
The Role of Gut Microbiota Modulation Strategies in Obesity: The Applications and Mechanisms


 ,
,  ,
,
Abstract
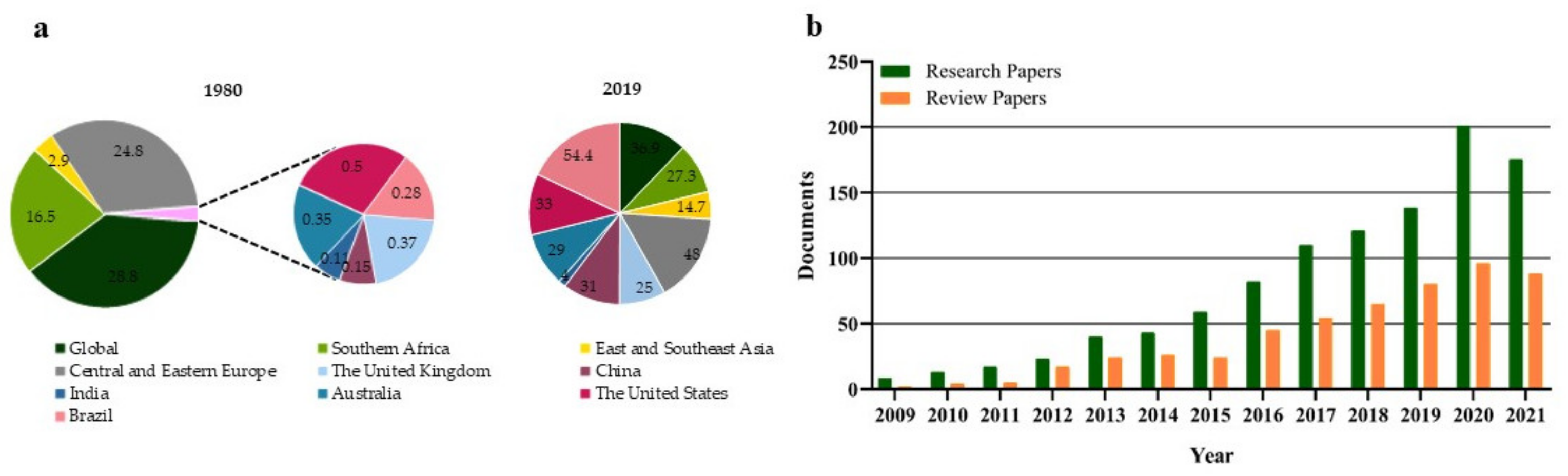
:1. Introduction
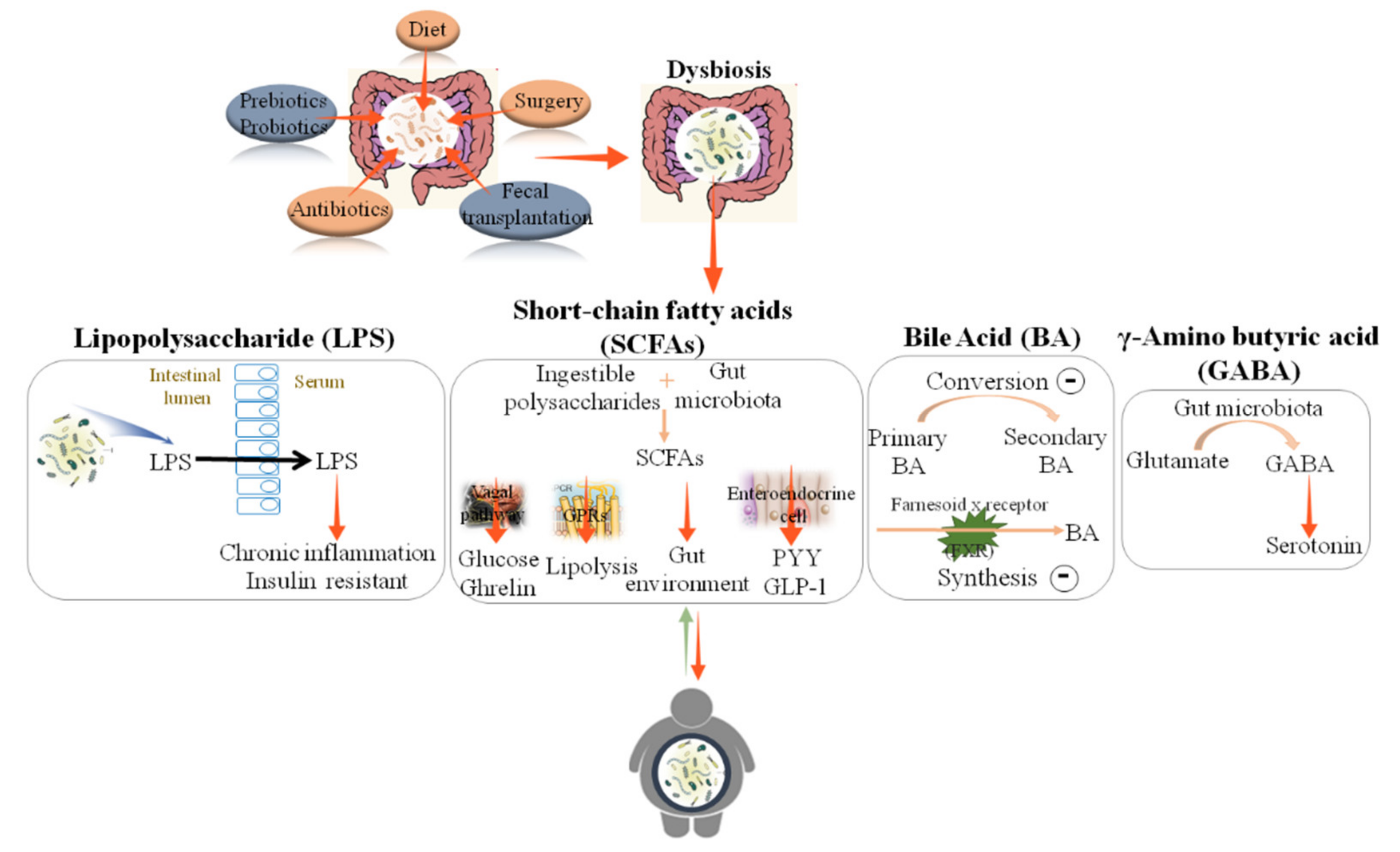
2. Gut Microbiota Dysbiosis in Obesity
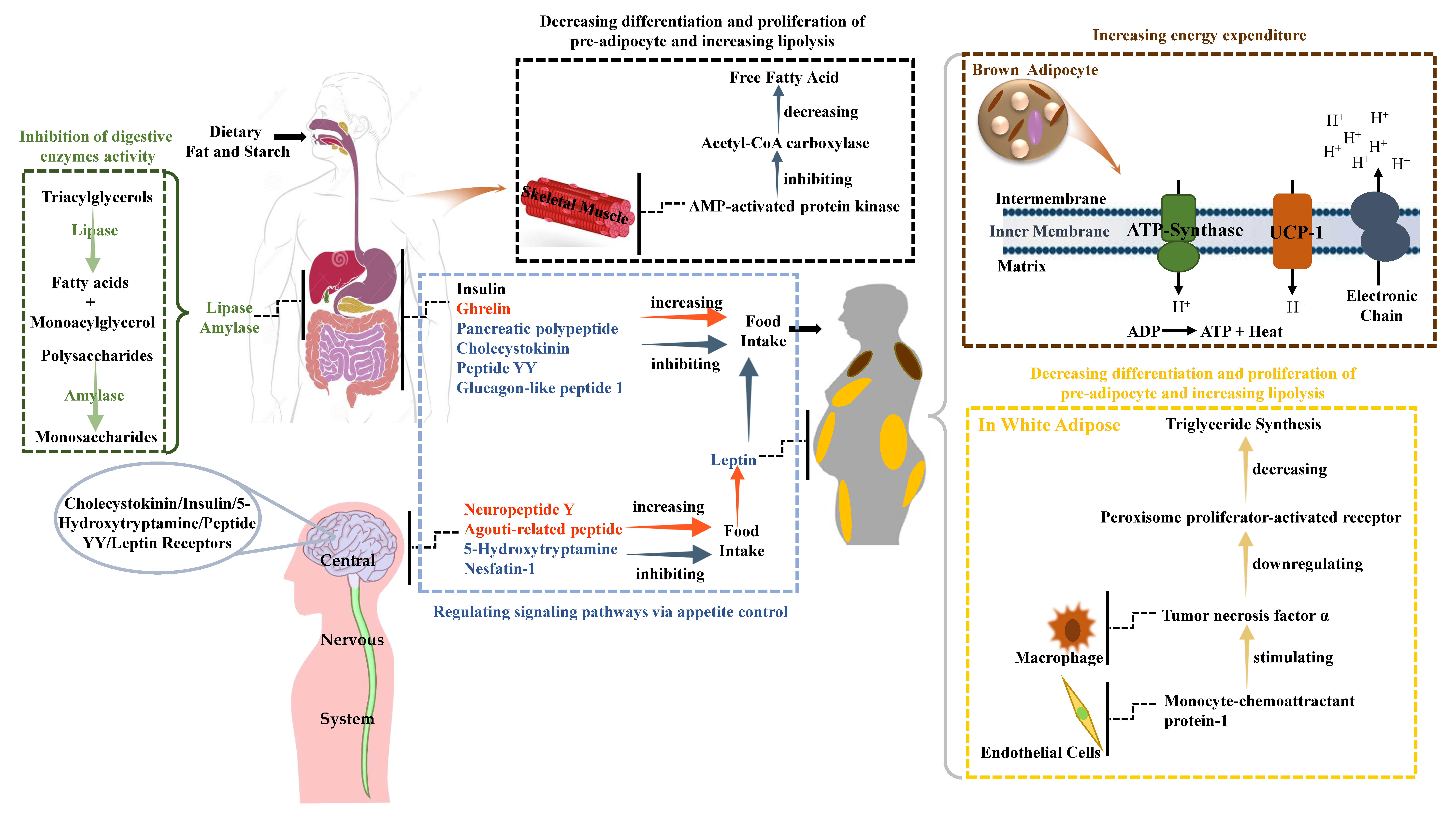
3. The Mechanistic Studies on Obesity and Related Treatment Methods
3.1. Inhibition of Digestive Enzymes Activity
3.2. Regulating Signaling Pathways via Appetite Control
3.3. Increasing Energy Expenditure
3.4. Decreasing Differentiation and Proliferation of Pre-Adipocyte and Increasing Lipolysis
4. Food-Derived Compounds in GM Modulation and Their Anti-Obesity Effects
4.1. Anti-Obesity Effect of Different Food Extracts and Gut Modulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | Experimental Results | Reference | |
|---|---|---|---|
| In Vivo/In Vitro | Gut Microbiota | ||
| Jamun | Alleviated insulin resistance, liver steatosis | Restored the Firmicutes/Bacteroidetes and SCFAs | [23] |
| Codium fragile | Decreased the size of adipocytes, levels of cholesterol and glucose | Increased the Bacteroidetes; decreased the Verrucomicrobia | [24] |
| Citrus peel | Decreased lipid content, adipocyte size, adipose tissue weight | Increased Prevotella; decreased rc4-4 bacteria | [25] |
| Aged citrus peel | Reduced the mass of adipose tissues, adipocyte size | Increased fecal SCFAs; decreased Proteobacteria, Firmicutes/Bacteroidetes | [27] |
| Blueberry | Returned lipid metabolism to normal | Modulated Proteobacteria, Deferribacteres, Actinobacteria, Bifidobacterium | [28] |
| Caffeic acid | Reduced fat accumulation; improved lipid profile | Increased the anti-obesity related and butyrate-producing bacteria | [29] |
| _ | Prevented perirenal, epididymal fat accumulation; decreased serum total cholesterol (TC), leptin and triglycerides (TG) | Decreased Firmicutes; increased Bifidobacterium, fecal acetic acid, propionic acid, total SCFAs | [30] |
| Rosadavurica Pall. | Inhibited the liver, kidney, epididymal adipose tissue weight | Decreased the Erysipelotrichaceae. | [26] |
| Green and Black tea | Decreased subcutaneous, epididymal fat | Decreased Firmicutes; increased Bacteroidetes, Pseudobutyrivibrio | [31] |
| _ | Reduced lipid levels in plasma | Inhibited Desulfovibrionaceae, Ruminococcaceae; raised the Bacteroidaceae, Lactobacillaceae | [32] |
| Capsaicin (CAP) | Lowered triglyceride, cholesterol, glucose, and insulin levels | Increased the Akkermansia, Bacteroides; reduced the Desulfovibrio, Escherichia, Helicobacter, | [33] |
| Capsicumannuum L. | Reduced the serum TG, TC, low density lipoprotein (LDL-C) levels; reversed glucose tolerance | Increased the Bifidobacterium and Akkermansia; decreased the Ruminococcus and Firmicute/Bacteroidetes | [34] |
| C.pyrenoidosa S.platensis | Protected, dyslipidemia, fat deposition; increased lipolysis and decreased lipogenesis | Increased Clostridia, Bacterioidia; decreased Actinobacteria, Verrucomicrobia; restored the SCFAs, BA | [35] |
| L.barbarum | Decreased serum TC, TG, and LDL-C | Improved community diversity of intestinal flora; could increase the content of SCFAs | [36] |
| Polymannuronic acid | Reduced the blood triacylglycerol levels and improved glucose tolerance | Increased the abundance of probiotic bacteria like Lactobacillus reuteri | [37] |
| Chitosan | Increased the serum leptin level, oral glucose tolerance | Increased Coprobacillus cateniformis; decreased Clostridium lactatifermentans | [38] |
| _ | Improved glucose intolerance | Increased the Bifidobacterium, Lachnospiraceae_NK4A136_group and Roseburia | [39] |
4.1.1. Polyphenolic Compounds
4.1.2. Polysaccharides Compounds
4.1.3. Alkaloid and Peptides
4.2. Anti-Obesity Effect of Different Food Combinations and Gut Modulations
| Food | Experimental Results | Reference | |
|---|---|---|---|
| In Vivo/In Vitro | Gut Microbiota | ||
| Euglena gracilis | Reduced visceral fat accumulation and adipocyte area | Increased SCFAs-producing beneficial bacteria; reduced harmful bacteria | [48] |
| Millet; Maize; Oat; Potato | Decreased fat accumulation; improved the blood glucose tolerance | Promoted the release of SCFAs; increased the Lactobacillus and Bifidobacterium | [49] |
| Mulberry leaf | Inhibited adipocyte differentiation and triglyceride synthesis | Reduced Firmicutes/Bacteroidetes and Lachnospiraceae; improved the Lactobacilli | [50] |
| _ | Reduced visceral adipose tissue weight, TNF-α, MCP-1, adiponectin, insulin, leptin | Decreased Desulfovibrionaceae, Coriobacteriaceae; increased Bacteroidales, Christensenellaceae, | [51] |
4.3. Anti-Obesity Effects of Fermented Foods and Gut Modulation
4.3.1. Probiotics
4.3.2. Fermented Food Materials
| Food | Experimental Results | Reference | |
|---|---|---|---|
| In Vivo/In Vitro | Gut Microbiota | ||
| C.tricuspidata | The fat mass, serum triglyceride, and alanine aminotransferase levels were reduced | Modulated Desulfovibrio, Adlercreutzia, Allobaculum, Coprococcus, Helicobacter, Flexispira, Odoribacter | [55] |
| Lactobacillusrhamnosus; L.crustorum | Mitigated insulin resistance | Decreased Bacteroides, Desulfovibrio; increased Lactobacillus, Bifdobacterium and feces SCFAs | [56] |
| Lactobacillusplantarum | Decreased deposition of fat droplets; lowered TC, TG | Increased the Bacteroidetes, Akkermansia, Lactobacillus; decreased the Firmicutes | [53] |
| Ligilactobacillus salivariu | Inhibited abdominal, liver fat accumulation, dyslipidemia | Reduced Firmicutes/Bacteroidetes; shifted overall GM structure | [54] |
| Kimchi | Decreased adipose tissue; decreased in arachidic acid, stearic acid, fumaric acid, and glucose | Muribaculaceae higher, lower Akkermansiaceae, Coriobacteriaceae, Erysipelotrichaceae | [57] |
| Andong sikhae kimchi | Reduced epididymal fat expansion, total blood cholesterol and glucose levels | Restored the Firmicutes/Bacteroidetes; increased the Bacteroides faecichinchillae and Alistipes | [58] |
| Tremella Blueberry | Dody weight (BW), TG level, fat level and ratio of lipid/BW were lower | Allobaculum, Parabacteroides, Prevotella were increased; pathogenic bacteria were reduced | [61] |
| Huyou | _ | Decreased Firmicutes/Bacteroidetes | [62] |
| Pueraria lobata | Reduced BW gain, adipocyte size | The S24_7 was increased; enriched Lactococcus and Ruminococcus | [66] |
| Panax ginseng Meyer | Decreased serum TC, LDL-C, and lipid accumulation in hepatocytes | Reduced the Firmicutes/Bacteroidetes, Muribaculaceae; increased the Prevotella_9 | [63] |
| Black barley | Inhibited liver and abdominal fat indexes; decreased thiobarbituric acid | Increased the Akkermansia, Lactococcus; Decreased Firmicutes/Bacteroidetes | [64] |
| Apples | Inhibited weight gain, fat accumulation; regulated the blood lipid levels | Decreased Firmicutes/Bacteroidotas; augmented Akkermansia, Bacteroides | [65] |
4.4. Anti-Obesity Effect of Germinated Foods with Gut Modulation
| Food | Experimental Results | Reference | |
|---|---|---|---|
| In Vivo/In Vitro | Gut Microbiota | ||
| Broccoli seeds | Reduced WAT mass, the BW and adipocyte size | Increased the Bacteroidetes; decreased the Firmicutes/Bacteroidetes | [69] |
| Black Tartary buckwheat | Decreased the serum TC, TG, and LDL-C | Increased the Lactobacillus, Faecalibaculum, Allobaculum and decreased the Romboutsia | [71] |
5. The Limitation of the Food-Derived Compounds in Modulating Gut Microbiota
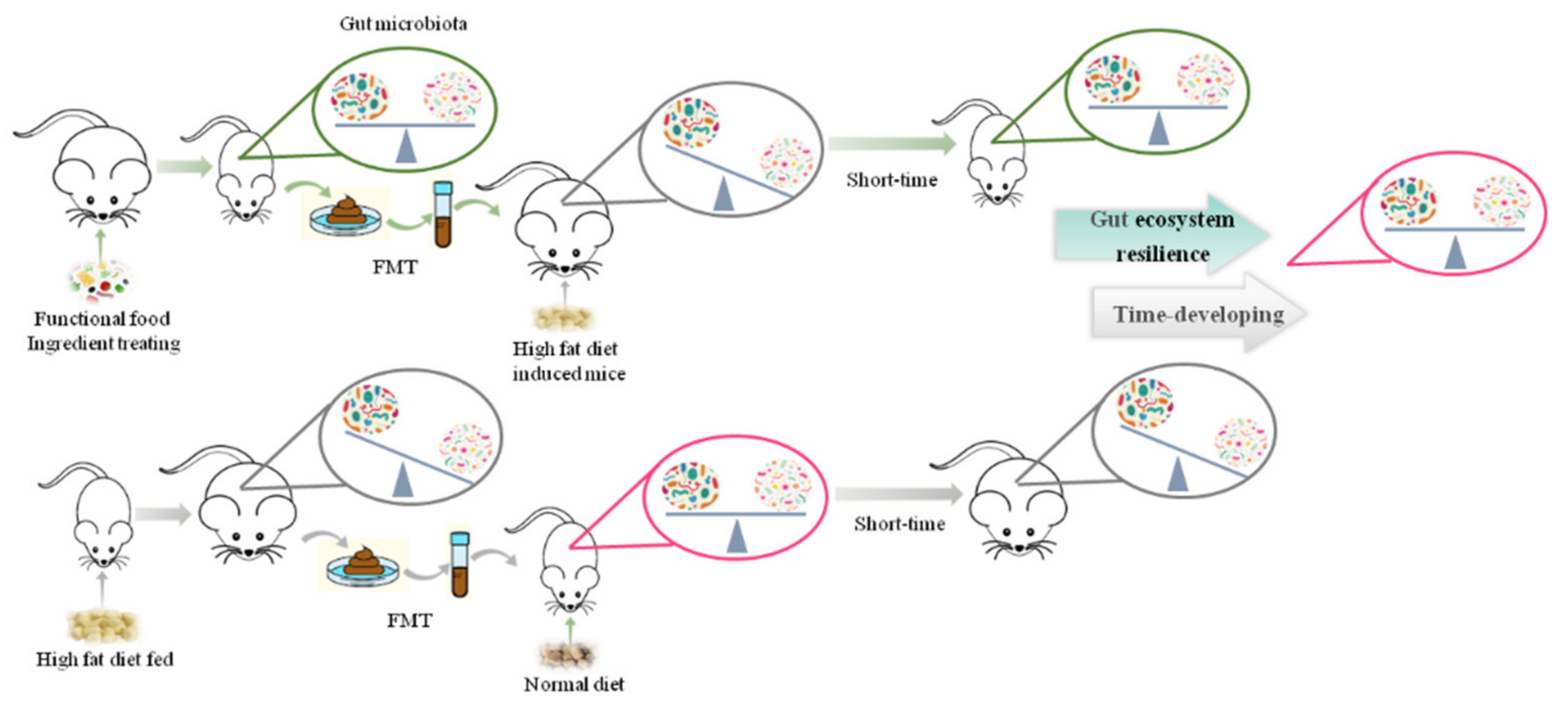
6. Combination of FMT and Dietary Compounds over Obesity
| Donors | Recipients | Methods | Results | Reference |
|---|---|---|---|---|
| Healthy lean adults | Obesity adults with insulin resistance | Weekly oral capsule administrations | Improvement in metabolism after FMT among study participants with low baseline microbiome diversity. | [85] |
| A single lean donor | Obese without metabolic syndrome | Oral capsule administrations | Patients who received FMT had sustained shifts in microbiomes toward those of the donor; BAs profiles began to resemble the donor more closely. | [86] |
| Healthy individuals from Chinese Kazaks | db/db mice | Orally | Desulfovibrio and Clostridium coccoides levels were decreased, but the fecal Akkermansia muciniphila and colon histone deacetylase-3 protein expression were increased. | [87] |
| A single healthy lean donor | Obese metabolically healthy patients | FMT capsules | There was a change in glucose and insulin under the curve compared to baseline in the FMT group. | [88] |
| Four healthy same-sex lean donors | Adolescents with obesity | FMT capsules | Altered the gut microbiome’s structure and function; two donor microbiomes dominated strain engraftment and were characterized by high Prevotella/Bacteroides. | [89] |
| Lean donor | Obesity patients with metabolic syndrome | Orally | Patients in the single-dose oral FMT combined with daily low-fermentable fiber group had significant improvements in HOMA2-IR. | [90] |
| Wild boar | Male C57BL/6N mice | Fecal suspension by oral gavage | FMT prevented HFD-induced obesity, lipid metabolism disorders; increased the Lactobacillus and Romboutsia. | [91] |
7. Future Perspective
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rodgers, J.R.; Tschop, M.H.; Wilding, J.P. Anti-obesity drugs: Past, present and future. Dis. Models Mech. 2012, 5, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.W.; Kim, S. Comprehensive review of current and upcoming anti-obesity drugs. Diabetes Metab. J. 2020, 44, 802–818. [Google Scholar] [CrossRef] [PubMed]
- Barathikannan, K.; Chelliah, R.; Rubab, M.; Daliri, E.B.; Elahi, F.; Kim, D.; Agastian, P.; Oh, S.; Oh, D.H. Gut microbiome modulation based on probiotic application for anti-obesity: A review on efficacy and validation. Microorganisms 2019, 7, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridlon, M.J.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Schellekens, H.; Dinan, T.G.; Cryan, J.F. Lean mean fat reducing “ghrelin” machine: Hypothalamic ghrelin and ghrelin receptors as therapeutic targets in obesity. Neuropharmacology 2010, 58, 2–16. [Google Scholar] [CrossRef]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Vivion, E.F.; Crowley, G.S.H.Y.; O’Rahilly, S. Obesity therapy: Altering the energy intake-and-expenditure balance sheet. Nat. Rev. Drug Discov. 2002, 1, 276–286. [Google Scholar]
- Van Den Hoek, A.M.; Heijboer, A.C.; Corssmit, E.P.; Voshol, P.J.; Romijn, J.A.; Havekes, L.M.; Pijl, H. PYY3–36 reinforces insulin action on glucose disposal in mice fed a high-fat diet. Diabetes 2004, 53, 1949–1952. [Google Scholar] [CrossRef] [Green Version]
- Keisuke Suzuki, K.A.S.; Minnion, J.S.; Shillito, J.C.; Bloom, S.R. The role of gut hormones and the hypothalamus in appetite regulation. Endocr. J. 2010, 57, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Jayasena, C.N.; Bloom, S.R. Obesity and appetite control. J. Diabetes Res. 2012, 2012, 824305. [Google Scholar] [CrossRef] [Green Version]
- Benoit, S.C.; Clegg, D.J.; Seeley, R.J.; Woods, S.C. Insulin and leptin as adiposity signals. Recent Prog. Horm. Res. 2004, 59, 267–286. [Google Scholar] [CrossRef]
- Sobrino Crespo, C.; Perianes Cachero, A.; Puebla Jimenez, L.; Barrios, V.; Arilla Ferreiro, E. Peptides and food intake. Front. Endocrinol. 2014, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Porte, D., Jr.; Baskin, D.G.; Schwartz, M.W. Leptin and insulin action in the central nervous system. Nutr. Rev. 2002, 60, S20–S29. [Google Scholar] [CrossRef] [Green Version]
- Evan, D.; Rosen, B.M.S. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar]
- Vanessa Pellegrinelli, S.C.; Antonio, V.-P. Adipose tissue plasticity: How fat depots respond differently to pathophysiological cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef] [Green Version]
- Wronska, A.; Kmiec, Z.J.A.P. Structural and biochemical characteristics of various white adipose tissue depots. Acta Physiol. 2012, 205, 194–208. [Google Scholar] [CrossRef]
- Qiong, A.; Wang, C.T.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar]
- Xia, D.-Z.; Yu, X.-F.; Wang, H.-M.; Ren, Q.-Y.; Chen, B.-M. Anti-obesity and hypolipidemic effects of ethanolic extract from Alpinia officinarum Hance (Zingiberaceae) in rats fed high-fat diet. J. Med. Food 2010, 13, 785–791. [Google Scholar] [CrossRef]
- Stuart, P.; Weisberg, D.H.; Reid, H.; Jacob, L.; Sarah, S.; Kris, V.; Israel, C.; Rudolph, L.; Leibel, A.; Ferrante, W., Jr. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J. Clin. Investig. 2006, 116, 115–124. [Google Scholar]
- Kim, J.Y.; Tillison, K.; Lee, J.H.; Rearick, D.A.; Smas, C.M. The adipose tissue triglyceride lipase ATGL/PNPLA2 is downregulated by insulin and TNF-α in 3T3-L1 adipocytes and is a target for transactivation by PPARγ. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E115–E127. [Google Scholar] [CrossRef]
- Mu, J.; Brozinick, J.T., Jr.; Valladares, O.; Bucan, M.; Birnbaum, M.J. A role for AMP-activated protein kinase in contraction-and hypoxia-regulated glucose transport in skeletal muscle. Mol. Cell 2001, 7, 1085–1094. [Google Scholar] [CrossRef]
- Kim, J.Y.; Tillison, K.; Lee, J.H.; Rearick, D.A.; Smas, C.M. Fruit Extract Prevents Obesity by Modulating the Gut Microbiome in High Fat Diet Fed Mice. Mol. Nutr. Food Res. 2019, 63, 1801307. [Google Scholar]
- Kim, J.; Choi, J.H.; Oh, T.; Ahn, B.; Unno, T. Codium fragile ameliorates high-fat diet-induced metabolism by modulating the gut microbiota in mice. Nutrients 2020, 12, 1848. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.C.; Chang, W.T.; Li, S.; Wu, J.C.; Badmeav, V.; Ho, C.T.; Pan, M.H. Citrus peel extracts attenuated obesity and modulated gut microbiota in mice with high-fat diet-induced obesity. Food Funct. 2018, 9, 3363–3373. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.Y.; Hao, Y.F.; Hao, Z.X.; Liu, Q.; Zhang, L.; Jiang, C.P.; Jiang, J.G. Flavonoids from Rosa davurica Pall. fruits prevent high-fat diet-induced obesity and liver injury via modulation of the gut microbiota in mice. Food Funct. 2021, 20, 10097–10106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, J.; Zhang, X.; Zhao, D.-G.; Ma, Y.-Y.; Li, D.; Ho, C.-T.; Huang, Q. Aged citrus peel (chenpi) extract causes dynamic alteration of colonic microbiota in high-fat diet induced obese mice. Food Funct. 2020, 11, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Wang, Y.; Lin, Y.; Lang, Y.; Li, E.; Zhang, X.; Zhang, Q.; Feng, Y.; Meng, X.; Li, B. Blueberry polyphenols extract as a potential prebiotic with anti-obesity effects on C57BL/6 J mice by modulating the gut microbiota. J. Nutr. Biochem. 2019, 64, 88–100. [Google Scholar] [CrossRef]
- Xu, J.; Ge, J.; He, X.; Sheng, Y.; Zheng, S.; Zhang, C.; Xu, W.; Huang, K. Caffeic acid reduces body weight by regulating gut microbiota in diet-induced-obese mice. J. Funct. Foods 2020, 74, 104061. [Google Scholar] [CrossRef]
- Zhu, M.-Z.; Zhou, F.; Ouyang, J.; Wang, Q.-Y.; Li, Y.-L.; Wu, J.-L.; Huang, J.-A.; Liu, Z.-H. Combined use of epigallocatechin-3-gallate (EGCG) and caffeine in low doses exhibits marked anti-obesity synergy through regulation of gut microbiota and bile acid metabolism. Food Funct. 2021, 12, 4105–4116. [Google Scholar] [CrossRef]
- Su, H.; Wang, W.J.; Zheng, G.D.; Yin, Z.P.; Li, J.E.; Chen, L.L.; Zhang, Q.F. Decaffeinated green and black tea polyphenols decrease weight gain and alter microbiome populations and function in diet-induced obese mice. Eur. J. Nutr. 2018, 57, 2759–2769. [Google Scholar]
- Wang, Z.; Lam, K.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Zeng, S.; Lin, S. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci. Nutr. Rev. 2019, 7, 579–588. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Tang, Y.; Yin, H.; Liu, X. Capsaicin has an anti-obesity effect through alterations in gut microbiota populations and short-chain fatty acid concentrations. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Gao, Y.; Hao, J.; Geng, J.; Zhang, J.; Yin, J.; Liu, R.; Sui, W.; Gong, L.; Zhang, M. Capsanthin extract prevents obesity, reduces serum TMAO levels and modulates the gut microbiota composition in high-fat-diet induced obese C57BL/6J mice. Food Res. Int. 2019, 128, 108774. [Google Scholar] [CrossRef]
- Guo, W.; Zhu, S.; Li, S.; Feng, Y.; Wu, H.; Zeng, M. Microalgae polysaccharides ameliorates obesity in association with modulation of lipid metabolism and gut microbiota in high-fat-diet fed C57BL/6 mice. Int. J. Biol. Macromol. 2021, 182, 1371–1383. [Google Scholar] [CrossRef]
- Yang, Y.; Chang, Y.; Wu, Y.; Liu, H.; Liu, Q.; Kang, Z.; Wu, M.; Yin, H.; Duan, J. A homogeneous polysaccharide from Lycium barbarum: Structural characterizations, anti-obesity effects and impacts on gut microbiota. Int. J. Biol. Macromol. 2021, 183, 2074–2087. [Google Scholar] [CrossRef]
- Liu, F.; Wang, X.; Shi, H.; Wang, Y.; Xue, C.; Tang, Q.-J. Polymannuronic acid ameliorated obesity and inflammation associated with a high-fat and high-sucrose diet by modulating the gut microbiome in a murine model. Br. J. Nutr. 2017, 117, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Wang, Y.; Kang, W.; Zhou, J.; Dong, R.; Feng, Q. Chitosan attenuates obesity by modifying the intestinal microbiota and increasing serum leptin levels in mice. J. Funct. Foods 2020, 64, 103659. [Google Scholar] [CrossRef]
- Zhang, C.; Adbo, A.A.A.; Kaddour, B.; Wu, Q.; Xin, L.; Lia, X.; Fan, G.; Teng, C. Xylan-oligosaccharides ameliorate high fat diet induced obesity and glucose intolerance and modulate plasma lipid profile and gut microbiota in mice. J. Funct. Foods 2020, 64, 103622. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Hu, T.; Zhao, H. Metabolic fate of tea polyphenols and their crosstalk with gut microbiota. Food Sci. Hum. Wellness 2022, 11, 455–466. [Google Scholar] [CrossRef]
- Su, H.; Wang, W.J.; Zheng, G.D.; Yin, Z.P.; Li, J.E.; Chen, L.L.; Zhang, Q.F. The anti-obesity and gut microbiota modulating effects of taxifolin in C57BL/6J mice fed with a high-fat diet. J. Sci. Food Agric. 2021, 102, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jeon, Y.E.; Jung, J.I.; Kim, S.M.; Hong, S.H.; Lee, J.; Hwang, J.S.; Hwang, M.O.; Kwon, K.; Kim, E.J. Anti-obesity effect of Cydonia oblonga Miller extract in high-fat diet-induced obese C57BL/6 mice. J. Funct. Foods 2022, 89, 104945. [Google Scholar] [CrossRef]
- Li, S.-Z.; Zeng, S.-L.; Liu, E.-H. Anti-obesity natural products and gut microbiota. Food Res. Int. 2022, 151, 110819. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Sun, Q.; Ma, Z.; Peng, J.; Zhang, M.; Wang, C.; Zhang, X.; Yan, X.; Chang, L.; Hou, X.; et al. Seabuckthorn polysaccharide ameliorates high-fat diet-induced obesity by gut microbiota-SCFAs-liver axis. Food Funct. 2022, 13, 2925–2937. [Google Scholar] [CrossRef]
- Li, N.; Gao, X.; Pan, Y.-Y.; Liu, B.; Pang, J.; Zhao, C.; Zeng, F. Effects of Alkaloid-Rich Extracts Obtained from Grifola frondosa on Gut Microbiota and Glucose Homeostasis in Rats. Food Funct. 2022, 13, 2729–2742. [Google Scholar] [CrossRef]
- Wang, S.; Lv, Z.; Zhao, W.; Wang, L.; He, N. Collagen peptide from Walleye pollock skin attenuated obesity and modulated gut microbiota in high-fat diet-fed mice. J. Funct. Foods 2020, 74, 104194. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, H.; Qin, N.; Ren, X.; Zhu, B.; Xia, X. Impact of food additives on the composition and function of gut microbiota: A review. Trends Food Sci. Technol. 2020, 99, 295–310. [Google Scholar] [CrossRef]
- Ran Okouchi, S.E.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K.; Tsuduki, T. Simultaneous intake of Euglena gracilis and vegetables exerts synergistic anti-obesity and anti-inflammatory effects by modulating the gut microbiota in diet-induced obese mice. Nutrients 2019, 11, 204. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Ma, N.; Zhang, J.; Wang, H.; Tao, T.; Pei, F.; Hu, Q. Dietary intake of mixture coarse cereals prevents obesity by altering the gut microbiota in high-fat diet fed mice. Food Chem. Toxicol. 2021, 147, 111901. [Google Scholar] [CrossRef]
- Liao, S.; Long, X.; Zou, Y.; Liu, F.; Li, Q. Mulberry leaf phenolics and fiber exert anti-obesity through the gut microbiota-host metabolism pathway. J. Food Sci. 2021, 86, 1432–1447. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Shabbir, U.; Arshad, M.; Sameen, A.; Oh, D.-H. Crosstalk between gut and brain in Alzheimer’s disease: The role of gut microbiota modulation strategies. Nutrients 2021, 13, 690. [Google Scholar] [CrossRef]
- Hussain, A.; Kwon, M.H.; Kim, H.K.; Lee, H.S.; Cho, J.S.; Lee, Y.I. Anti-obesity effect of Lactobacillus plantarum LB818 is associated with regulation of gut microbiota in high-fat diet-fed obese mice. J. Med. Food 2020, 23, 750–759. [Google Scholar] [CrossRef]
- Liang, C.; Zhou, X.; Jiao, Y.; Guo, M.; Meng, L.; Gong, P.; Lyu, L.; Niu, H.; Wu, Y.; Chen, S.; et al. Ligilactobacillus Salivarius LCK11 Prevents Obesity by Promoting PYY Secretion to Inhibit Appetite and Regulating Gut Microbiota in C57BL/6J Mice. Mol. Nutr. Food Res. Int. 2021, 65, 2100136. [Google Scholar] [CrossRef]
- Oh, J.K.; Vasquez, R.; Hwang, I.-C.; Na Oh, Y.; Kim, S.H.; Kang, S.H.; Joung, J.Y.; Oh, N.S.; Kim, S.; Yoon, Y.; et al. Cudrania tricuspidata Combined with Lacticaseibacillus rhamnosus Modulate Gut Microbiota and Alleviate Obesity-Associated Metabolic Parameters in Obese Mice. Microorganisms 2021, 9, 1908. [Google Scholar] [CrossRef]
- Wang, T.; Yan, H.; Lu, Y.; Li, X.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lü, X. Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation. Eur. J. Nutr. 2020, 59, 2709–2728. [Google Scholar] [CrossRef]
- Park, S.-E.; Kwon, S.J.; Cho, K.-M.; Seo, S.-H.; Kim, E.-J.; Unno, T.; Bok, S.-H.; Park, D.-H.; Son, H.-S. Intervention with kimchi microbial community ameliorates obesity by regulating gut microbiota. J. Microbiol. 2020, 58, 859–867. [Google Scholar] [CrossRef]
- Won, S.-M.; Chen, S.; Lee, S.Y.; Lee, K.E.; Park, K.W.; Yoon, J.-H. Lactobacillus sakei ADM14 Induces Anti-Obesity Effects and Changes in Gut Microbiome in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 3703. [Google Scholar] [CrossRef]
- Baruah, R.; Ray, M.; Halami, P.M. Preventive and Therapeutic aspects of Fermented Foods. J. Appl. Microbiol. 2022, 132, 3476–3489. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, X.; Zhang, L.; Deng, Z. Fermented dairy foods intake and risk of cardiovascular diseases: A meta-analysis of cohort studies. Crit. Rev. Food Sci. Nutr. 2019, 60, 1189–1194. [Google Scholar] [CrossRef]
- Zhili Sheng, L.; Li, X.; Zhao, Y.; Dai, W.; Chang, S.K.; Liu, J. The anti-obesity effect of fermented tremella/blueberry and its potential mechanisms in metabolically healthy obese rats. J. Funct. Foods 2021, 86, 104670. [Google Scholar] [CrossRef]
- Yan, X.; Wang, F.; Weng, P.; Wu, Z. The effect of fermented Huyou juice on intestinal microbiota in a high-fat diet-induced obesity mouse model. J. Food Biochem. 2020, 44, e13480. [Google Scholar] [CrossRef]
- Zhao, C.; Qu, Q.; Yang, F.; Li, Z.; Yang, P.; Han, L.; Shi, X. Monascus ruber fermented Panax ginseng ameliorates lipid metabolism disorders and modulate gut microbiota in rats fed a high-fat diet. J. Ethnopharmacol. 2021, 278, 114300. [Google Scholar] [CrossRef]
- Zhu, C.; Guan, Q.; Song, C.; Zhong, L.; Ding, X.; Zeng, H.; Nie, P.; Song, L. Regulatory effects of Lactobacillus fermented black barley on intestinal microbiota of NAFLD rats. Food Res. Int. 2021, 147, 110467. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Zhang, M.; Wang, X.; Bai, X.; Yue, T.; Gao, Z. Cloudy Apple Juice Fermented by Lactobacillus Prevents Obesity via Modulating Gut Microbiota and Protecting Intestinal Tract Health. Nutrients 2021, 13, 971. [Google Scholar] [CrossRef]
- Choi, Y.; Bose, S.; Shin, N.R.; Song, E.; Nam, Y.; Kim, H. Lactate-Fortified Puerariae radix fermented by Bifidobacterium breve improved diet-induced metabolic dysregulation via alteration of gut microbial communities. Nutrients 2020, 12, 276. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, A.; Chen, X.; Shabbir, U.; Chelliah, R.; Oh, D.H. Effect of slightly acidic electrolyzed water on amino acid and phenolic profiling of germinated brown rice sprouts and their antioxidant potential. LWT 2022, 157, 113119. [Google Scholar] [CrossRef]
- Kim, H.-J.; Eun-Ji, C.; Kim, H.S.; Choi, C.; Choi, S.; Kim, S.; Seo, W.; Do, S.H. Germinated soy germ extract ameliorates obesity through beige fat activation. Food Funct. 2019, 10, 836–848. [Google Scholar] [CrossRef]
- Li, X.; Tian, S.; Wang, Y.; Liu, J.; Wang, J.; Lu, Y. Broccoli microgreens juice reduces body weight by enhancing insulin sensitivity and modulating gut microbiota in high-fat diet-induced C57BL/6J obese mice. Eur. J. Nutr. 2021, 60, 3829–3839. [Google Scholar] [CrossRef]
- Owolabi, I.O.; Datarun, P.; Yupanqui, C.T.; Wichienchot, S. Gut microbiota metabolism of functional carbohydrates and phenolic compounds from soaked and germinated purple rice. J. Funct. Foods 2020, 66, 103787. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, S.; Xia, Y.; Huang, J.; Ye, J.; Xuan, Z.; Li, P.; Du, B. Probiotic-fermented black tartary buckwheat alleviates hyperlipidemia and gut microbiota dysbiosis in rats fed with a high-fat diet. Food Funct. 2021, 12, 6045–6057. [Google Scholar] [CrossRef] [PubMed]
- Rebecca, M.; Thomas, L.H.S. Gastrointestinal cancer. Cancer 1995, 75, 154–170. [Google Scholar]
- Glenn, R.; Gibson, M.B.R. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. Biochem. 1995, 125, 1401–1412. [Google Scholar]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Eric, C.; Martens, A.G.K.; Alexandra, S.; Tauzin, H.B. The devil lies in the details: How variations in polysaccharide fine-structure impact the physiology and evolution of gut microbes. J. Mol. Biol. 2014, 426, 3851–3865. [Google Scholar]
- Bouayed, J.; Hoffmann, L.; Bohn, T.; Phenolics, T. Flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. In vitro bioaccessibility of health-related compounds as affected by the formulation of fruit juice- and milk-based beverages. Food Res. Int. 2014, 62, 771–778. [Google Scholar] [CrossRef]
- Bárcena, C.; Valdés-Mas, R.; Mayoral, P.; Garabaya, C.; Durand, S.; Rodríguez, F.; Fernández-García, M.T.; Salazar, N.; Nogacka, A.M.; Garatachea, N.; et al. Healthspan and lifespan extension by fecal microbiota transplantation into progeroid mice. Nat. Med. 2019, 25, 1234–1242. [Google Scholar] [CrossRef]
- Lee, P.; Yacyshynm, R.B.; Mary, B. Yacyshyn Gut microbiota and obesity: An opportunity to alter obesity through faecal microbiota transplant (FMT). Diabetes Obes. Metab. 2019, 21, 479–490. [Google Scholar] [CrossRef]
- Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Karen, L.M. Impact of fecal microbiota transplantation on obesity and metabolic syndrome—A systematic review. Nutrients 2019, 11, 2291. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Liao, W.; Li, Q.; Zhang, H.; Liu, Z.; Zheng, X.; Zheng, L.; Feng, X. Interactions between resveratrol and gut microbiota affect the development of hepatic steatosis: A fecal microbiota transplantation study in high-fat diet mice. J. Funct. Foods 2020, 67, 103883. [Google Scholar] [CrossRef]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-induced gut microbiota reduces obesity in high-fat diet-fed mice. Int. J. Obes. 2019, 44, 213–225. [Google Scholar] [CrossRef]
- Morissette, A.; Kropp, C.; Songpadith, J.; Moreira, R.J.; Costa, J.; Mariné-Casadó, R.; Pilon, G.; Thibault, V.V.; Dudonné, S.; Boutekrabt, L.; et al. Blueberry proanthocyanidins and anthocyanins improve metabolic health through a gut microbiota-dependent mechanism in diet-induced obese mice. Am. J. Physiol.-Endocrinol. Metab. 2020, 318, E965–E980. [Google Scholar] [CrossRef]
- Wang, J.; Kim, S.B.; Han, K.; Kim, H. Ephedra-treated donor-derived gut microbiota transplantation ameliorates high fat diet-induced obesity in rats. Int. J. Environ. Res. Public Health 2017, 14, 555. [Google Scholar] [CrossRef] [Green Version]
- Yu, E.W.; Gao, L.; Stastka, P.; Cheney, M.C.; Mahabamunuge, J.; Soto, M.T.; Ford, C.B.; Bryant, J.A.; Henn, M.R.; Hohmann, E.L. Fecal microbiota transplantation for the improvement of metabolism in obesity: The FMT-TRIM double-blind placebo-controlled pilot trial. PLoS Med. 2020, 17, e1003051. [Google Scholar] [CrossRef]
- Jessica, R.; Allegretti, K.Z.; Benjamin, H.; Mullish, C.A.; Carrellas, M.; Hurtado, J.; Julian, R.; Marchesi, J.; McDonald, A.K.; Pechlivanis, A.; et al. Effects of fecal microbiota transplantation with oral capsules in obese patients. Clin. Gastroenterol. Hepatol. 2020, 18, 855–863. [Google Scholar]
- Zhang, P.-P.; Li, L.-L.; Han, X.; Li, Q.-W.; Zhang, X.-H.; Liu, J.J.; Wang, Y. Fecal microbiota transplantation improves metabolism and gut microbiome composition in db/db mice. Acta Pharmacol. Sin. 2020, 41, 678–685. [Google Scholar] [CrossRef]
- Jessica, R.; Allegretti, K.Z.; Jonathan, H.; Julian, R.; Marchesi, B.; Mullish, H.; Chiang, A.; Christopher, C.; Thompson, B.P. Cummings, Impact of fecal microbiota transplantation with capsules on the prevention of metabolic syndrome among patients with obesity. Hormones 2021, 20, 209–211. [Google Scholar]
- Wilson, B.C.; Vatanen, T.; Jayasinghe, T.N.; Karen, S.W.; Leong, J.G.B.; Derraik, B.; Albert, B.; Chiavaroli, V.; Darren, M.; Svirskis, K.; et al. Strain engraftment competition and functional augmentation in a multi-donor fecal microbiota transplantation trial for obesity. Microbiome 2021, 9, 107. [Google Scholar] [CrossRef]
- Mocanu, V.; Zhang, Z.; Edward, C.D.; Dina, H.; Kao, N.H.; Karmali, S.; Daniel, W.; Birch, K.; Samarasinghe, K.; Walter, J.; et al. Fecal microbial transplantation and fiber supplementation in patients with severe obesity and metabolic syndrome: A randomized double-blind, placebo-controlled phase 2 trial. Nat. Med. 2021, 27, 1272–1279. [Google Scholar] [CrossRef]
- Zhu, L.; Fu, J.; Xiao, X.; Wang, F.; Jin, M.; Fang, W.; Wang, Y.; Zong, X. Faecal microbiota transplantation-mediated jejunal microbiota changes halt high-fat diet-induced obesity in mice via retarding intestinal fat absorption. Microb. Biotechnol. 2022, 15, 337–352. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, L.; Tyagi, A.; Shabbir, U.; Chen, X.; Vijayalakshmi, S.; Yan, P.; Oh, D.-H. The Role of Gut Microbiota Modulation Strategies in Obesity: The Applications and Mechanisms. Fermentation 2022, 8, 376. https://doi.org/10.3390/fermentation8080376
Shan L, Tyagi A, Shabbir U, Chen X, Vijayalakshmi S, Yan P, Oh D-H. The Role of Gut Microbiota Modulation Strategies in Obesity: The Applications and Mechanisms. Fermentation. 2022; 8(8):376. https://doi.org/10.3390/fermentation8080376
Chicago/Turabian StyleShan, Lingyue, Akanksha Tyagi, Umair Shabbir, Xiuqin Chen, Selvakumar Vijayalakshmi, Pianpian Yan, and Deog-Hwan Oh. 2022. "The Role of Gut Microbiota Modulation Strategies in Obesity: The Applications and Mechanisms" Fermentation 8, no. 8: 376. https://doi.org/10.3390/fermentation8080376