
Pentostatin Biosynthesis Pathway Elucidation and Its Application


Abstract
:1. Introduction
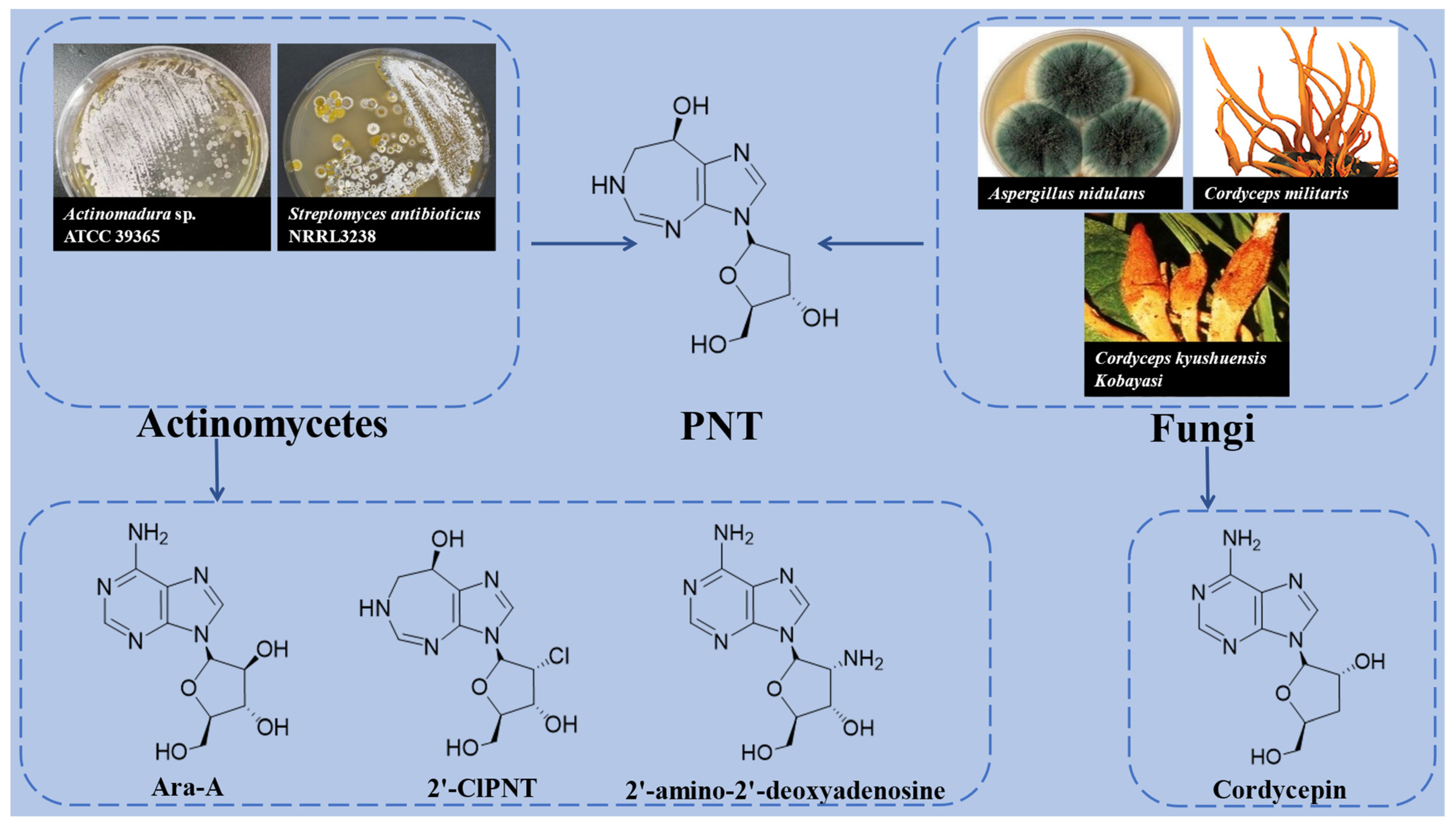
2. Resource Microorganisms of PNT
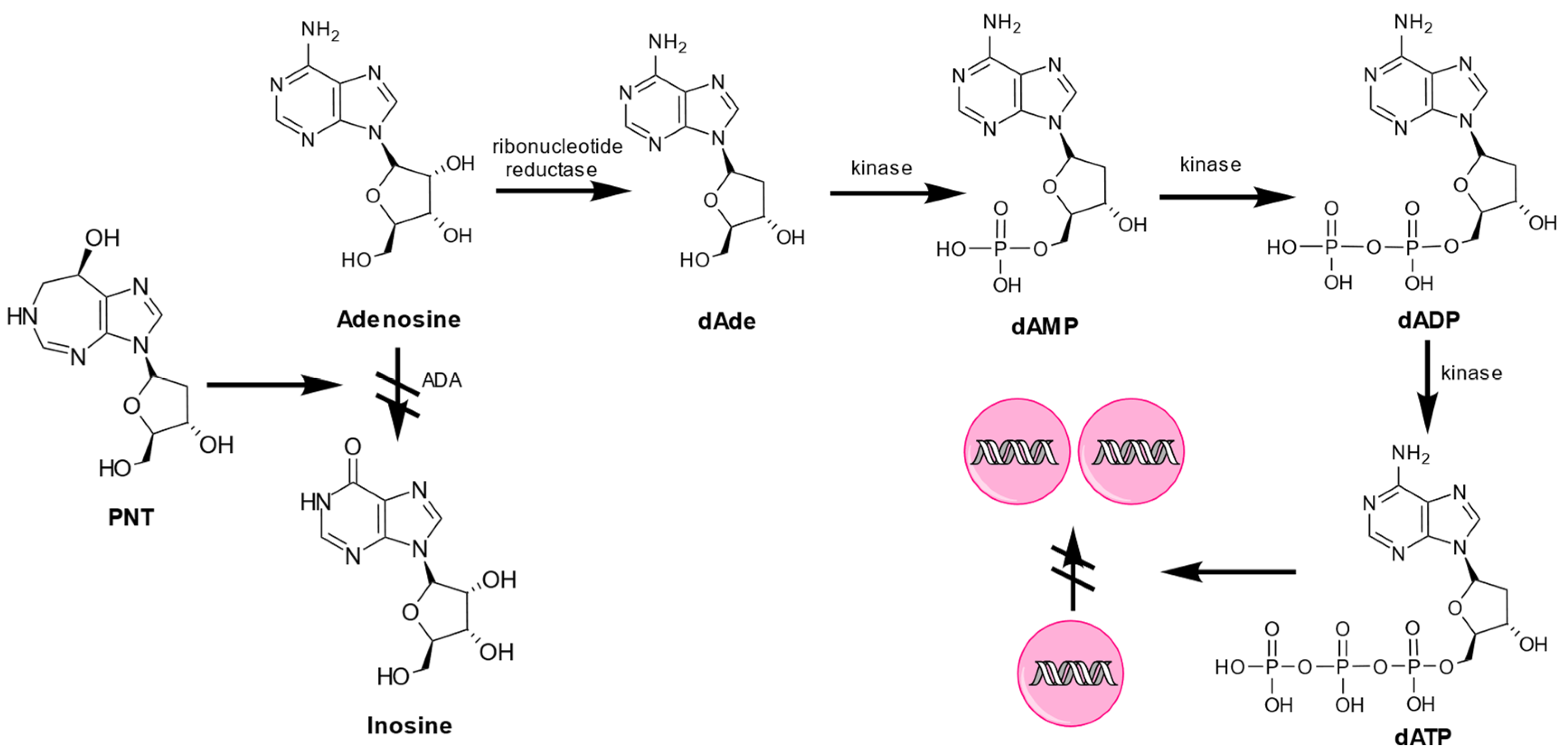
3. Action Mechanism of PNT
4. Biosynthesis Pathways of PNT
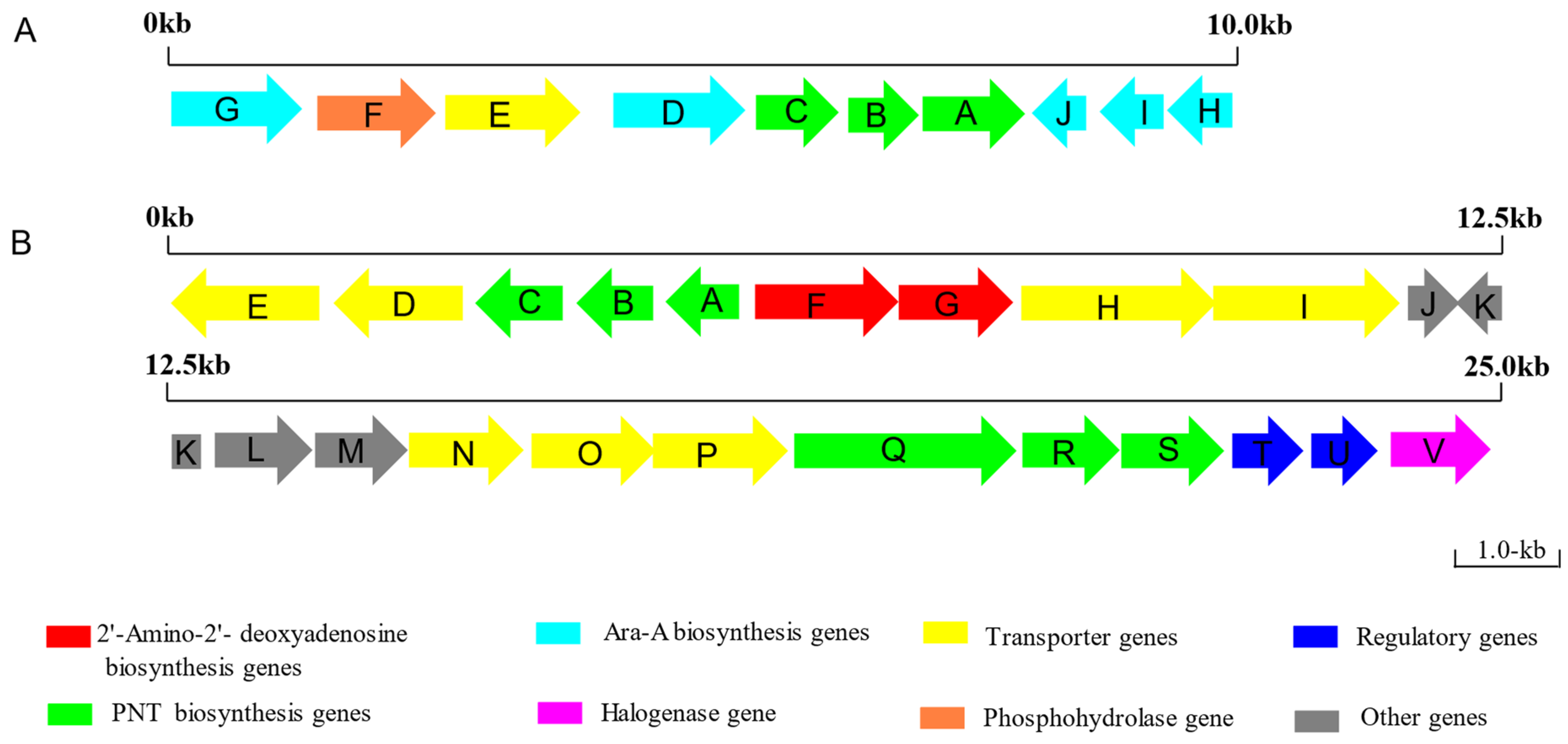
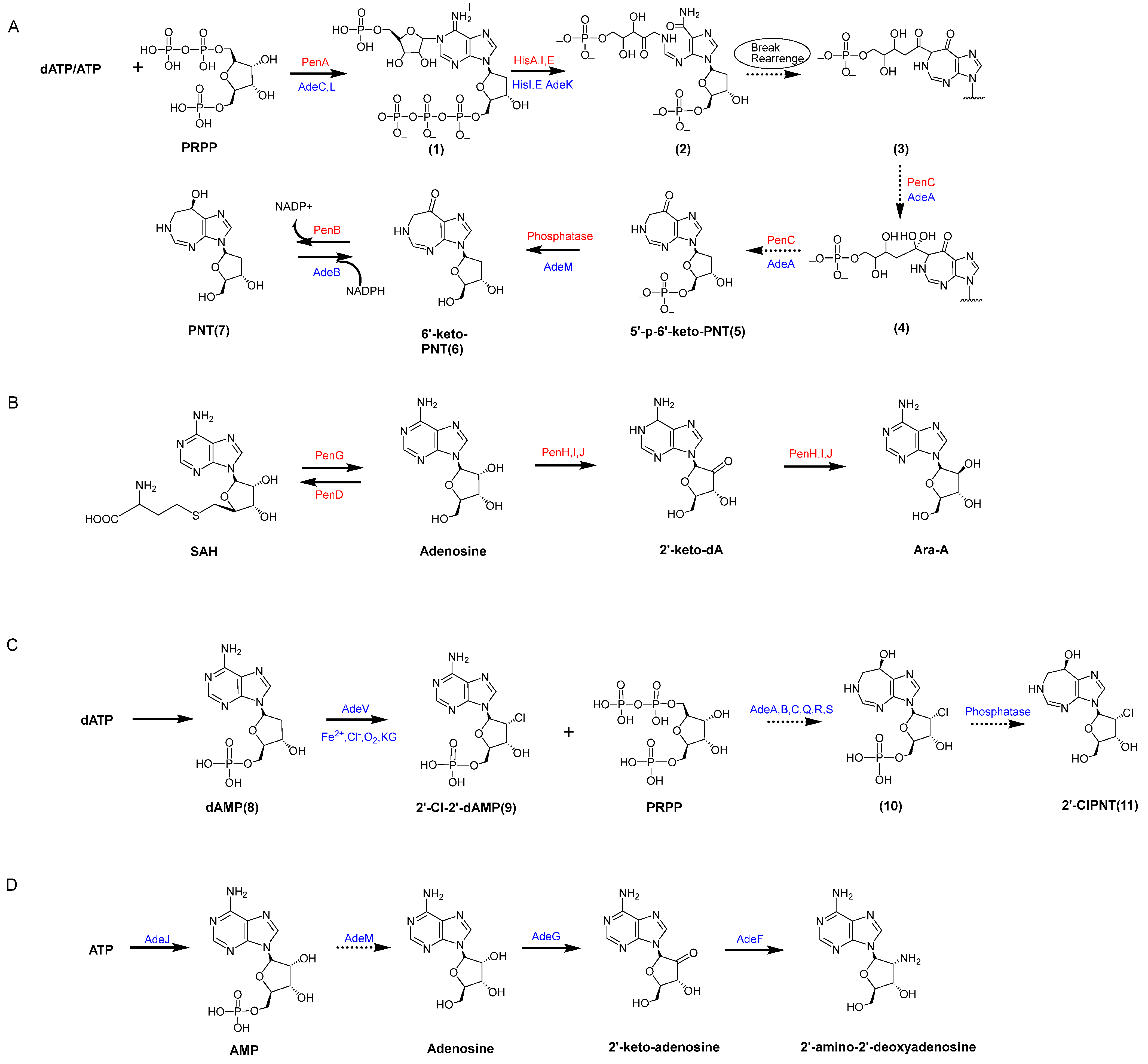
4.1. The PNT Biosynthesis Pathway in Streptomyces antibioticus NRRL 3238
4.2. The PNT Biosynthesis Pathway in Actinomadura sp. ATCC39365
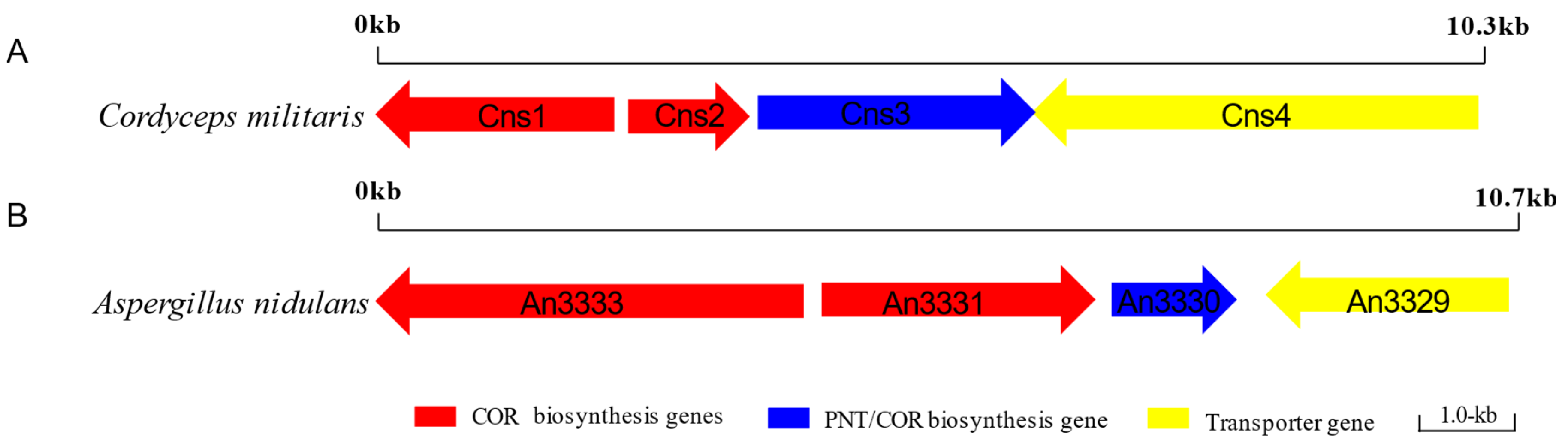
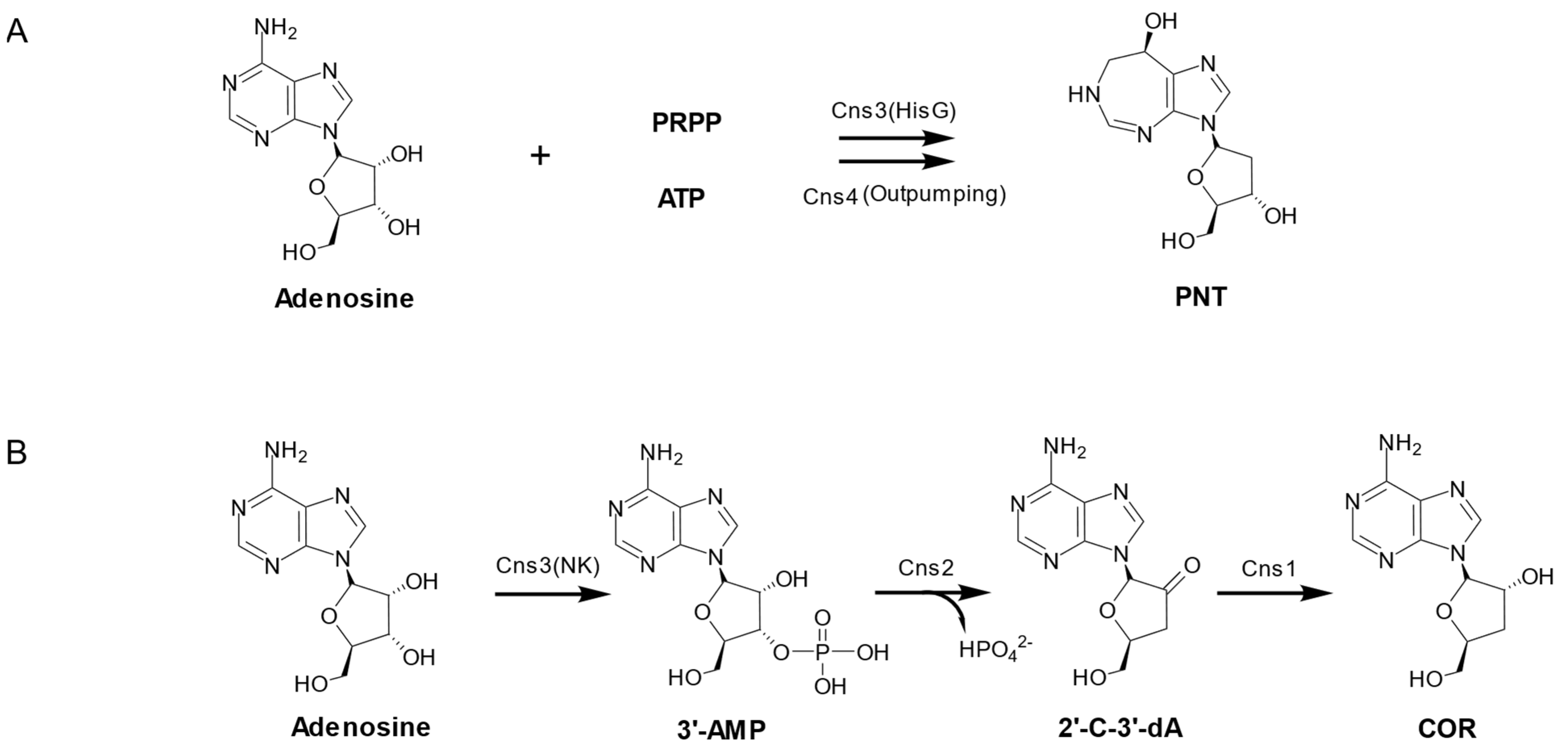
4.3. The PNT Biosynthesis Pathway in Cordyceps Militaris
5. Applications of PNT
5.1. Treatment of the Hairy Cell Leukemia
5.2. Treatment of the Chronic Lymphoblastic Leukemia
5.3. Treatment of Waldenstrom’s Macroglobulinemia
5.4. Inhibition of the Trypanosoma
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.A.; Wright, G.D. The past, present, and future of antibiotics. Sci. Transl. Med. 2022, 14, eabo7793. [Google Scholar] [CrossRef]
- Butler, M.S.; Buss, A.D. Natural products--the future scaffolds for novel antibiotics? Biochem. Pharmacol. 2006, 71, 919–929. [Google Scholar] [CrossRef]
- Shiraishi, T.; Kuzuyama, T. Recent advances in the biosynthesis of nucleoside antibiotics. J. Antibiot. 2019, 72, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Tan, H. Nucleoside antibiotics: Biosynthesis, regulation, and biotechnology. Trends Microbiol. 2015, 23, 110–119. [Google Scholar] [CrossRef]
- Osada, H. Discovery and applications of nucleoside antibiotics beyond polyoxin. J. Antibiot. 2019, 72, 855–864. [Google Scholar] [CrossRef]
- Woo, P.W.K.; Dion, H.W.; Lange, S.M.; Dahl, L.F.; Durham, L.J. A novel adenosine and Ara—A deaminase inhibitor, (R)-3-(2-deoxy-β-D-erythro-pentofuranosyl)-3,6,7,8- tetrahydroimidazo[4,5-d][1,3]diazepin-8-ol. J. Heterocycl. Chem. 1974, 11, 3. [Google Scholar] [CrossRef]
- Albert, R.; Henry, W.D.; Peter, W.W.; John, D.H. (R)-3-(2-deoxy-β-D-erythro-pentofuranosyl)-3,6,7,8-tetrahydroimidazo [4,5-d] [1,3]diazepin-8-ol. U.S. Patent 3,923,785, 22 April 1974. [Google Scholar]
- Chan, E.; Putt, S.R.; Showalter, H.H.; Baker, D.C. Total Synthesis of (8R)-3-(2-Deoxy-β-D-erythro -pentofuranosyl)-3,6,7,8-tetrahydroimidazo[4,5-d][1,3]diazepin-8-ol (Pentostatin), the Potent Inhibitor of Adenosine Deaminase. Am. Chem. Soc. 1982, 47, 3457–3464. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, G.Y.; Li, C.; Ling, J. Cordycepin and pentostatin biosynthesis gene identified through transcriptome and proteomics analysis of Cordyceps kyushuensis Kob. Microbiol. Res. 2019, 218, 12–21. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wan, D.; Xu, G.; Wang, G.; Ma, H.; Wang, T.; Gao, Y.; Qi, J.; Chen, X.; Zhu, J.; et al. An Unusual Protector-Protege Strategy for the Biosynthesis of Purine Nucleoside Antibiotics. Cell Chem. Biol. 2017, 24, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xu, G.; Wu, P.; Liu, J.; Cai, Y.S.; Deng, Z.; Chen, W. Biosynthesis of 2’-Chloropentostatin and 2’-Amino-2’-Deoxyadenosine Highlights a Single Gene Cluster Responsible for Two Independent Pathways in Actinomadura sp. Strain ATCC 39365. Appl. Environ. Microbiol. 2017, 83, e00078–e00087. [Google Scholar] [CrossRef]
- Kenjiro, K.; Kusakabe, H.; Machida, H.; Midorikawa, Y.; Shibuya, S.; Kuninaka, A.; Yoshino, H. Isolation of 2′-Deoxycoformycin and Cordycepin from Wheat Bran Culture of Aspergillus nidulans Y 176–2. Agric. Biol. Chem. 1979, 43, 2375–2377. [Google Scholar]
- Hanvey, J.C.; Hawkins, E.S.; Baker, D.C.; Suhadolnik, R.J. 8-Ketodeoxycoformycin and 8-Ketocoformycin as Intermediates in the Biosynthesis of 2’-Deoxycoformycin and Coformycin. Biochemistry 1988, 27, 5795–5800. [Google Scholar] [CrossRef]
- Isono, K. Nucleoside antibiotics: Structure, biological activity, and biosynthesis. J. Antibiot 1988, 41, 1711–1739. [Google Scholar] [CrossRef]
- Ho, A.D.; Hensel, M. Pentostatin for the treatment of indolent lymphoproliferative disorders. Semin. Hematol. 2006, 43 (Suppl. 2), S2–S10. [Google Scholar] [CrossRef]
- Iannitto, E.; Tripodo, C. How I diagnose and treat splenic lymphomas. Blood 2011, 117, 2585–2595. [Google Scholar] [CrossRef]
- Herth, I.; Hensel, M.; Rieger, M.; Horstmann, K.; Hiddemann, W.; Dreyling, M.; Koniczek, S.; Witzens-Harig, M.; Ho, A.D. Pentostatin, cyclophosphamide and rituximab is a safe and effective treatment in patients with Waldenstrom’s macroglobulinemia. Leuk. Lymphoma 2015, 56, 97–102. [Google Scholar] [CrossRef]
- do Carmo, G.M.; de Sa, M.F.; Grando, T.H.; Gressler, L.T.; Baldissera, M.D.; Monteiro, S.G.; Henker, L.C.; Mendes, R.E.; Stefani, L.M.; Da Silva, A.S. Cordycepin (3’-deoxyadenosine) and pentostatin (deoxycoformycin) against Trypanosoma cruzi. Exp. Parasitol. 2019, 199, 47–51. [Google Scholar] [CrossRef]
- Kanbe, T.; Endo, A.; Hashizume, T. Production of 2’-deoxycoformycin by the fungus Emericella nidulans and its inhibitory effect on adenosine deaminase. Nucleic Acids Symp. Ser. 1983, 12, 115–118. [Google Scholar]
- Alfayez, M.; Thakral, B.; Jain, P.; Ravandi, F.; Ferrajoli, A.; Jain, N.; Pemmaraju, N.; Wierda, W.; Kadia, T. First report of clinical response to venetoclax combination with pentostatin in T-cell-prolymphocytic leukemia (T-PLL). Leuk. Lymphoma 2020, 61, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Janus, A.; Robak, T. Moxetumomab pasudotox for the treatment of hairy cell leukemia. Expert Opin. Biol. Ther. 2019, 19, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Kempin, S.; Sun, Z.; Kay, N.E.; Paietta, E.M.; Mazza, J.J.; Ketterling, R.P.; Frankfurt, O.; Claxton, D.F.; Saltzman, J.N.; Srkalovic, G.; et al. Pentostatin, Cyclophosphamide, and Rituximab Followed by Alemtuzumab for Relapsed or Refractory Chronic Lymphocytic Leukemia: A Phase 2 Trial of the ECOG-Acrin Cancer Research Group (E2903). Acta Haematol. 2019, 142, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B. Mechanism of action of pentostatin and cladribine in Hairy Cell Leukemia. Leuk. Lymphoma 2011, 52 (Suppl. 2), 43–45. [Google Scholar] [CrossRef]
- Brogden, R.N.; Sorkin, E.M. Pentostatin A Review of its Pharmacodynamic and Pharmacokinetic Properties, and Therapeutic Potential in Lymphoproliferative Disorders. Drug Eval. 1993, 46, 652–677. [Google Scholar] [CrossRef]
- Malki, Y.; Martinez, J.; Masurier, N. 1,3-Diazepine: A privileged scaffold in medicinal chemistry. Med. Res. Rev. 2021, 41, 2247–2315. [Google Scholar] [CrossRef]
- Hanvey, J.C.; Hardman, J.K.; Suhadolnik, R.J.; Baker, D.C. Evidence for the Conversion of Adenosine to 2’-Deoxycoformycin by Streptomyces antibioticus. Biochemistry 1984, 23, 904–907. [Google Scholar] [CrossRef]
- Hanvey, J.C.; Hawkins, E.S.; Tunac, J.B.; Dechter, J.J.; Baker, D.C.; Suhadolnik, R.J. Biosynthesis of 2’-Deoxycoformycin: Evidence for Ring Expansion of the Adenine Moiety of Adenosine to a Tetrahydroimidazo[ 4,5-d][1,3]diazepine System. Biochemistry 1986, 26, 6. [Google Scholar] [CrossRef]
- Bauman, K.D.; Butler, K.S.; Moore, B.S.; Chekan, J.R. Genome mining methods to discover bioactive natural products. Nat. Prod. Rep. 2021, 38, 2100–2129. [Google Scholar] [CrossRef]
- Xu, Y.; Du, X.; Yu, X.; Jiang, Q.; Zheng, K.; Xu, J.; Wang, P. Recent Advances in the Heterologous Expression of Biosynthetic Gene Clusters for Marine Natural Products. Mar. Drugs 2022, 20, 341. [Google Scholar] [CrossRef] [PubMed]
- Pornbanlualap, S.; Chalopagorn, P. Adenosine deaminase from Streptomyces coelicolor: Recombinant expression, purification and characterization. Protein Expr. Purif. 2011, 78, 167–173. [Google Scholar] [CrossRef]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Ruszczycky, M.W.; Ko, Y.; Wang, S.-A.; Ogasawara, Y.; Kim, M.; Liu, H.-W. Characterization of the coformycin biosynthetic gene cluster in Streptomyces kaniharaensis. Proc. Natl. Acad. Sci. USA 2020, 117, 10265–10270. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.C.; Kuppan, G.; Shetty, N.D.; Owen, J.L.; Ioerger, T.R.; Sacchettini, J.C. Crystal structures of Mycobacterium tuberculosis S-adenosyl-L-homocysteine hydrolase in ternary complex with substrate and inhibitors. Protein Sci. 2008, 17, 2134–2144. [Google Scholar] [CrossRef]
- Zhao, C.H.; Yan, S.; Li, Q.; Zhu, H.C.; Zhong, Z.Y.; Ye, Y.; Deng, Z.X.; Zhang, Y.H. An Fe2+- and alpha-Ketoglutarate-Dependent Halogenase Acts on Nucleotide Substrates. Angew. Chem. Int. Edit. 2020, 59, 9478–9484. [Google Scholar] [CrossRef]
- Matsuyama, K.; Takahashi, Y.; Yamashita, M.; Hirano, A.; Omura, S. 2’-Amino-2’-deoxyadenosine produced by a strain of Actinomadura. J. Antibiot. 1979, 32, 1367–1369. [Google Scholar] [CrossRef]
- Suhadolnik, R.J.; Pornbanlualap, S.; Baker, D.C.; Tiwari, K.N.; Hebbler, A.K. Stereospecific 2’-amination and 2’-chlorination of adenosine by Actinomadura in the biosynthesis of 2’-amino-2’-deoxyadenosine and 2’-chloro-2’-deoxycoformycin. Arch. Biochem. Biophys. 1989, 270, 374–382. [Google Scholar] [CrossRef]
- Goto, Y.; Li, B.; Claesen, J.; Shi, Y.; Bibb, M.J.; van der Donk, W.A. Discovery of unique lanthionine synthetases reveals new mechanistic and evolutionary insights. PLoS Biol. 2010, 8, e1000339. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Y.; Xiao, G.; Zheng, P.; Xia, Y.; Zhang, X.; Leger, R.J.S.; Liu, X.; Wang, C. Genome survey uncovers the secrets of sex and lifestyle in caterpillar fungus. Chin. Sci. Bull. 2013, 58, 2846–2854. [Google Scholar] [CrossRef]
- Goldstone, D.C.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.T.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W.; et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Saven, A.; Burian, C.; Koziol, J.A.; Piro, L.D. Long-Term Follow-Up of Patients With Hairy Cell Leukemia After Cladribine Treatment. Blood 1998, 92, 1918–1926. [Google Scholar] [CrossRef] [PubMed]
- Paillassa, J.; Troussard, X. Patients with relapsed/refractory hairy-cell leukemia. Cancer Rep. 2022, 5, e1495. [Google Scholar] [CrossRef] [PubMed]
- Kreitman, R.J. Hairy cell leukemia: Present and future directions. Leuk Lymphoma 2019, 60, 2869–2879. [Google Scholar] [CrossRef] [PubMed]
- Paillassa, J.; Maitre, E.; Troussard, X. Hairy Cell Leukemia (HCL) and HCL Variant: Updates and Spotlights on Therapeutic Advances. Curr. Oncol. Rep. 2022, 13, 974–982. [Google Scholar] [CrossRef]
- Grever, M.; Kopecky, K.; Foucar, M.K.; Head, D.; Bennett, J.M.; Hutchison, R.E.; Corbett, W.E.; Cassileth, P.A.; Habermann, T.; Golomb, H. Randomized Comparison of Pentostatin Versus Interferon Alfa-2a in Previously Untreated Patients With Hairy Cell Leukemia: An Intergroup Study. J. Clin. Oncol. 1995, 13, 974–982. [Google Scholar] [CrossRef]
- Bohn, J.-P.; Gastl, G.; Steurer, M. Long-term treatment of hairy cell leukemia with interferon-alpha: Still a viable therapeutic option. Memo 2016, 9, 63–65. [Google Scholar] [CrossRef]
- Kost, S.E.; Bouchard, E.D.; Labossière, E.; Ye, X.; Queau, M.L.; Liang, W.S.; Banerji, V.; Gibson, S.B.; Katyal, S.; Johnston, J.B. Cross-resistance and synergy with bendamustine in chronic lymphocytic leukemia. Leuk. Res. 2016, 50, 63–71. [Google Scholar] [CrossRef]
- Else, M.; Dearden, C.E.; Matutes, E.; Garcia-Talavera, J.; Rohatiner, A.Z.S.; Johnson, S.A.N.; O’Connor, N.T.J.; Haynes, A.; Osuji, N.; Forconi, F.; et al. Long-term follow-up of 233 patients with hairy cell leukaemia, treated initially with pentostatin or cladribine, at a median of 16 years from diagnosis. Br. J. Haematol. 2009, 145, 733–740. [Google Scholar] [CrossRef]
- Salam, L.; Abdel-Wahab, O. Hairy cell leukemia: Update and current therapeutic approach. Curr. Opin. Hematol. 2015, 22, 355–361. [Google Scholar] [CrossRef]
- Grever, M.R. Hairy cell leukemia: A successful model for experimental therapeutics-Pentostatin and new ideas. Leuk. Lymphoma 2011, 52 (Suppl. 2), 25–28. [Google Scholar] [CrossRef]
- Tedeschi, A.; Rossi, D.; Motta, M.; Quaresmini, G.; Rossi, M.; Coscia, M.; Anastasia, A.; Rossini, F.; Cortelezzi, A.; Nador, G.; et al. A phase II multi-center trial of pentostatin plus cyclophosphamide with ofatumumab in older previously untreated chronic lymphocytic leukemia patients. Haematologica 2015, 100, E501–E504. [Google Scholar] [CrossRef] [Green Version]
- Dillman, R.O. Pentostatin (Nipent) in the treatment of chronic lymphocyte leukemia and hairy cell leukemia. Expert Rev. Anticancer Ther. 2004, 4, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Keating, M.J.; O’brien, S.; Koller, C.; Beran, M.; Robertson, L.E.; Freireich, E.J.; Estey, E.; Kantarjian, H. Long-term follow-up of patients with chronic lymphocytic leukemia (CLL) receiving fludarabine regimens as initial therapy. Blood 1998, 92, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Samaniego, F.; Hagemeister, F.; Romaguera, J.E.; Fanale, M.A.; Pro, B.; McLaughlin, P.; Rodriguez, M.A.; Neelapu, S.S.; Fayad, L.; Younes, A.; et al. Pentostatin, cyclophosphamide and rituximab for previously untreated advanced stage, low-grade B-cell lymphomas. Br. J. Haematol. 2015, 169, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Leleu, X.; Soumerai, J.; Roccaro, A.; Hatjiharissi, E.; Hunter, Z.R.; Manning, R.; Ciccarelli, B.T.; Sacco, A.; Ioakimidis, L.; Adamia, S.; et al. Increased incidence of transformation and myelodysplasia/acute leukemia in patients with Waldenstrom macroglobulinemia treated with nucleoside analogs. J. Clin. Oncol 2009, 27, 250–255. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Anagnostopoulos, A. Waldenstrom’s macroglobulinemia. Best Pract. Res. Clin. Haematol. 2005, 18, 747–765. [Google Scholar] [CrossRef]
- Hensel, M.; Villalobos, M.; Kornacker, M.; Krasniqi, F.; Ho, A.D. Pentostatin/cyclophosphamide with or without rituximab: An effective regimen for patients with Waldenstrom’s macroglobulinemia/lymphoplasmacytic lymphoma. Clin. Lymphoma Myeloma 2005, 6, 131–135. [Google Scholar] [CrossRef]
- Bern, C. Chagas’ Disease. N. Engl. J. Med. 2015, 373, 456–466. [Google Scholar] [CrossRef]
- Rodrigues, A.; Fighera, R.A.; Souza, T.M.; Schild, A.L.; Soares, M.P.; Milano, J.; Barros, C.S. Outbreaks of trypanosomiasis in horses by Trypanosoma evansi in the state of Rio Grande do Sul, Brazil: Epidemiological, clinical, hematological, and pathological aspects. Pesqui. Vet. Bras. 2005, 25, 11. [Google Scholar]
- Herrera, H.M.; Davila, A.M.; Norek, A.; Abreu, U.G.; Souza, S.S.; D’Andrea, P.S.; Jansen, A.M. Enzootiology of Trypanosoma evansi in Pantanal, Brazil. Vet. Parasitol. 2004, 125, 263–275. [Google Scholar] [CrossRef]
- Rosa, L.D.; Da Silva, A.S.; Ruchel, J.B.; Gressler, L.T.; Oliveira, C.B.; Franca, R.T.; Lopes, S.T.; Leal, D.B.; Monteiro, S.G. Influence of treatment with 3’-deoxyadenosine associated deoxycoformycin on hematological parameters and activity of adenosine deaminase in infected mice with Trypanosoma evansi. Exp. Parasitol. 2013, 135, 357–362. [Google Scholar] [CrossRef]
- Rosa, L.D.; Da Silva, A.S.; Oliveira, C.B.; Gressler, L.T.; Arnold, C.B.; Baldissera, M.D.; Sagrillo, M.; Sangoi, M.; Moresco, R.; Mendes, R.E.; et al. Dose finding of 3′ deoxyadenosine and deoxycoformycin for the treatment of Trypanosoma evansi infection: An effective and nontoxic dose. Microb. Pathog. 2015, 85, 21–28. [Google Scholar] [CrossRef]
- Carmo, G.M.D.; Doleski, P.H.; de Sá, M.F.; Grando, T.H.; Azevedo, M.I.; Manzoni, A.G.; Leal, D.B.; Gressler, L.T.; Henker, L.C.; Mendes, R.E.; et al. Treatment with 3’-deoxyadenosine and deoxycoformycin in mice infected by Trypanosoma cruzi and its side effect on purinergic enzymes. Microb. Pathog. 2017, 113, 51–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Resource Microorganisms | Associated Products | References |
|---|---|---|---|
| Actinomycetes | Streptomyces antibioticus NRRL3238 | Ara-A | [13,21,22] |
| Actinomadura sp. ATCC 39365 | 2′-ClPNT, 2′-amino-2′-deoxyadenosine | [8,13] | |
| Fungi | Aspergillus nidulans | COR | [15] |
| Cordyceps militaris | COR | [12] | |
| Cordyceps kyushuensis Kobayasi | COR | [11] |
| Protein | Size (aa) | Proposed Function |
|---|---|---|
| PenG | 387 | SAH hydrolase |
| PenF | 358 | Phosphohydrolase |
| PenE | 401 | membrane transport protein |
| PenD | 396 | SAH hydrolase |
| PenC | 247 | SAICAR synthetase |
| PenB | 234 | Short-chain dehydrogenase |
| PenA | 302 | ATP phosphoribosyl-transferase |
| PenJ | 161 | Dehydrogenase |
| PenI | 189 | Dehydrogenase |
| PenH | 213 | Dehydrogenase |
| Protein | Size (aa) | Proposed Function |
|---|---|---|
| AdeE | 479 | Cation/H+ antiporter |
| AdeD | 402 | MFS transporter |
| AdeC | 295 | ATP phosphoribosyl-transferase |
| AdeB | 234 | Short-chain dehydrogenase |
| AdeA | 239 | SAICAR synthetase |
| AdeF | 425 | Aminotransferase |
| AdeG | 351 | Dehydrogenase |
| AdeH | 595 | ABC transporter, partial |
| AdeI | 592 | ABC transporter |
| AdeJ | 161 | Nudix hydrolase |
| AdeK | 257 | Phosphoribosyl isomerase A |
| AdeL | 288 | ATP phosphoribosyl-transferase |
| AdeM | 264 | Hydrolase |
| AdeN | 347 | ABC transporter substrate-binding protein |
| AdeO | 363 | Sugar ABC transporter permease |
| AdeP | 406 | Nucleoside ABC transporter |
| AdeQ | 700 | N, N-dimethylformamidase |
| AdeR | 303 | Glucosamine-6-phosphate deaminase |
| AdeS | 309 | Ribokinase |
| AdeT | 241 | Bacterial regulatory protein, gntR family |
| AdeU | 231 | Bacterial regulatory protein, luxR family |
| AdeV | 310 | 2OG-Fe (II) oxygenase |
| Protein | Size (aa) | Proposed Function |
|---|---|---|
| Cns1 | 792 | Oxidoreductase/dehydrogenase |
| Cns2 | 345 | Metal-dependent phosphohydrolase |
| Cns3 | 871 | Phosphoribosyltransferase |
| Cns4 | 1364 | ATP-binding cassette (ABC) transporter |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, R.; Lou, T.; Zhao, P.; Wang, S. Pentostatin Biosynthesis Pathway Elucidation and Its Application. Fermentation 2022, 8, 459. https://doi.org/10.3390/fermentation8090459
Zhang H, Liu R, Lou T, Zhao P, Wang S. Pentostatin Biosynthesis Pathway Elucidation and Its Application. Fermentation. 2022; 8(9):459. https://doi.org/10.3390/fermentation8090459
Chicago/Turabian StyleZhang, Hongyu, Ran Liu, Tingting Lou, Pei Zhao, and Suying Wang. 2022. "Pentostatin Biosynthesis Pathway Elucidation and Its Application" Fermentation 8, no. 9: 459. https://doi.org/10.3390/fermentation8090459