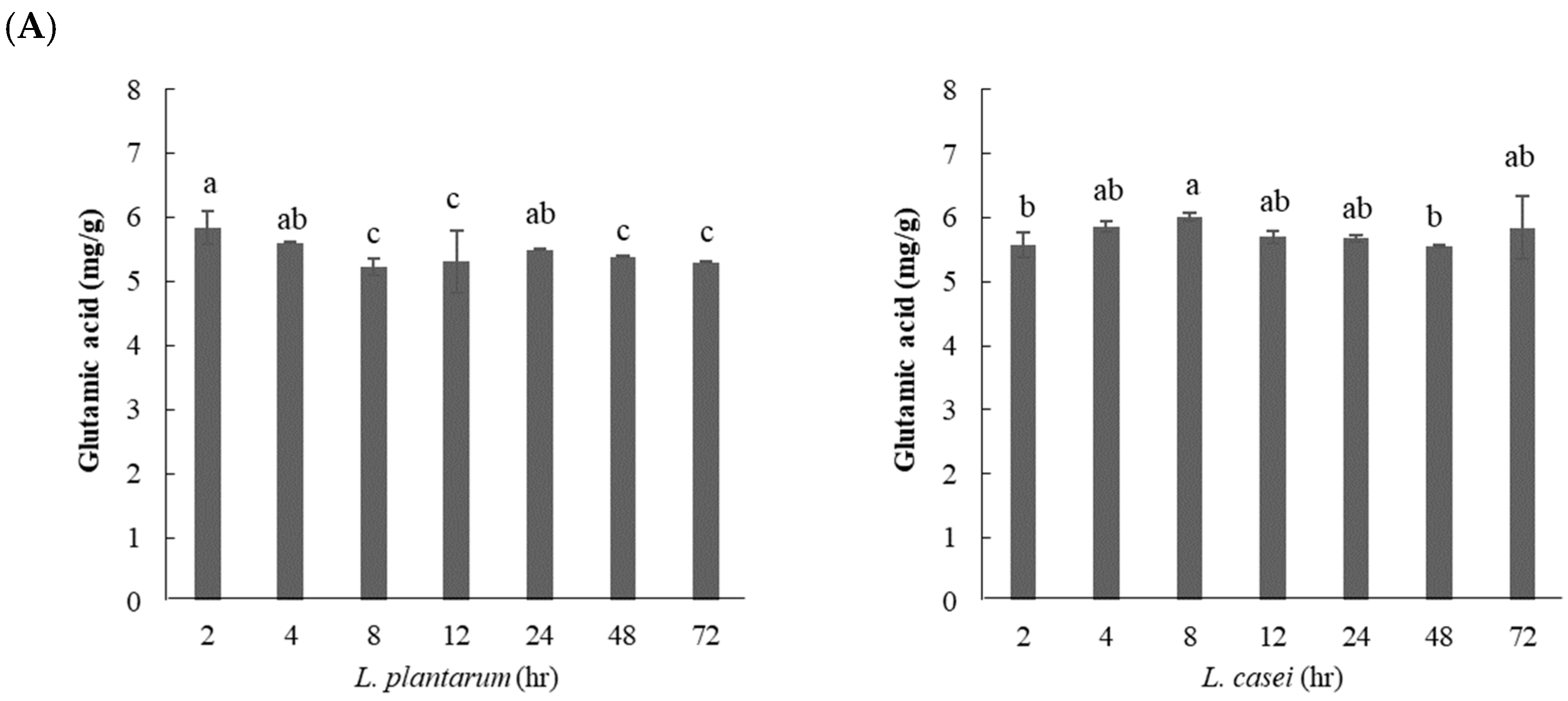
3.2.4. Effect of Fermentation Time on GABA Content
3.3. Analysis of GABA Content for Four Types of Fermented Grains
3.4. Anti-Oxidant Activities of Five Types of Fermented Grains
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ueno, H. Enzymatic and structural aspects on glutamate decarboxylase. J. Mol. Catal. B Enzym. 2000, 10, 67–79. [Google Scholar] [CrossRef]
- Checkoway, H.; Jessica, I.L.; Samir, N.K. Neurodegenerative diseases. IARC Sci. Publ. 2011, 163, 407–419. [Google Scholar]
- Neelam, K.; Duddu, V.; Anyim, N.; Neelam, J.; Lewis, S. Pandemics and pre-existing mental illness: A systematic review and meta-analysis. Brain Behav. Immun. 2021, 10, 100177. [Google Scholar] [CrossRef] [PubMed]
- Sengoku, R. Aging and Alzheimer’s disease pathology. Neuropathology 2020, 40, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Soghomonian, J.J.; David, L.M. Two isoforms of glutamate decarboxylase: Why? Trends Pharmacol. Sci. 1998, 19, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.D.; Jeng, L.M.; Ching, A.H. Bioactive components and antioxidant properties of γ-aminobutyric acid (GABA) tea leaves. LWT-Food Sci. Technol. 2012, 46, 64–70. [Google Scholar] [CrossRef]
- Farahmandfar, M.; Zarrindast, M.R.; Kadivar, M.; Karimian, S.M.; Naghdi, N. The effect of morphine sensitization on extracellular concentrations of GABA in dorsal hippocampus of male rats. Eur. J. Pharmacol. 2011, 669, 66–70. [Google Scholar] [CrossRef]
- Buzzi, A.; Chikhladze, M.; Falcicchia, C.; Paradiso, B.; Lanza, G.; Soukupova, M.; Marti, M.; Morari, M.; Franceschetti, S.; Simonato, M. Loss of cortical GABA terminals in Unverricht Lundborg disease. Neurobiol. Dis. 2012, 47, 216–224. [Google Scholar] [CrossRef]
- Leventhal, A.G.; Wang, Y.C.; Pu, M.L.; Zhou, Y.F.; Ma, Y. GABA and its agonists improved visual cortical function in senescent monkeys. Science 2003, 300, 812–815. [Google Scholar] [CrossRef]
- Seok, H.M. Functional Description of GABA and Possibility of New Material Development. J. Food Technol. 2002, 15, 81–85. [Google Scholar]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Gironell, A.; Figueiras, F.P.; Pagonabarraga, J.; Herance, J.R.; Pascual-Sedano, B.; Trampal, C.; Gispert, J.D. GABA and serotonin molecular neuroimaging in essential tremor: A clinical correlation study. Parkinsonism Relat. Disord. 2012, 18, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yang, T.; Feng, H.; Dong, M.; Slavin, M.; Xiong, S.; Zhao, S. Enhancing contents of γ-aminobutyric acid (GABA) and other micronutrients in dehulled rice during germination under normoxic and hypoxic conditions. J. Agric. Food Chem. 2016, 64, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Han, S.I.; Ra, J.E.; Seo, K.H.; Park, J.Y.; Seo, W.D.; Park, D.S.; Cho, J.H.; Lee, J.H.; Sim, E.Y.; Nam, M.H. Effects of physico-chemical treatment on ‘Nunkeunhukchal’ (black sticky rice with giant embryo) for the enhancement of GABA (γ-aminobutyric acid) contents. Korean J. Crop. Sci. 2014, 59, 398–405. [Google Scholar] [CrossRef]
- Zushi, K.; Naotaka, M. Salt stress-enhanced γ-aminobutyric acid (GABA) in tomato fruit. Acta Hort. 2007, 761, 431–436. [Google Scholar] [CrossRef]
- Nejad, A.F.; Nasibi, F.; Kalantari, K.M. 24-epibrassinolide pre-treatment alleviates the salt-induced deleterious effects in medicinal pumpkin (Cucurbita pepo) by enhancement of GABA content and enzymatic antioxidants. S. Afr. J. Bot. 2019, 124, 111–117. [Google Scholar] [CrossRef]
- Sun, X.; Wang, J.; Li, C.; Zheng, M.; Zhang, Q.; Xiang, W.; Tang, J. The use of γ-aminobutyric acid-producing saccharomyces cerevisiae SC125 for functional fermented beverage production from apple juice. Foods 2022, 11, 1202. [Google Scholar] [CrossRef]
- Ly, D.; Mayrhofer, S.; Yogeswara, I.B.A.; Nguyen, T.-H.; Domig, K.J. Identification, classification and screening for γ-amino-butyric acid production in lactic acid bacteria from Cambodian fermented foods. Biomolecules 2019, 9, 768. [Google Scholar] [CrossRef] [Green Version]
- Ueno, Y.; Hayakawa, K.; Takahashi, S.; Oda, K. Purification and characterization of glutamate decarboxylase from Lactobacillus brevis IFO 12005. Biosci. Biotechnol. Biochem. 1997, 61, 1168–1171. [Google Scholar] [CrossRef]
- Higuchi, T.; Hisanobu, H.; Keietsu, A. Exchange of glutamate and gamma-aminobutyrate in a Lactobacillus strain. J. Bacteriol. Res. 1997, 179, 3362–3364. [Google Scholar] [CrossRef] [Green Version]
- Park, K.B.; Oh, S.H. Production of yogurt with enhanced levels of gamma-aminobutyric acid and valuable nutrients using lactic acid bacteria and germinated soybean extract. Bioresour. Technol. 2007, 98, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Han, M.H.; Yang, J.M.; Joo, Y.C.; Moon, G.S. Production of functional fermented soymilk by γ-aminobutyric acid producing lactic acid bacteria. Curr. Top. Lact. Acid Bact. Probiotics 2019, 5, 59–64. [Google Scholar] [CrossRef]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Jeong, M.H.; Ahn, C.B.; Je, J.Y. Antioxidant activity and γ- aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A. Solid-state fermentation. Biochem. Eng. J. 2003, 13, 81–84. [Google Scholar] [CrossRef]
- Thomas, L.; Christian, L.; Ashok, P. Current developments in solid-state fermentation. Biochem. Eng. J. 2013, 81, 146–161. [Google Scholar] [CrossRef]
- Guardado-Félix, D.; Antunes-Ricardo, M.; Rocha-Pizaña, M.R.; Martínez-Torres, A.C.; Gutiérrez-Uribe, J.A.; Saldivar, S.O.S. Chickpea (Cicer arietinum L.) sprouts containing supranutritional levels of selenium decrease tumor growth of colon cancer cells xenografted in immune-suppressed mice. J. Funct. Foods 2019, 53, 76–84. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, J.Y.; Kim, K.W.; Yoon, K.Y. Nutritional com- position and functionality of mixed cereals powder. Korean J. Food Preserv. 2014, 54, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Amarowicz, R.; Estrella, I.; Hernández, T.; Robredo, S.; Troszyńska, A.; Kosińska, A.; Pegg, R.B. Free radical-scavenging capacity, antioxidant activity, and phenolic composition of green lentil (Lens culinaris). Food chem. 2010, 121, 705–711. [Google Scholar] [CrossRef]
- Lee, H.K.; Hwang, I.G.; Kim, H.Y.; Woo, K.S.; Lee, S.H.; Woo, S.H.; Lee, J.S.; Jeong, H.S. Physicochemical characteristic and antioxidant activities of cereals and legumes in Korea. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1399–1404. [Google Scholar] [CrossRef]
- Lu, Q.; Goto, K.; Nishizu, T. Study on the conditions and quality of GABA enriched brown rice. J. JSAM 2010, 72, 291–296. [Google Scholar]
- Ma, Y.; Wang, A.; Yang, M.; Wang, S.; Wang, L.; Zhou, S.; Blecker, C. Influences of cooking and storage on γ-aminobutyric acid (GABA) content and distribution in mung bean and its noodle products. LWT 2022, 154, 112783. [Google Scholar] [CrossRef]
- Yun, I.J. Effect of Selection Attributes of Convenience Food Instant Japgog (Mixed Cereals) Rice on Customer Satisfaction and Loyalty. Ph.D. Thesis, Catholic Kwandong University, Gangneung, Republic of Korea, 2016. [Google Scholar]
- Nongsaro. Available online: https://www.nongsaro.go.kr/portal/ps/pst/pstb/pstbc/mngmtDtaDtl.ps?menuId=PS03213&nttSn=694&totalSearchYn=Y (accessed on 1 September 2017).
- Kim, S.I.; Kang, J.H.; Lee, J.S.; Lee, J.S. Effect on rice image for connectedness formation and meal culture settlement. J. Foodserv. Manag. 2016, 19, 25–49. [Google Scholar]
- Kim, H.S.; Oh, I.K.; Yang, S.K.; Lee, S.Y. A comparison of rheological measurement methods of instant cooked rice by a texture analyzer. Food Eng. Prog. 2018, 22, 381–385. [Google Scholar] [CrossRef]
- Jung, Y.J.; Cho, Y.J.; Kim, K.W.; Yoon, K.Y. Current status and development plan of domestic cereal industry. Food Preserv. Process. Indust. 2013, 12, 31–39. [Google Scholar]
- Panrod, K.; Anyarporn, T.; Vipaporn, P. Comparison of validated high performance liquid chromatography methods using two derivatizing agents for gamma-aminobutyric acid quantification. Thai J. Pharm. Sci. 2016, 40, 203–208. [Google Scholar]
- Hayat, A.; Jahangir, T.M.; Khuhawar, M.Y.; Alamgir, M.; Siddiqui, A.J.; Musharraf, S.G. Simultaneous HPLC determination of gamma amino butyric acid (GABA) and lysine in selected Pakistani rice varieties by pre-column derivatization with 2-Hydroxynaphthaldehyde. J. Cereal Sci. 2014, 60, 356–360. [Google Scholar] [CrossRef]
- Jang, G.W.; Choi, S.I.; Han, X.; Men, X.; Kwon, H.Y.; Choi, Y.E.; Kang, N.Y.; Park, B.W.; Kim, J.J.; Lee, O.H. Antioxidant and Anti-inflammatory Activities of Phellodendron amurense Extract Fermented with Lactobacillus plantarum CM. J. Food Hyg. Saf. 2021, 36, 196–203. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Radhika, D.; Bajpai, V.K.; Baek, K.H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar]
- Das, D.; Goyal, A. Antioxidant activity and γ-aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. LWT-Food Sci. Technol. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Wang, H.K.; Dong, C.; Chen, Y.F.; Cui, L.M.; Zhang, H.P. A new probiotic cheddar cheese with high ACE-Inhibitory activity and g-aminobutyric acid content produced with koumiss-derived Lactobacillus casei Zhang. Food Technol. Biotechnol. 2010, 48, 62–70. [Google Scholar]
- Rayavarapu, B.; Tallapragada, P.; Usha, M.S. Statistical optimization of γ-aminobutyric acid production by response surface methodology and artificial neural network models using Lactobacillus fermentum isolated from palm wine. Biocatal. Agric. Biotechnol. 2019, 22, 101362. [Google Scholar] [CrossRef]
- Lin, Q. Submerged fermentation of Lactobacillus rhamnosus YS9 for γ-aminobutyric acid (GABA) production. Braz. J. Microbiol. 2013, 44, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batch fermentation. Microb. Cell Fact. 2010, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef]
- Hwang, E.Y.; Park, J.Y. Isolation and characterization of gamma-aminobutyric acid (GABA)-producing lactic Acid Bacteria from Kimchi. Curr. Top. Lact. Acid Bact. Probiotics 2020, 6, 64–69. [Google Scholar] [CrossRef]
- Singh, R.; De, S.; Belkheir, A. Avena sativa (Oat), a potential neutraceutical and therapeutic agent: An overview. Crit. Rev. Food Sci. Nutr. 2013, 53, 126–144. [Google Scholar] [CrossRef]
- Xu, D.; Ren, G.Y.; Liu, L.L.; Zhu, W.X.; Liu, Y.H. The influences of drying process on crude protein content of naked oat cut herbage (Avena nuda L.). Dry Technol. 2014, 32, 321–332. [Google Scholar]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.L.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [Green Version]
- Joyce, B.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar]
- Jarpa, P.M. Lentil protein: A review of functional properties and food application. An overview of lentil protein functionality. Int. J. Food Sci. 2018, 53, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Wang, N. Optimization of a laboratory dehulling process for lentil (Lens culinaris). Cereal Chem. 2005, 82, 671–676. [Google Scholar] [CrossRef]
- Thavarajah, D.; Thavarajah, P.; Sarker, A.; Vandenberg, A. Lentils (Lens culinaris Medikus Subspecies culinaris): A whole food for increased iron and zinc intake. J. Agric. Food Chem. 2009, 57, 5413–5419. [Google Scholar] [CrossRef] [PubMed]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH. free radical method. LWT-Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- van den Berg, R.; Haenen, G.R.; van den Berg, H.; Bast, A.A.L.T. Applicability of an improved Trolox equivalent antioxidant capacity (TEAC) assay for evaluation of antioxidant capacity measurements of mixtures. Food Chem. 1999, 66, 511–517. [Google Scholar] [CrossRef]
- Ghamry, M.; Ghazal, A.F.; Al-Maqtqri, Q.A.; Li, L.; Zhao, W. Impact of a novel probiotic Lactobacillus strain isolated from the bee gut on GABA content, antioxidant activity, and potential cytotoxic activity against HT-29 cell line of rice bran. J. Food Sci. Technol. 2022, 59, 3031–3042. [Google Scholar] [CrossRef]
- Ghamry, M.; Zhao, W.; Li, L. Impact of Lactobacillus apis on the antioxidant activity, phytic acid degradation, nutraceutical value and flavor properties of fermented wheat bran, compared to Saccharomyces cerevisiae and Lactobacillus plantarum. Food Res. Int. 2022, 163, 112142. [Google Scholar] [CrossRef]
- Nisa, K.; Rosyida, V.T.; Nurhayati, S.; Indrianingsih, A.W.; Darsih, C.; Apriyana, W. Total phenolic contents and antioxidant activity of rice bran fermented with lactic acid bacteria. IOP Conf. Ser. Earth Environ. Sci. 2019, 251, 012020. [Google Scholar] [CrossRef]
- Jhan, J.K.; Chang, W.F.; Wang, P.M.; Chou, S.T.; Chung, Y.C. Production of fermented red beans with multiple bioactivities using co-cultures of Bacillus subtilis and Lactobacillus delbrueckii subsp. bulgaricus. LWT-Food Sci. Technol. 2015, 63, 1281–1287. [Google Scholar] [CrossRef]
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Lactobacillus plantarum WCFS1 fermentation differentially affects antioxidant capacity and polyphenol content in mung bean (Vigna radiata) and soya bean (Glycine max) milks. J. Food Process Preserv. 2017, 41, e12944. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instrument | Conditions |
|---|---|
| Column | Symmetry C18 (4.6 × 250 mm, 5 um), Waters Corporation |
| Column temp. | 35 °C |
| Mobile phase (isocratic) | 100 mM acetate buffer (in water): Methanol = 60: 40, v/v |
| Detector | UV detector (235 nm) |
| Flow rate | 0.8 mL/min |
| Injection volume | 10 μL |
| Run time | 50 min |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, H.-y.; Choi, J.-s.; Kim, S.-j.; Kim, E.-m.; Uhm, J.-h.; Kim, B.-k.; Lee, J.-y.; Kim, Y.-d.; Hwang, K.-t. Optimization of Solid-Phase Lactobacillus Fermentation Conditions to Increase γ-Aminobutyric Acid (GABA) Content in Selected Substrates. Fermentation 2023, 9, 22. https://doi.org/10.3390/fermentation9010022
Kwon H-y, Choi J-s, Kim S-j, Kim E-m, Uhm J-h, Kim B-k, Lee J-y, Kim Y-d, Hwang K-t. Optimization of Solid-Phase Lactobacillus Fermentation Conditions to Increase γ-Aminobutyric Acid (GABA) Content in Selected Substrates. Fermentation. 2023; 9(1):22. https://doi.org/10.3390/fermentation9010022
Chicago/Turabian StyleKwon, Hee-yeon, Ji-soo Choi, Soo-jin Kim, Eun-min Kim, Ji-hyun Uhm, Bo-kyung Kim, Jae-yeon Lee, Yong-deok Kim, and Kwon-tack Hwang. 2023. "Optimization of Solid-Phase Lactobacillus Fermentation Conditions to Increase γ-Aminobutyric Acid (GABA) Content in Selected Substrates" Fermentation 9, no. 1: 22. https://doi.org/10.3390/fermentation9010022