
Improved Tolerance of Lactiplantibacillus plantarum in the Presence of Acid by the Heterologous Expression of trxA from Oenococcus oeni
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Growth Conditions, and Plasmids
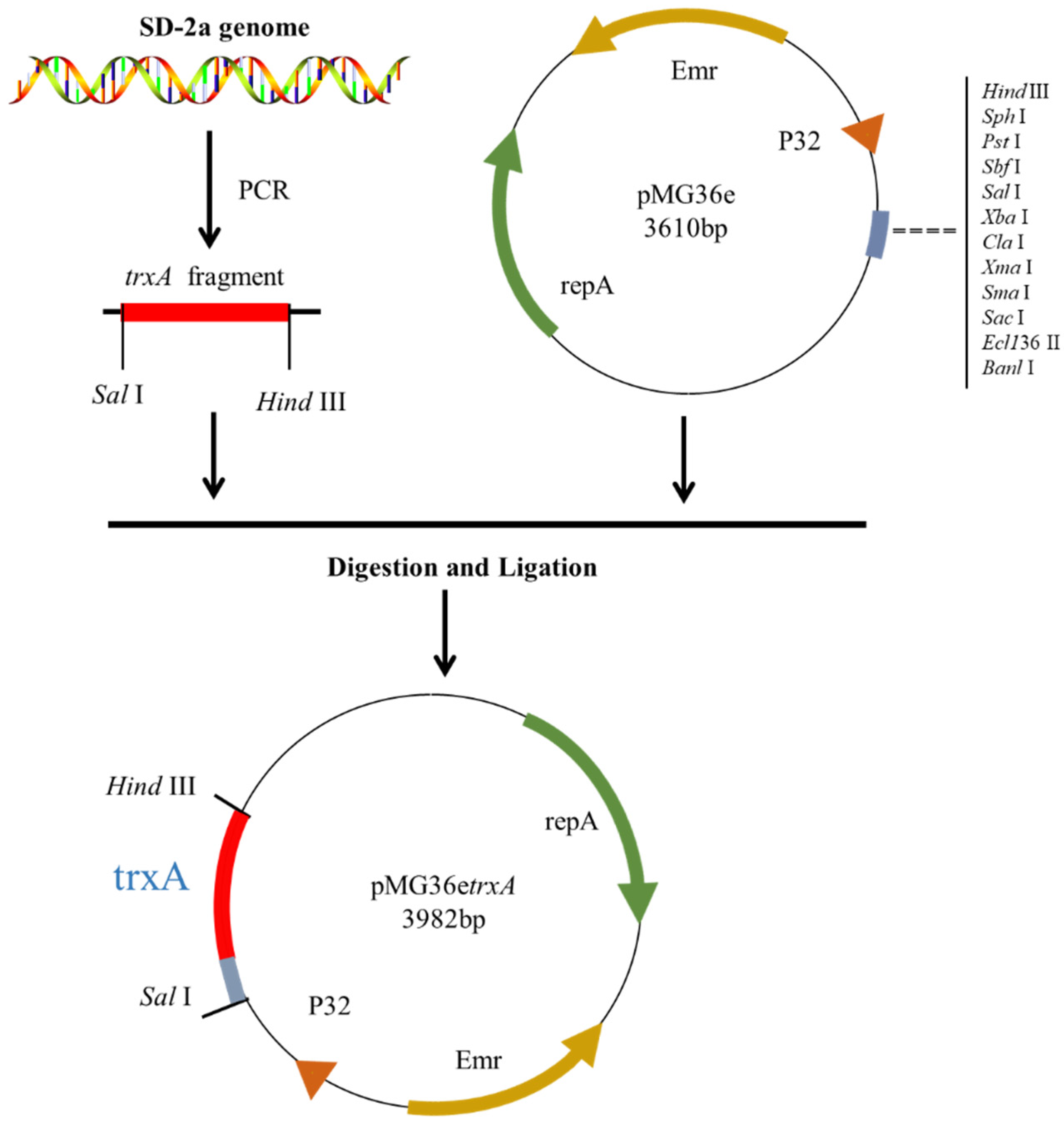
2.2. Plasmid Construction and Transformation
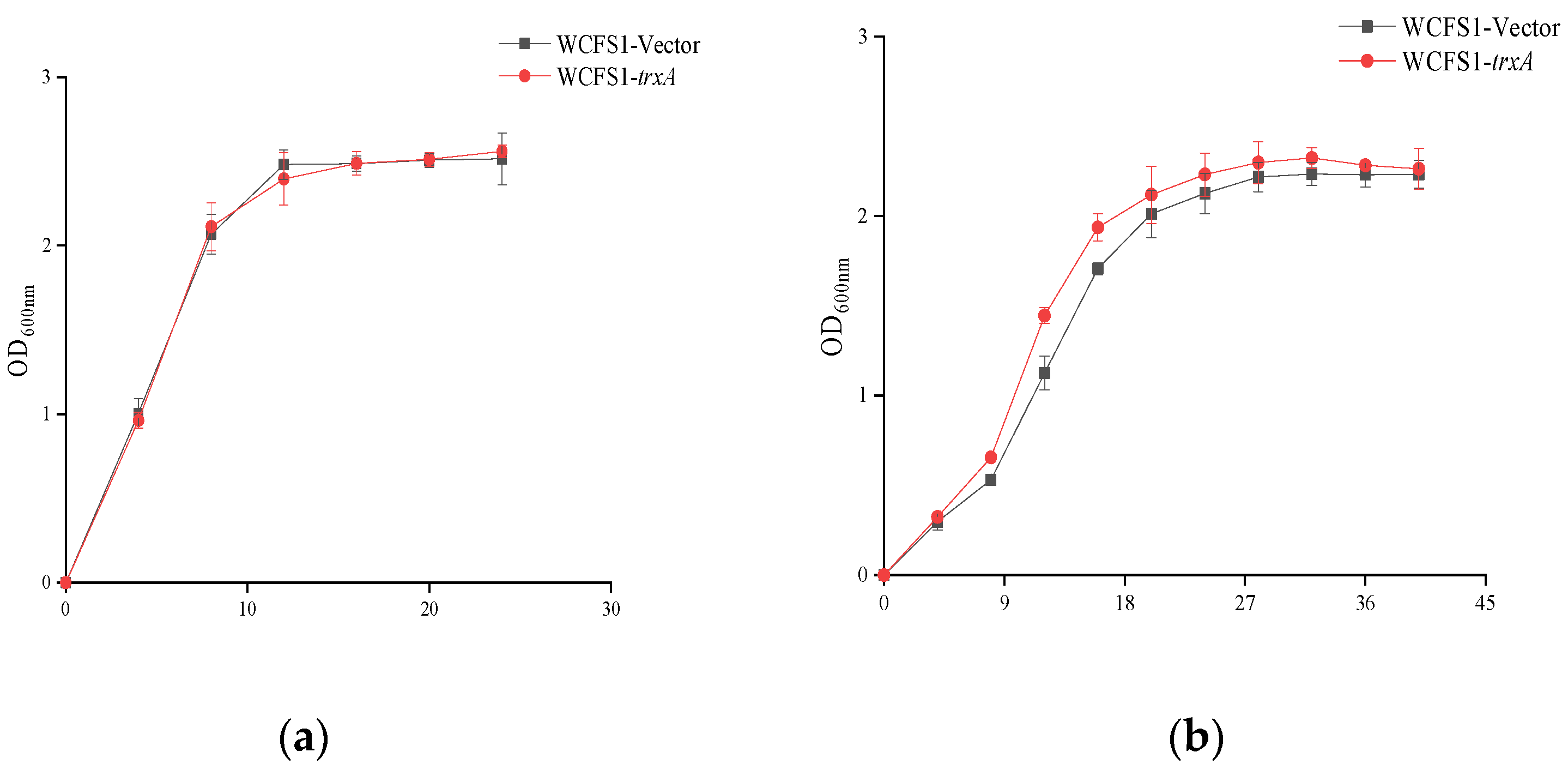
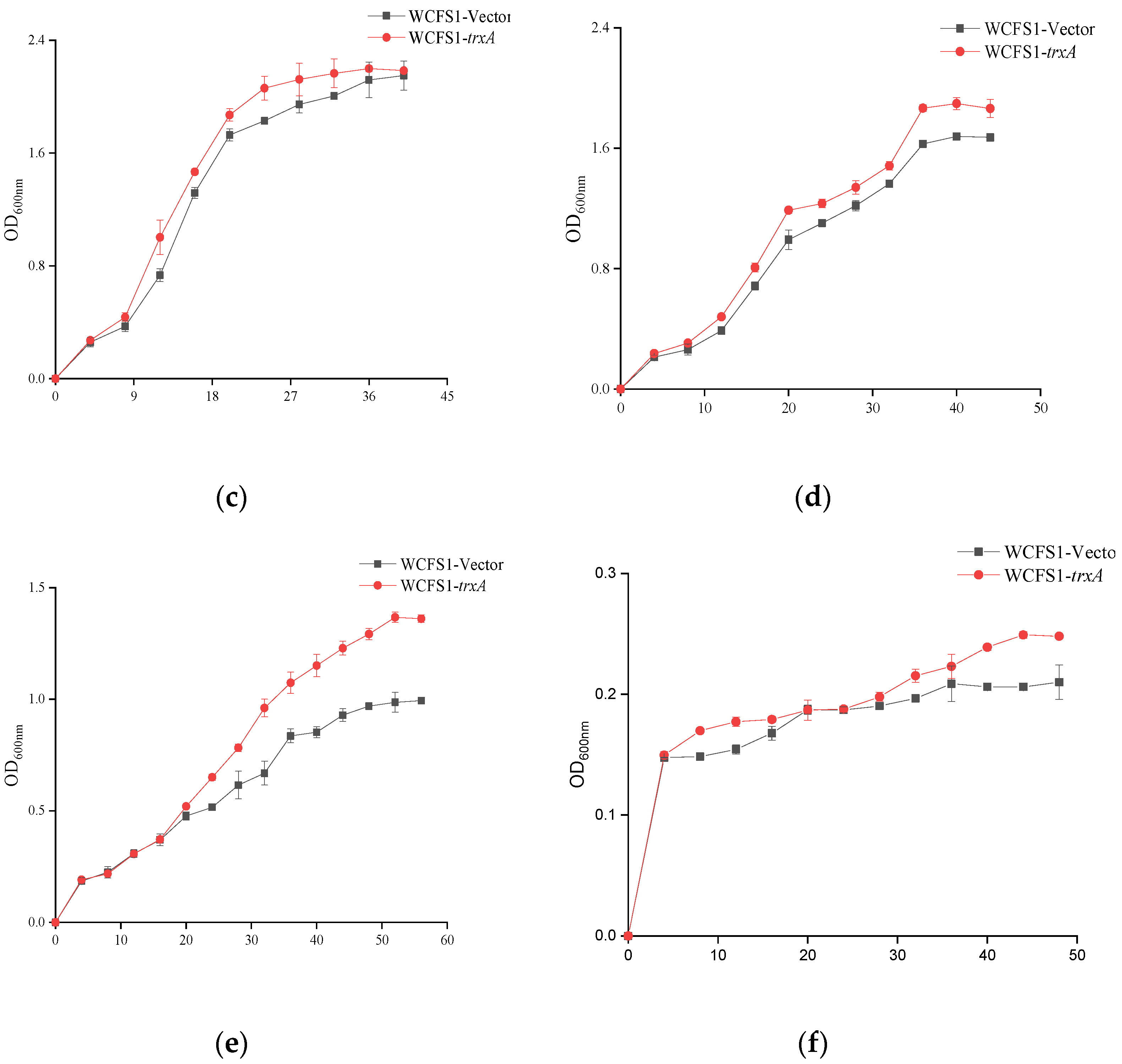
2.3. Stress Challenges and the Growth Performance
2.4. ROS Accumulation and Membrane Integrity Evaluation
2.5. Measurement of Cell Membrane Fluidity
2.6. Intracellular ATP Production, Superoxide Dismutase Activity, and Glutathione Concentration
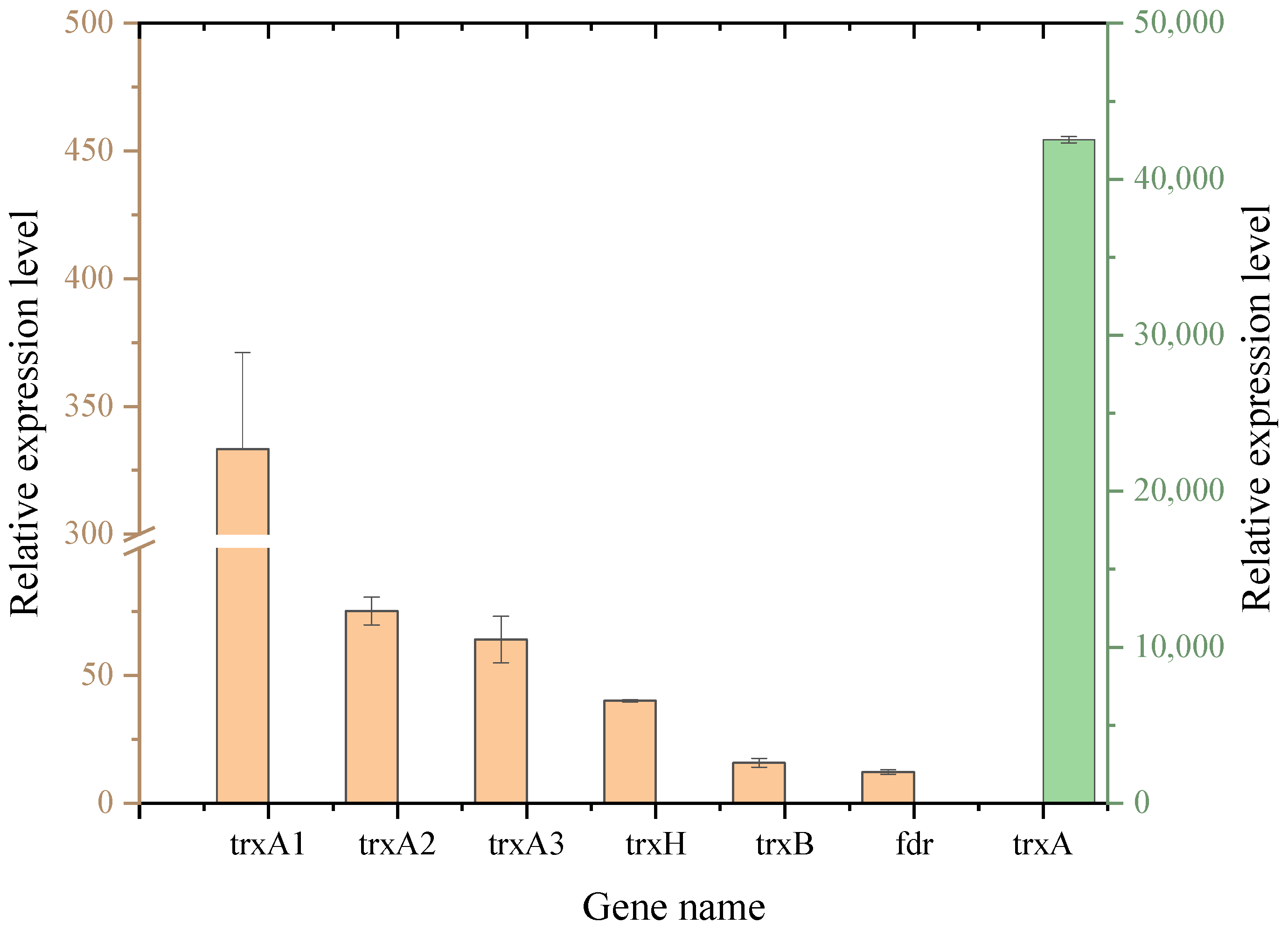
2.7. Real-Time Quantitative PCR
2.8. Statistical Analysis
3. Results and Discussion
3.1. Heterologous Expression of pMG36etrxA in L. plantarum WCFS1
3.2. Improved Growth Ability of WCFS1-pMG36etrxA
3.3. Heterologous Expression of trxA Affects the Transcription Levels of the Inherent Trx System Genes in L. plantarum WCFS1
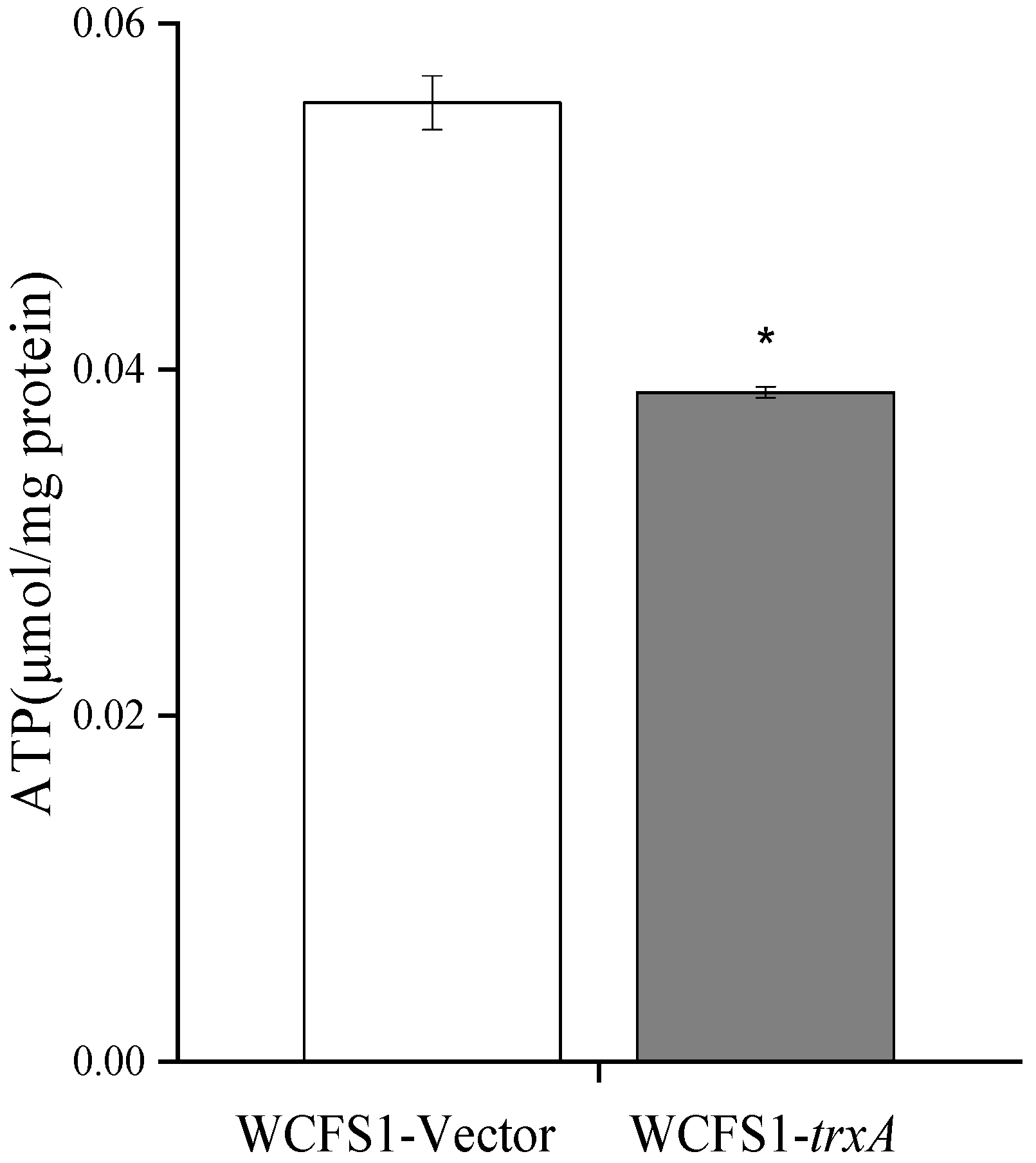
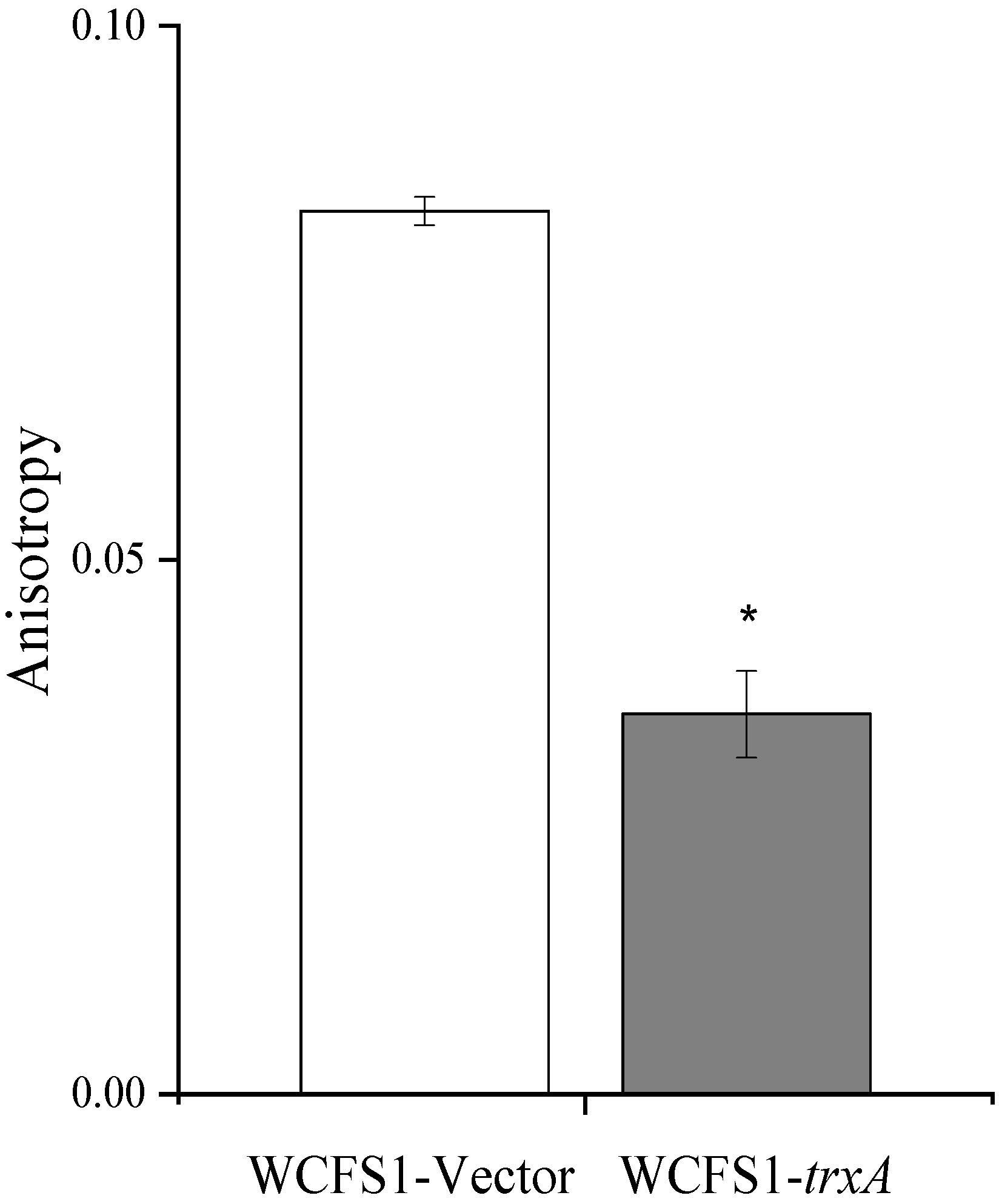
3.4. Heterologous Expression of trxA Decreased ATP Content and Increased Cell Membrane Fluidity in L. plantarum WCFS1
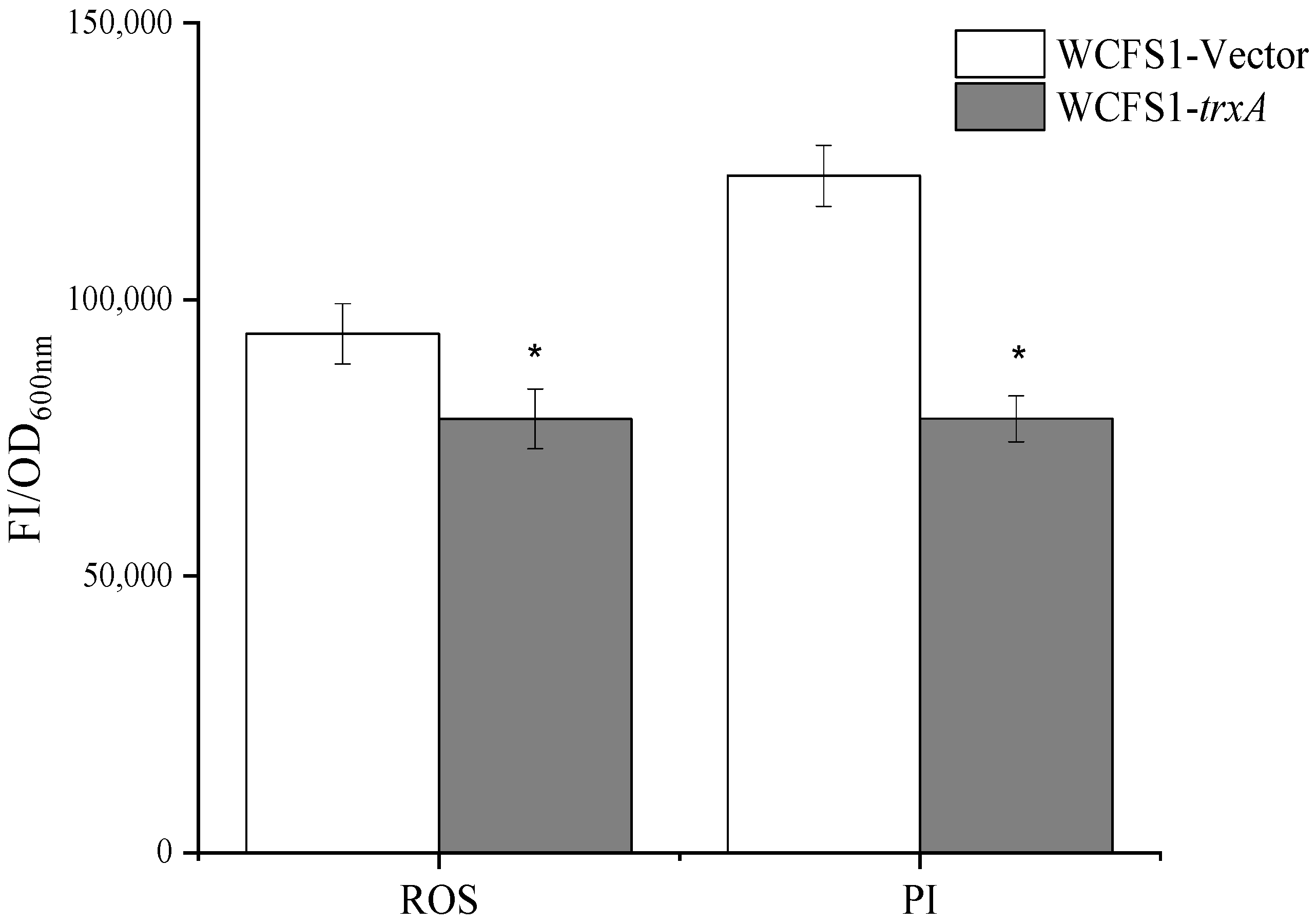
3.5. Heterologous Expression of trxA Results in Decreased ROS Accumulation and Enhanced Cell Membrane Integrity of L. plantarum WCFS1
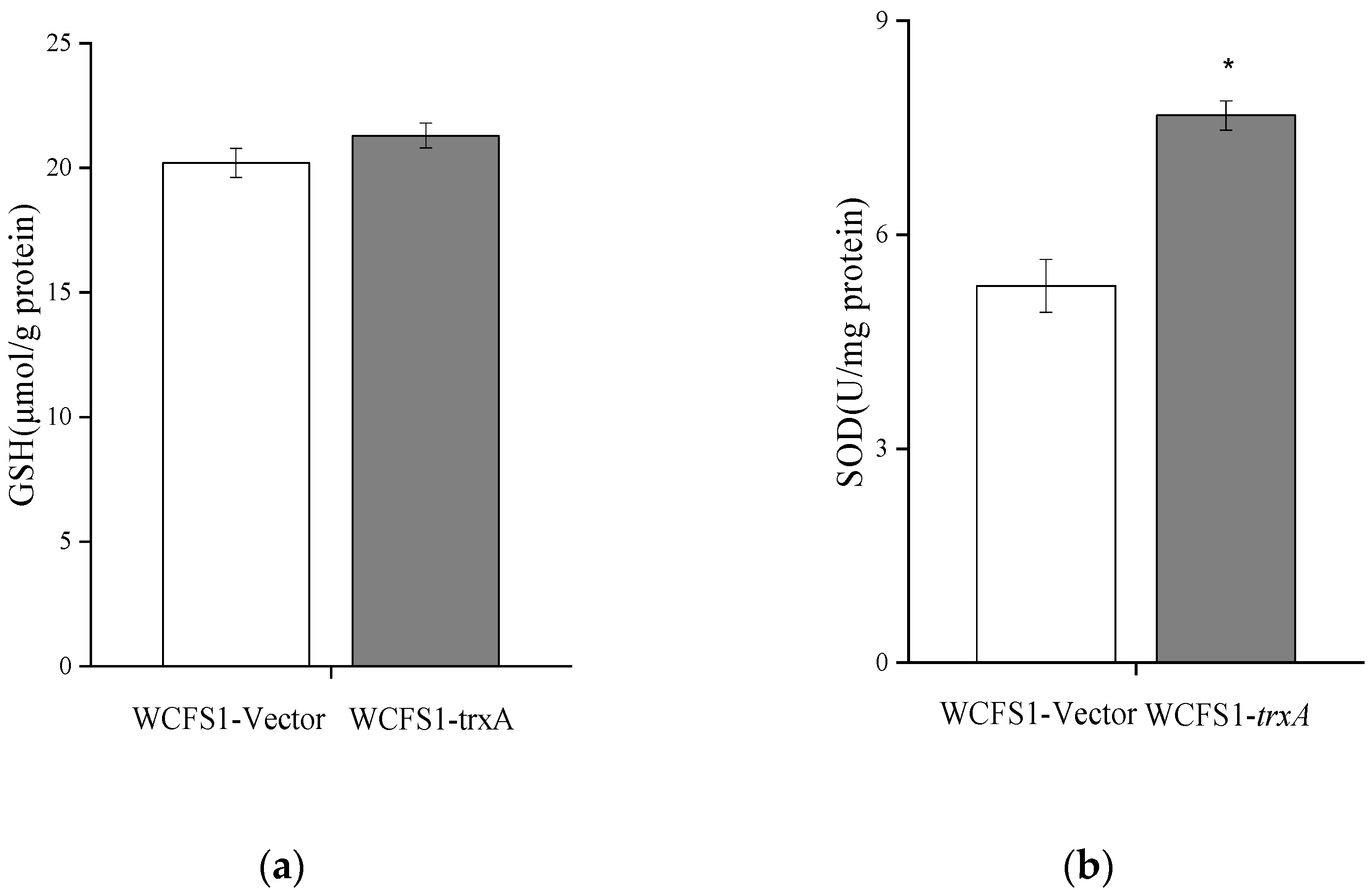
3.6. Heterologous Expression of trxA Decreased ROS Accumulation through Improving SOD Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, H.; Li, Y.; Liu, L.; Zheng, M.; Feng, Z.; Hu, K.; Tao, Y. Effects of inoculation timing and mixed fermentation with Pichia fermentans on Oenococcus oeni viability, fermentation duration and aroma production during wine malolactic fermentation. Food Res. Int. 2022, 159, 111604. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Peng, S.; Song, W.; Zhao, H.; Li, H.; Wang, H. Genomic analysis of an excellent wine-making strain Oenococcus oeni SD-2a. Pol. J. Microbiol. 2022, 71, 279–292. [Google Scholar] [CrossRef]
- Balmaseda, A.; Rozes, N.; Bordons, A.; Reguant, C. Molecular adaptation response of Oenococcus oeni in non-Saccharomyces fermented wines: A comparative multi-omics approach. Int. J. Food Microbiol. 2022, 362, 109490. [Google Scholar] [CrossRef] [PubMed]
- Diez-Ozaeta, I.; Lavilla, M.; Amarita, F. Effect of inoculation strategy with autochthonous Oenococcus oeni strains on aroma development in Rioja Alavesa Tempranillo wines. LWT–Food Sci. Technol. 2022, 162, 113399. [Google Scholar] [CrossRef]
- Noemi, B.; Giorgia, P.; Carlo, P.; Giuseppe, A.; Camillo, Z.; Alessio, P.R.; Rosanna, T. Characterization of natural Oenococcus oeni strains for Montepulciano d’Abruzzo organic wine production. Eur. Food Res. Technol. 2020, 246, 1031–1039. [Google Scholar]
- Maret, D.T.; Lynn, E.; Elda, L.; Sibylle, K. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioproc. Technol. 2011, 4, 876–906. [Google Scholar]
- Jin, G.; Jiranek, V.; Hayes, A.M.; Grbin, P.R. Isolation and characterization of high-ethanol-tolerance lactic acid bacteria from Australian Wine. Foods 2022, 11, 1231. [Google Scholar] [CrossRef] [PubMed]
- Olguin, N.; Champomier-Verges, M.; Anglade, P.; Baraige, F.; Cordero-Otero, R.; Bordons, A.; Zagorec, M.; Reguant, C. Transcriptomic and proteomic analysis of Oenococcus oeni PSU-1 response to ethanol shock. Food Microbiol. 2015, 51, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Darsonval, M.; Msadek, T.; Alexandre, H.; Grandvalet, C. The antisense RNA approach: A new application for in vivo investigation of the stress response of Oenococcus oeni, a wine-associated lactic acid bacterium. Appl. Environ. Microbiol. 2016, 82, 18–26. [Google Scholar] [CrossRef]
- Darsonval, M.; Julliat, F.; Msadek, T.; Alexandre, H.; Grandvalet, C. CtsR, the master regulator of stress-response in Oenococcus oeni, is a heat sensor interacting with ClpL1. Front. Microbiol. 2018, 9, 3135. [Google Scholar] [CrossRef]
- Darsonval, M.; Alexandre, H.; Grandvalet, C. Genetically engineered Oenococcus oeni strains to highlight the impact of estA2 and estA7 esterase genes on wine ester profile. Food Microbiol. 2016, 60, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liu, L.; Peng, S.; Yuan, L.; Li, H.; Wang, H. Heterologous expression of argininosuccinate synthase from Oenococcus oeni enhances the acid resistance of Lactobacillus plantarum. Front. Microbiol. 2019, 10, 1393. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhao, H.; Liu, L.; Peng, S.; Li, H.; Wang, H. Heterologous expression of the puuE from Oenococcus oeni SD-2a in Lactobacillus plantarum WCFS1 improves ethanol tolerance. J. Basic Microbiol. 2019, 59, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yuan, L.; Hu, K.; Liu, L.; Peng, S.; Li, H.; Wang, H. Heterologous expression of ctsR from Oenococcus oeni enhances the acid-ethanol resistance of Lactobacillus plantarum. FEMS Microbiol. Lett. 2019, 366, fnz192. [Google Scholar] [CrossRef]
- Schumann, C.; Michlmayr, H.; Eder, R.; Del, H.A.; Kulbe, K.D.; Mathiesen, G.; Nguyen, T.H. Heterologous expression of Oenococcus oeni malolactic enzyme in Lactobacillus plantarum for improved malolactic fermentation. AMB Express 2012, 2, 19. [Google Scholar] [CrossRef]
- Weidmann, S.; Maitre, M.; Laurent, J.; Coucheney, F.; Rieu, A.; Guzzo, J. Production of the small heat shock protein Lo18 from Oenococcus oeni in Lactococcus lactis improves its stress tolerance. Int. J. Food Microbiol. 2017, 247, 18–23. [Google Scholar] [CrossRef]
- Costantini, A.; Rantsiou, K.; Majumder, A.; Jacobsen, S.; Pessione, E.; Svensson, B.; Garcia-Moruno, E.; Cocolin, L. Complementing DIGE proteomics and DNA subarray analyses to shed light on Oenococcus oeni adaptation to ethanol in wine-simulated conditions. J. Proteom. 2015, 123, 114–127. [Google Scholar] [CrossRef]
- Guzzo, J.; Jobin, M.P.; Delmas, F.; Fortier, L.C.; Garmyn, D.; Tourdot-Marechal, R.; Lee, B.; Divies, C. Regulation of stress response in Oenococcus oeni as a function of environmental changes and growth phase. Int. J. Food Microbiol. 2000, 55, 27–31. [Google Scholar] [CrossRef]
- Margalef-Catala, M.; Stefanelli, E.; Araque, I.; Wagner, K.; Felis, G.E.; Bordons, A.; Torriani, S.; Reguant, C. Variability in gene content and expression of the thioredoxin system in Oenococcus oeni. Food Microbiol. 2017, 61, 23–32. [Google Scholar] [CrossRef]
- Yang, K.; Dai, X.; Fan, M.; Zhang, G. Influences of acid and ethanol stresses on Oenococcus oeni SD-2a and its proteomic and transcriptional responses. J. Sci. Food Agric. 2021, 101, 2892–2900. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, H.; Peng, S.; Wang, T.; Su, J.; Liang, Y.; Li, H.; Wang, H. Transcriptomic analysis of Oenococcus oeni SD-2a response to acid shock by RNA-Seq. Front. Microbiol. 2017, 8, 1586. [Google Scholar] [CrossRef] [PubMed]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Zhang, M.M.; Zhao, X.Q.; Cheng, C.; Bai, F.W. Improved growth and ethanol fermentation of Saccharomyces cerevisiae in the presence of acetic acid by overexpression of SET5 and PPR1. Biotechnol. J. 2015, 10, 1903–1911. [Google Scholar] [CrossRef]
- Graca, D.S.M.; Vitoria, S.R.M.; Loureiro-Dias, M.C.; Rombouts, F.M.; Abee, T. Flow cytometric assessment of membrane integrity of ethanol-stressed Oenococcus oeni cells. Appl. Environ. Microbiol. 2002, 68, 6087–6093. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, J.; Wang, M.; Du, G.; Chen, J. Lactobacillus casei combats acid stress by maintaining cell membrane functionality. J. Ind. Microbiol. Biotechnol. 2012, 39, 1031–1039. [Google Scholar] [CrossRef]
- Li, Y.; Hugenholtz, J.; Sybesma, W.; Abee, T.; Molenaar, D. Using Lactococcus lactis for glutathione overproduction. Appl. Microbiol. Biotechnol. 2005, 67, 83–90. [Google Scholar] [CrossRef]
- Liu, L.; Peng, S.; Zhao, H.; Yuan, L.; Li, H.; Wang, H. Acid stress response mechanism in Oenococcus oeni SD-2a analyzed by RNA sequencing. Food Sci. 2022, 43, 153–162. [Google Scholar]
- Mendoza, S.N.; Cañón, P.M.; Contreras, Á.; Ribbeck, M.; Agosín, E. Genome-scale reconstruction of the metabolic network in Oenococcus oeni to assess wine malolactic fermentation. Front. Microbiol. 2017, 8, 534. [Google Scholar] [CrossRef]
- Chu-Ky, S.; Tourdot-Marechal, R.; Marechal, P.A.; Guzzo, J. Combined cold, acid, ethanol shocks in Oenococcus oeni: Effects on membrane fluidity and cell viability. Biochim. Biophys. Acta 2005, 1717, 118–124. [Google Scholar] [CrossRef]
- Matsumoto, S.; Breniaux, M.; Claisse, O.; Gotti, C.; Bourassa, S.; Droit, A.; Deleris-Bou, M.; Krieger, S.; Weidmann, S.; Rudolf, J.; et al. The production of preconditioned freeze-dried Oenococcus oeni primes its metabolism to withstand environmental stresses encountered upon inoculation into wine. Int. J. Food Microbiol. 2022, 369, 109617. [Google Scholar] [CrossRef]
- Zhao, H.; Li, H.; Liu, L.; Peng, S.; Wang, H. The research progress of stress adaptation mechanism of Oenococcus oeni. J. Chin. Inst. Food Sci. Technol. 2019, 19, 292–299. [Google Scholar]
- Hu, S.; Yu, Y.; Lv, Z.; Shen, J.; Ke, Y.; Xiao, X. Proteomics study unveils ROS balance in acid-adapted Salmonella enteritidis. Food Microbiol. 2020, 92, 103585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fu, R.Y.; Hugenholtz, J.; Li, Y.; Chen, J. Glutathione protects Lactococcus lactis against acid stress. Appl. Environ. Microbiol. 2007, 73, 5268–5275. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Property | Reference/Source |
|---|---|---|
| E.coli DH5α | Cloning host | Takara |
| O. oeni SD-2a | Donor bacteria | Our lab |
| L. plantarum WCFS1 | Plasmid-free bacteria | Our lab |
| WCFS1(pMG36e) | L. plantarum harboring pMG36e, Em r | Our lab |
| WCFS1(pMG36etrxA) | L. plantarum harboring pMG36etrxA, Em r | This study |
| Plasmids | ||
| pMG36e | E. coli-L. lactis shuttle vector (3,6 kb), Em r | Our lab |
| pMG36etrxA | pMG36e-derivative vector containing the 552 bp region with the ctsR gene, Em r | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Yu, X.; Wu, M.; Zhang, K.; Shang, S.; Peng, S.; Song, W. Improved Tolerance of Lactiplantibacillus plantarum in the Presence of Acid by the Heterologous Expression of trxA from Oenococcus oeni. Fermentation 2022, 8, 452. https://doi.org/10.3390/fermentation8090452
Liu L, Yu X, Wu M, Zhang K, Shang S, Peng S, Song W. Improved Tolerance of Lactiplantibacillus plantarum in the Presence of Acid by the Heterologous Expression of trxA from Oenococcus oeni. Fermentation. 2022; 8(9):452. https://doi.org/10.3390/fermentation8090452
Chicago/Turabian StyleLiu, Longxiang, Xinyu Yu, Ming Wu, Keying Zhang, Shuai Shang, Shuai Peng, and Weiyu Song. 2022. "Improved Tolerance of Lactiplantibacillus plantarum in the Presence of Acid by the Heterologous Expression of trxA from Oenococcus oeni" Fermentation 8, no. 9: 452. https://doi.org/10.3390/fermentation8090452