

Antioxidant and Functional Features of Pre-Fermented Ingredients Obtained by the Fermentation of Milling By-Products
 ,
,  ,
,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Material: Wheat Germ and Rye Bran
2.2. Microbial Consortium
2.3. Fermentation Process
- Milling by-product mixture fermented by the microbial consortium of yeasts and LAB (FM),
- Milling by-product mixture fermented by a commercial bakery yeast (benchmark)
- Unfermented milling by-product mixture (UF).
2.4. Acidification Kinetics and Plate Count
2.5. Nutritional Profile
2.6. Color Analysis
2.7. Volatile Molecule Profiles and Short-Chain Fatty Acids
2.8. Fatty Acid Profiles
2.9. Peptide and Phytate Content
2.10. Antioxidant Activity and Phenol Content
2.11. Prebiotic Activity
2.12. Antimicrobial Activity against Gastrointestinal Pathogens
2.13. In Vitro Digestion, Cell Culture and Viability Assay
2.14. Statistical Analysis
3. Results and Discussion
3.1. Fermentation Kinetics and Microbial Characteristics
3.2. Nutritional Profile and Fatty Acid Composition
3.3. Color Analysis
3.4. Volatile Molecule Profiles
3.5. Functionality
3.6. Prebiotic Activity
3.7. Antimicrobial Activity against Gastrointestinal Pathogens
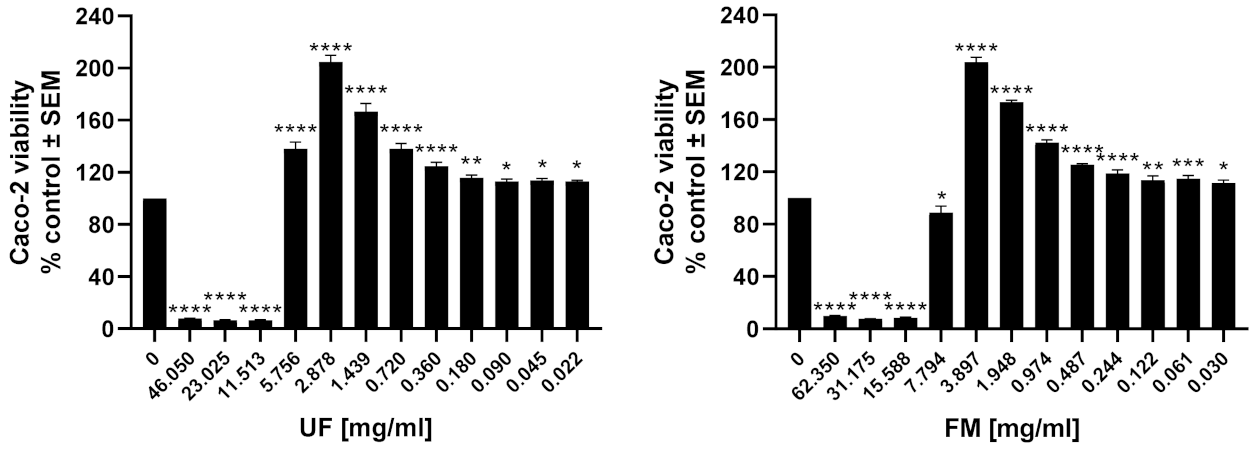
3.8. Caco-2 Viability Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Cereal Supply and Demand Brief. World Food Situation. Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 14 November 2022).
- Verni, M.; Rizzello, C.G.; Coda, R. Fermentation Biotechnology Applied to Cereal Industry By-Products: Nutritional and Functional Insights. Front. Nutr. 2019, 6, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamaratskaia, G.; Gerhardt, K.; Wendin, K. Biochemical Characteristics and Potential Applications of Ancient Cereals—An Underexploited Opportunity for Sustainable Production and Consumption. Trends. Food Sci. Technol. 2021, 107, 114–123. [Google Scholar] [CrossRef]
- Coda, R.; Kärki, I.; Nordlund, E.; Heiniö, R.L.; Poutanen, K.; Katina, K. Influence of Particle Size on Bioprocess Induced Changes on Technological Functionality of Wheat Bran. Food Microbiol. 2014, 37, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; de Angelis, M.; di Cagno, R.; Polo, A.; Rizzello, C.G. The Sourdough Fermentation Is the Powerful Process to Exploit the Potential of Legumes, Pseudo-Cereals and Milling by-Products in Baking Industry. Crit. Rev. Food Sci. Nutr. 2019, 60, 2158–2173. [Google Scholar] [CrossRef] [PubMed]
- Onipe, O.O.; Jideani, A.I.O.; Beswa, D. Composition and Functionality of Wheat Bran and Its Application in Some Cereal Food Products. Int. J. Food Sci. Technol. 2015, 50, 2509–2518. [Google Scholar] [CrossRef]
- Prückler, M.; Siebenhandl-Ehn, S.; Apprich, S.; Höltinger, S.; Haas, C.; Schmid, E.; Kneifel, W. Wheat Bran-Based Biorefinery 1: Composition of Wheat Bran and Strategies of Functionalization. LWT.-Food Sci. Technol. 2014, 56, 211–221. [Google Scholar] [CrossRef]
- Coda, R.; Katina, K.; Rizzello, C.G. Bran Bioprocessing for Enhanced Functional Properties. Curr. Opin. Food Sci. 2015, 1, 50–55. [Google Scholar] [CrossRef]
- Luithui, Y.; Baghya Nisha, R.; Meera, M.S. Cereal By-Products as an Important Functional Ingredient: Effect of Processing. J. Food Sci. Technol. 2019, 56, 1–11. [Google Scholar] [CrossRef]
- Galanakis, C.M. Sustainable Applications for the Valorization of Cereal Processing By-Products. Foods 2022, 11, 241. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, M.; Zhao, Y.; Zhu, Y.; Bai, J.; Fan, S.; Zhu, L.; Song, C.; Xiao, X. Recent Developments in Fermented Cereals on Nutritional Constituents and Potential Health Benefits. Foods 2022, 11, 2243. [Google Scholar] [CrossRef]
- Dziki, D. Rye Flour and Rye Bran: New Perspectives for Use. Processes 2022, 10, 293. [Google Scholar] [CrossRef]
- Boukid, F.; Folloni, S.; Ranieri, R.; Vittadini, E. A Compendium of Wheat Germ: Separation, Stabilization and Food Applications. Trends. Food Sci. Technol. 2018, 78, 120–133. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; de Angelis, M.; Gobbetti, M. Effect of Sourdough Fermentation on Stabilisation, and Chemical and Nutritional Characteristics of Wheat Germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; di Cagno, R.; Gobbetti, M. Use of Sourdough Fermented Wheat Germ for Enhancing the Nutritional, Texture and Sensory Characteristics of the White Bread. Eur. Food Res. Technol. 2010, 230, 645–654. [Google Scholar] [CrossRef]
- Onipe, O.O.; Ramashia, S.E.; Jideani, A.I.O. Wheat Bran Modifications for Enhanced Nutrition and Functionality in Selected Food Products. Molecules 2021, 26, 3918. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, A.; Roy, D.; LaPointe, G. Fermentation of Wheat Bran and Whey Permeate by Mono-Cultures of Lacticaseibacillus Rhamnosus Strains and Co-Culture With Yeast Enhances Bioactive Properties. Front. Bioeng. Biotechnol. 2020, 8, 956. [Google Scholar] [CrossRef]
- Filannino, P.; di Cagno, R.; Gobbetti, M. Metabolic and Functional Paths of Lactic Acid Bacteria in Plant Foods: Get out of the Labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]
- Arte, E.; Rizzello, C.G.; Verni, M.; Nordlund, E.; Katina, K.; Coda, R. Impact of Enzymatic and Microbial Bioprocessing on Protein Modification and Nutritional Properties of Wheat Bran. J. Agric. Food Chem. 2015, 63, 8685–8693. [Google Scholar] [CrossRef]
- Arora, K.; Ameur, H.; Polo, A.; di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty Years of Knowledge on Sourdough Fermentation: A Systematic Review. Trends. Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Portincasa, P.; Montemurro, M.; di Palo, D.M.; Lorusso, M.P.; de Angelis, M.; Bonfrate, L.; Genot, B.; Gobbetti, M. Sourdough Fermented Breads Are More Digestible than Those Started with Baker’s Yeast Alone: An In Vivo Challenge Dissecting Distinct Gastrointestinal Responses. Nutrients 2019, 11, 2954. [Google Scholar] [CrossRef]
- Khalid, K.H.; Ohm, J.B.; Simsek, S. Whole Wheat Bread: Effect of Bran Fractions on Dough and End-Product Quality. J. Cereal. Sci. 2017, 78, 48–56. [Google Scholar] [CrossRef]
- Official Methods of Analysis of AOAC International. WorldCat.Org. Available online: https://www.worldcat.org/it/title/official-methods-of-analysis-of-aoac-international/oclc/1085712083 (accessed on 11 November 2022).
- AACC. Approved Methods of the AACC, 10th ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 2000; Available online: http://www.sciepub.com/reference/241265 (accessed on 11 November 2022).
- Buddrick, O.; Jones, O.A.H.; Cornell, H.J.; Small, D.M. The Influence of Fermentation Processes and Cereal Grains in Wholegrain Bread on Reducing Phytate Content. J. Cereal. Sci. 2014, 59, 3–8. [Google Scholar] [CrossRef]
- da Rocha Lemos Mendes, G.; Souto Rodrigues, P.; de las Mercedes Salas-Mellado, M.; Fernandes de Medeiros Burkert, J.; Badiale-Furlong, E. Defatted Rice Bran as a Potential Raw Material to Improve the Nutritional and Functional Quality of Cakes. Plant. Foods Hum. Nutr. 2021, 76, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Aoac 920.39 2003_05 Extracto Etereo Dietileter-VSIP. INFO. Available online: https://vsip.info/aoac-92039-200305-extracto-etereo-dietileter-2-pdf-free.html (accessed on 11 November 2022).
- Oliveira, M.D.S.; Feddern, V.; Kupski, L.; Cipolatti, E.P.; Badiale-Furlong, E.; de Souza-Soares, L.A. Changes in Lipid, Fatty Acids and Phospholipids Composition of Whole Rice Bran after Solid-State Fungal Fermentation. Bioresour. Technol. 2011, 102, 8335–8338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, L.; Lukšič, L.; Molinari, R.; Kreft, I.; Bonafaccia, G.; Manzi, L.; Merendino, N. Development of Gluten-Free Bread Using Tartary Buckwheat and Chia Flour Rich in Flavonoids and Omega-3 Fatty Acids as Ingredients. Food Chem. 2014, 165, 232–240. [Google Scholar] [CrossRef]
- Luchese, C.L.; Gurak, P.D.; Marczak, L.D.F. Osmotic Dehydration of Physalis (Physalis peruviana L.): Evaluation of Water Loss and Sucrose Incorporation and the Quantification of Carotenoids. LWT.-Food Sci. Technol. 2015, 63, 1128–1136. [Google Scholar] [CrossRef]
- Pointer, M.R. A Comparison of the CIE 1976 Colour Spaces. Color. Res. Appl. 1981, 6, 108–118. [Google Scholar] [CrossRef]
- Burns, P.; Patrignani, F.; Serrazanetti, D.; Vinderola, G.C.; Reinheimer, J.A.; Lanciotti, R.; Guerzoni, M.E. Probiotic Crescenza Cheese Containing Lactobacillus Casei and Lactobacillus Acidophilus Manufactured with High-Pressure Homogenized Milk. J. Dairy Sci. 2008, 91, 500–512. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Parrotta, L.; del Duca, S.; Rosa, M.D.; Patrignani, F.; Schluter, O.; Lanciotti, R. Effect of Yarrowia Lipolytica RO25 Cricket-Based Hydrolysates on Sourdough Quality Parameters. LWT 2021, 148, 111760. [Google Scholar] [CrossRef]
- Boselli, E.; Velazco, V.; Fiorenza Caboni, M.; Lercker, G. Pressurized Liquid Extraction of Lipids for the Determination of Oxysterols in Egg-Containing Food. J. Chromatogr. A. 2001, 917, 239–244. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected Lactic Acid Bacteria Synthesize Antioxidant Peptides during Sourdough Fermentation of Cereal Flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC 986.11-1988, Phytate in Foods Anion Exchange Method-$14.15: AOAC Official Method. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&cPath=1&products_id=313 (accessed on 11 November 2022).
- Rizzello, C.G.; Coda, R.; Mazzacane, F.; Minervini, D.; Gobbetti, M. Micronized By-Products from Debranned Durum Wheat and Sourdough Fermentation Enhanced the Nutritional, Textural and Sensory Features of Bread. Food Res. Int. 2012, 46, 304–313. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors Influencing the Antioxidant Activity Determined by the ABTS Radical Cation Assay. Free. Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Slinkard, K.; Singleton, V.L. Total Phenol Analysis: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Huebner, J.; Wehling, R.L.; Hutkins, R.W. Functional Activity of Commercial Prebiotics. Int. Dairy. J. 2007, 17, 770–775. [Google Scholar] [CrossRef]
- Giordani, B.; Melgoza, L.M.; Parolin, C.; Foschi, C.; Marangoni, A.; Abruzzo, A.; Dalena, F.; Cerchiara, T.; Bigucci, F.; Luppi, B.; et al. Vaginal Bifidobacterium Breve for Preventing Urogenital Infections: Development of Delayed Release Mucoadhesive Oral Tablets. Int. J. Pharm. 2018, 550, 455–462. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Static in Vitro Digestion Method Suitable for Food—An International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Scarnato, L.; Serrazanetti, D.I.; Aloisi, I.; Montanari, ·C.; del Duca, S.; Lanciotti, R. Combination of Transglutaminase and Sourdough on Gluten-Free Flours to Improve Dough Structure. Amino. Acids. 2016, 48, 2453–2465. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; di Cagno, R.; de Angelis, M. How the Sourdough May Affect the Functional Features of Leavened Baked Goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- de Luca, L.; Aiello, A.; Pizzolongo, F.; Blaiotta, G.; Aponte, M.; Romano, R. Volatile Organic Compounds in Breads Prepared with Different Sourdoughs. Applied. Sci. 2021, 11, 1330. [Google Scholar] [CrossRef]
- Scarnato, L.; Montanari, C.; Serrazanetti, D.I.; Aloisi, I.; Balestra, F.; del Duca, S.; Lanciotti, R. New Bread Formulation with Improved Rheological Properties and Longer Shelf-Life by the Combined Use of Transglutaminase and Sourdough. LWT 2017, 81, 101–110. [Google Scholar] [CrossRef]
- Manini, F.; Brasca, M.; Plumed-Ferrer, C.; Morandi, S.; Erba, D.; Casiraghi, M.C. Study of the Chemical Changes and Evolution of Microbiota During Sourdoughlike Fermentation of Wheat Bran; Study of the Chemical Changes and Evolution of Microbiota During Sourdoughlike Fermentation of Wheat Bran. Cereal Chem. 2014, 91, 342–349. [Google Scholar] [CrossRef]
- Zhao, H.M.; Guo, X.N.; Zhu, K.X. Impact of Solid State Fermentation on Nutritional, Physical and Flavor Properties of Wheat Bran. Food Chem. 2017, 217, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [Green Version]
- Decimo, M.; Quattrini, M.; Ricci, G.; Fortina, M.G.; Brasca, M.; Silvetti, T.; Manini, F.; Erba, D.; Criscuoli, F.; Casiraghi, M.C. Evaluation of Microbial Consortia and Chemical Changes in Spontaneous Maize Bran Fermentation. AMB Express. 2017, 7, 205. [Google Scholar] [CrossRef]
- Ndagijimana, M.; Vallicelli, M.; Cocconcelli, P.S.; Cappa, F.; Patrignani, F.; Lanciotti, R.; Guerzoni, M.E. Two 2[5H]-Furanones as Possible Signaling Molecules in Lactobacillus Helveticus. Appl. Environ. Microbiol. 2006, 72, 6053–6061. [Google Scholar] [CrossRef] [Green Version]
- Kaseleht, K.; Paalme, T.; Mihhalevski, A.; Sarand, I. Analysis of Volatile Compounds Produced by Different Species of Lactobacilli in Rye Sourdough Using Multiple Headspace Extraction. Int. J. Food Sci. Technol. 2011, 46, 1940–1946. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y.; Tang, K.; Hu, Y.; Xu, X.; Gänzle, M.G. Effect of Mixed Cultures of Yeast and Lactobacilli on the Quality of Wheat Sourdough Bread. Front. Microbiol. 2019, 10, 2113. [Google Scholar] [CrossRef] [Green Version]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Fluorescence Labeling of Wheat Proteins for Determination of Gluten Hydrolysis and Depolymerization during Dough Processing and Sourdough Fermentation. J. Agric. Food Chem. 2003, 51, 2745–2752. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Zheng, J. Lifestyles of Sourdough Lactobacilli—Do They Matter for Microbial Ecology and Bread Quality? Int. J. Food Microbiol. 2019, 302, 15–23. [Google Scholar] [CrossRef]
- Pérez-Alvarado, O.; Zepeda-Hernández, A.; Garcia-Amezquita, L.E.; Requena, T.; Vinderola, G.; García-Cayuela, T. Role of Lactic Acid Bacteria and Yeasts in Sourdough Fermentation during Breadmaking: Evaluation of Postbiotic-like Components and Health Benefits. Front. Microbiol. 2022, 13, 969460. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial Effects on Host Energy Metabolism of Short-Chain Fatty Acids and Vitamins Produced by Commensal and Probiotic Bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziata, G.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Novellino, E. Fermentation of Foods and Beverages as a Tool for Increasing Availability of Bioactive Compounds. Focus on Short-Chain Fatty Acids. Foods 2020, 9, 999. [Google Scholar] [CrossRef] [PubMed]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary Gut Microbial Metabolites, Short-Chain Fatty Acids, and Host Metabolic Regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birch, A.N.; Petersen, M.A.; Hansen, Å.S. The Aroma Profile of Wheat Bread Crumb Influenced by Yeast Concentration and Fermentation Temperature. LWT.-Food Sci. Technol. 2013, 50, 480–488. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food. Fermentation 2019, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Pessione, E.; Cirrincione, S. Bioactive Molecules Released in Food by Lactic Acid Bacteria: Encrypted Peptides and Biogenic Amines. Front. Microbiol. 2016, 7, 876. [Google Scholar] [CrossRef] [Green Version]
- Liukkonen, K.-H.; Katina, K.; Wilhelmsson, A.; Myllymaki, O.; Lampi, A.-M.; Kariluoto, S.; Piironen, V.; Heinonen, S.-M.; Nurmi, T.; Adlercreutz, H.; et al. Process-Induced Changes on Bioactive Compounds in Whole Grain Rye. Proc. Nutr. Soc. 2003, 62, 117–122. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I.G. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.; Cheng, Z.; Hao, J.; Guo, G.; Liu, J.G.; Lin, C.; Yu, L. Effects of Solid-State Yeast Treatment on the Antioxidant Properties and Protein and Fiber Compositions of Common Hard Wheat Bran. J. Agric. Food Chem. 2007, 55, 10173–10182. [Google Scholar] [CrossRef]
- Spaggiari, M.; Ricci, A.; Calani, L.; Bresciani, L.; Neviani, E.; Dall’Asta, C.; Lazzi, C.; Galaverna, G. Solid State Lactic Acid Fermentation: A Strategy to Improve Wheat Bran Functionality. LWT 2020, 118, 108668. [Google Scholar] [CrossRef]
- Laurent-Babot, C.; Guyot, J.P. Should Research on the Nutritional Potential and Health Benefits of Fermented Cereals Focus More on the General Health Status of Populations in Developing Countries? Microorganisms 2017, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and Cereal Fermentation in a Nutritional Perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef]
- Brown, L.; Pingitore, E.V.; Mozzi, F.; Saavedra, L.M.; Villegas, J.M.; Hebert, E. Lactic Acid Bacteria as Cell Factories for the Generation of Bioactive Peptides. Protein Pept. Lett. 2017, 24, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and Medium-Chain Fatty Acids Exhibit Antimicrobial Activity for Oral Microorganisms. Arch. Oral. Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Gong, S.; Wang, Y.; Sun, Q.; Duo, K.; Fei, P. Antibacterial Activity of Olive Oil Polyphenol Extract Against Salmonella Typhimurium and Staphylococcus Aureus: Possible Mechanisms. Foodborne. Pathog. Dis. 2020, 17, 396–403. [Google Scholar] [CrossRef]
- Tang, A.S.; Chikhale, P.J.; Shah, P.K.; Borchardt, R.T. Utilization of a Human Intestinal Epithelial Cell Culture System (Caco-2) for Evaluating Cytoprotective Agents. Pharm. Res. 1993, 10, 1620–1626. [Google Scholar] [CrossRef]
- Bruininx, E.M.A.M.; Koninkx, J.F.J.G.; Binnendijk, G.P.; Zandstra, T.; Heetkamp, M.J.W.; van der Peet-Schwering, C.M.C.; Gerrits, W.J.J. Effects of Prefermented Cereals or the End Products of Fermentation on Growth and Metabolism of Enterocyte-like Caco-2 Cells and on Intestinal Health of Restrictedly Fed Weanling Pigs. Animal 2010, 4, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Galli, V.; Mazzoli, L.; Luti, S.; Venturi, M.; Guerrini, S.; Paoli, P.; Vincenzini, M.; Granchi, L.; Pazzagli, L. Effect of Selected Strains of Lactobacilli on the Antioxidant and Anti-Inflammatory Properties of Sourdough. Int. J. Food Microbiol. 2018, 286, 55–65. [Google Scholar] [CrossRef]
- Yang, H.S.; Haj, F.G.; Lee, M.; Kang, I.; Zhang, G.; Lee, Y. Laminaria Japonica Extract Enhances Intestinal Barrier Function by Altering Inflammatory Response and Tight Junction-Related Protein in Lipopolysaccharide-Stimulated Caco-2 Cells. Nutrients 2019, 11, 1001. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component % | Wheat Germ | Rye Bran |
|---|---|---|
| Proteins % | 28.9 | 13.8 |
| Humidity % | 12.1 | 11.6 |
| Ashes % | 3.9 | 2.5 |
| Soluble Fiber % | 1.1 | 4.6 |
| Insoluble Fiber % | 11.9 | 18.5 |
| Total fat % | 7.5 | 2.9 |
| Starch % | 23.6 | 45.0 |
| Phytic acid % | 1.6 | 1.3 |
| Pre-Fermentation | Post-Fermentation | |||||
|---|---|---|---|---|---|---|
| Acidity | LAB | Yeasts | LAB | Yeasts | ||
| Sample | pH | meqNaOH | log CFU/g | log CFU/g | log CFU/g | log CFU/g |
| FM 1 | 3.88 ± 0.16 a | 21.10 ± 0.40 a | 6.54 ± 0.21 | 4.67 ± 0.18 a | 9.46 ± 0.19 a | 7.38 ± 0.18 a |
| Benchmark 2 | 5.69 ± 0.13 b | 9.40 ± 0.30 b | <1.0 | 6.89 ± 0.22 b | 2.11 ± 0.11 b | 8.21 ± 0.21 b |
| UF 3 | 6.43 ± 0.12 c | 6.10 ± 0.10 c | <1.0 | <1.0 | - | - |
| FM 1 | Benchmark 2 | UF 3 | |
|---|---|---|---|
| Fat g/100 g | 4.0 ± 0.5 a | 3.0 ± 0.4 a | 3.5 ± 0.3 a |
| Saturated fatty acids g/100 g | 1.2 ± 0.2 a | 0.9 ± 0.2 a | 0.9 ± 0.2 a |
| starch g/100 g | 9.1 ± 0.4 b | 9.3 ± 0.5 b | 10.5 ± 0.7 a |
| Sugars g/100 g | 0.3 ± 0.1 a | 0.3 ± 0.1 a | 2.8 ± 0.3 b |
| Insoluble fiber g/100 g | 38.8 ± 1.2 a | 41.3 ± 1.6 a | 40.5 ± 1.3 a |
| Soluble fiber g/100 g | 9.3 ± 0.4 a | 8.5 ± 0.5 ab | 8.3 ± 0.3 b |
| Protein g/100 g | 16.6 ± 0.4 a | 15.7 ± 0.5 ab | 15.3 ± 0.4 b |
| Lactic acid g/100 g | 1.011 | 0.02 | - |
| Acetic acid g/100 g | 0.104 | 0.06 | 0.02 |
| FA Relative Percentage | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C14:0 | C15:0 | C16:1 | C16:0 | C18:2 n-6 | C18:1 Δ9 | C18:3 n-3 | C18:0 | CL * | UL† | |
| UF 1 | 0.4 a | 0.3 a | 0.4 a | 37.7 b | 42.2 a | 15.1 a | 2.1 a | 1.8 a | 1722 | 1.06 |
| Benchmark 2 | 0.6 a | 0.3 a | 0.3 a | 35.5 b | 42.0 a | 16.9 a | 2.2 a | 2.2 a | 1725 | 1.08 |
| FM 3 | 0.7 a | 0.1 a | 0.8 b | 30.5 a | 40.1 a | 22.6 b | 2.8 b | 2.4 a | 1732 | 1.11 |
| L* | a* | b* | |
|---|---|---|---|
| UF | 55.32 ± 1.07 b | 0.85 ± 0.20 a | 12.45 ± 0.56 c |
| Benchmark | 52.63 ± 1.11 c | 2.62 ± 0.26 c | 8.77 ± 0.52 a |
| FM | 63.15 ± 0.92 a | 1.68 ± 0.40 b | 10.66 ± 0.39 b |
| UF | FM | Benchmark | ||
|---|---|---|---|---|
| Odor Perception a | ppm | |||
| Pentanal | Fruity | - | 0.03 | 0.02 |
| Hexanal | Green, Fruity | 0.73 | 2.24 | 3.10 |
| Heptanal | Green, Herbal | - | 0.23 | 0.54 |
| 2-Hexenal | Fruity, Green | 0.06 | 1.23 | 0.87 |
| 2-Nonenal, (E)- | Fatty, Green | 0.05 | 0.18 | 0.83 |
| Acetaldehyde | Ethereal, Pungent, Fruity | 0.24 | 1.18 | 1.01 |
| Benzaldehyde | Fruity, Sharp | - | 0.20 | 0.19 |
| Total Aldehydes | 1.08 | 5.29 | 6.56 | |
| 2,3-Butanedione | Buttery, Creamy, Pungent | - | 0.13 | 0.15 |
| 3-Pentanone | Ethereal | - | 0.44 | - |
| Methyl Isobutyl Ketone | Green, Sharp, Herbal | 0.34 | 0.16 | 0.31 |
| 2-Hexanone, 4-methyl- | Fruity, Fungal, Meaty | 0.08 | 0.11 | 0.20 |
| 3-Penten-2-one, 4-methyl- | Vegetable, Pungent | 0.40 | 0.58 | 0.42 |
| 4-Heptanone, 2,6-dimethyl- | Green, Fruity | 0.13 | 0.22 | 0.35 |
| 2-Butanone, 3-hydroxy- | Buttery, Sweet, Creamy | - | 1.23 | 0.28 |
| Total Ketones | 0.96 | 2.85 | 1.71 | |
| Ethyl Alcohol | Alcoholic, Ethereal | 0.35 | 26.71 | 40.33 |
| 1-Propanol | Alcoholic, Fermented | - | 0.20 | 0.19 |
| 1-Penten-3-ol | Green, Ethereal | - | 0.21 | 0.18 |
| 2-Hexanol | Winey, Fruity | 0.21 | 0.29 | 0.18 |
| 1-Pentanol | Fermented, Balsamic | 0.61 | 0.76 | 0.21 |
| 2-Penten-1-ol, (E)- | Green | 0.13 | 0.38 | 0.17 |
| 1-Hexanol | Herbal, Hethereal | 1.29 | 8.38 | 4.12 |
| 1-Octen-3-ol | Earthy, Mushroom | 0.19 | 0.49 | 0.09 |
| Heptanol | Green, Musty | - | 0.79 | 2.89 |
| 1-Hexanol, 2-ethyl- | Citrus, Floral | 0.06 | 0.19 | 0.10 |
| 1-Octanol | Waxy, Green | 0.06 | 0.53 | 0.47 |
| 2-Furanmethanol | Bready, Alcoholic | - | 0.06 | 0.10 |
| Benzyl Alcohol | Floral, Balsamic | - | 0.59 | 0.67 |
| 1-Butanol, 3-methyl | Fruity, Sweet | - | 1.62 | 2.88 |
| 3-Nonen-1-ol, (Z)- | Waxy, Green | - | 0.15 | 3.18 |
| Phenylethyl Alcohol | Floral | 0.08 | 0.36 | - |
| Total Alcohols | 2.96 | 41.70 | 55.67 | |
| Acetic Acid | Acidic, Sharp | 0.15 | 7.96 | 1.85 |
| Butanoic Acid | Cheesy, Sharp | - | 0.12 | - |
| Butanoic Acid, 3-methyl- | Sweet, Mentholic | - | 0.36 | - |
| Pentanoic Acid | Cheesy, Acidic | - | 0.43 | - |
| Hexanoic Acid | Fatty, Sour | 0.08 | 3.60 | 0.35 |
| Heptanoic Acid | Cheesy, Rancid, Sour | - | 0.11 | - |
| Octanoic Acid | Fatty, Waxy, Rancid | - | 0.18 | - |
| Total Acids | 0.23 | 9.76 | 2.20 | |
| Ethyl Acetate | Ethereal, Fruity, Sweet | - | 8.45 | 18.90 |
| Pentanoic Acid, Ethyl Ester | Fruity, Sweet | - | 0.41 | 1.35 |
| Hexanoic Acid, Ethyl Ester | Fruity | - | 1.21 | 5.21 |
| Heptanoic Acid, Ethyl Ester | Fruity | 0.05 | 0.05 | 0.73 |
| Octanoic Acid, Ethyl Ester | Waxy, Fruity, Winey | 0.11 | - | 0.29 |
| Nonanoic Acid, Ethyl Ester | Waxy, Fruity | - | - | 0.51 |
| Butanoic Acid, Ethyl Ester | Fruity, Juicy | - | - | 0.27 |
| 1-Butanol, 3-methyl-, Acetate | Fruity, Sweet | - | 0.15 | - |
| 1-Butanol, 3-methyl-, Formate | Green, Fruity | - | 4.19 | 6.87 |
| Total Esters | 0.16 | 14.46 | 34.13 | |
| Butylated Hydroxytoluene | Phenolic, Camphoreous | 1.13 | 1.05 | 1.05 |
| Cyclohexene, 1,6,6-trimethyl- | Citrus, Herbal | 1.59 | 1.53 | 1.19 |
| Cyclohexene, 3-methyl- | Citrus | - | 0.07 | 0.11 |
| Hexadecane | - | 0.08 | 0.02 | 0.08 |
| Furan, 2-pentyl- | Fruity, Green | - | 0.02 | 0.07 |
| Benzenamine | - | 1.69 | 0.78 | 0.99 |
| Total Other Compounds | 5.25 | 4.25 | 4.18 | |
| 10.63 | 81.32 | 104.45 | ||
| FM 1 | Benchmark 2 | UF 3 | |
|---|---|---|---|
| Bioactive peptides (mg/g) | 5.41 ± 0.17 b | 3.88 ± 0.34 a | 3.55 ± 0.28 a |
| Total SCFA (mg/kg) | 12.64 ± 0.89 c | 2.21 ± 0.50 b | 0.23 ± 0.09 a |
| Total phenols (Gallic acid mg eq/kg DM) | 451 ± 22 b | 295 ± 16 a | 263 ± 16 a |
| ABTS (TROLOX mg/kg DM) | 432 ± 24 b | 355 ± 16 a | 349 ± 26 a |
| DPPH (TROLOX mg/kg DM) | 229 ± 14 b | 199 ± 9 a | 196 ± 15 a |
| Phytic acid (g/100 g DM) | 0.76 ± 0.06 b | 0.95 ± 0.04 a | 1.05 ± 0.05 a |
| Bacterial Strains | UF 1 | FM 2 | FOS | INULIN |
|---|---|---|---|---|
| L. plantarum ATCC 8014 | 2.73 a | 2.78 a | 1.17 b | 0.95 c |
| L. rhamnosus ATCC 7469 | 2.80 a | 2.75 a | 1.03 b | 0.28 c |
| L. acidophilus ATCC 4356 | 2.08 b | 2.65 a | 0.97 c | 0.23 d |
| L. fermentum ATCC 9338 | 5.60 b | 7.86 a | 1.95 c | 1.13 c |
| B. angulatum DSM 20098 | 1.61 b | 2.66 a | 0.46 c | 2.16 ab |
| B. breve DSM 20091 | 1.96 b | 0.82 c | 1.59 bc | 5.70 a |
| B. longum DSM 20219 | 3.10 a | 2.15 b | 0.79 d | 1.27 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siroli, L.; Giordani, B.; Rossi, S.; Gottardi, D.; McMahon, H.; Augustyniak, A.; Menon, A.; Vannini, L.; Vitali, B.; Patrignani, F.; et al. Antioxidant and Functional Features of Pre-Fermented Ingredients Obtained by the Fermentation of Milling By-Products. Fermentation 2022, 8, 722. https://doi.org/10.3390/fermentation8120722
Siroli L, Giordani B, Rossi S, Gottardi D, McMahon H, Augustyniak A, Menon A, Vannini L, Vitali B, Patrignani F, et al. Antioxidant and Functional Features of Pre-Fermented Ingredients Obtained by the Fermentation of Milling By-Products. Fermentation. 2022; 8(12):722. https://doi.org/10.3390/fermentation8120722
Chicago/Turabian StyleSiroli, Lorenzo, Barbara Giordani, Samantha Rossi, Davide Gottardi, Helena McMahon, Aleksandra Augustyniak, Abhay Menon, Lucia Vannini, Beatrice Vitali, Francesca Patrignani, and et al. 2022. "Antioxidant and Functional Features of Pre-Fermented Ingredients Obtained by the Fermentation of Milling By-Products" Fermentation 8, no. 12: 722. https://doi.org/10.3390/fermentation8120722