
Selection of Producer of α-Ketoglutaric Acid from Ethanol-Containing Wastes and Impact of Cultivation Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Substrate
2.3. Medium and Cultivation Conditions
2.4. Analytical Procedures
2.5. Calculations
2.6. Statistical Analysis
3. Results and Discussion
3.1. Selection of Producer
3.2. Effect of Thiamine
3.3. Effect of Nitrogen
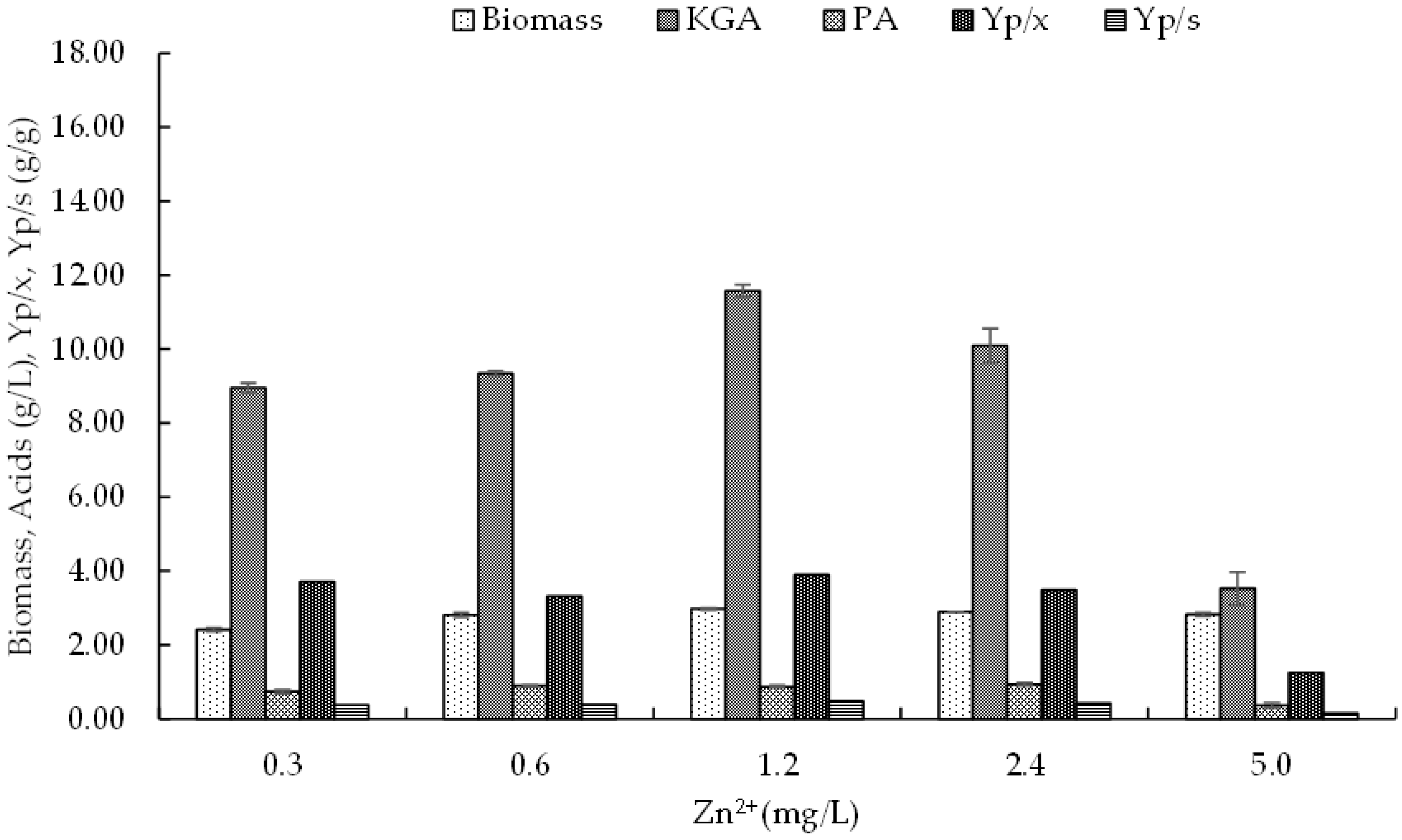
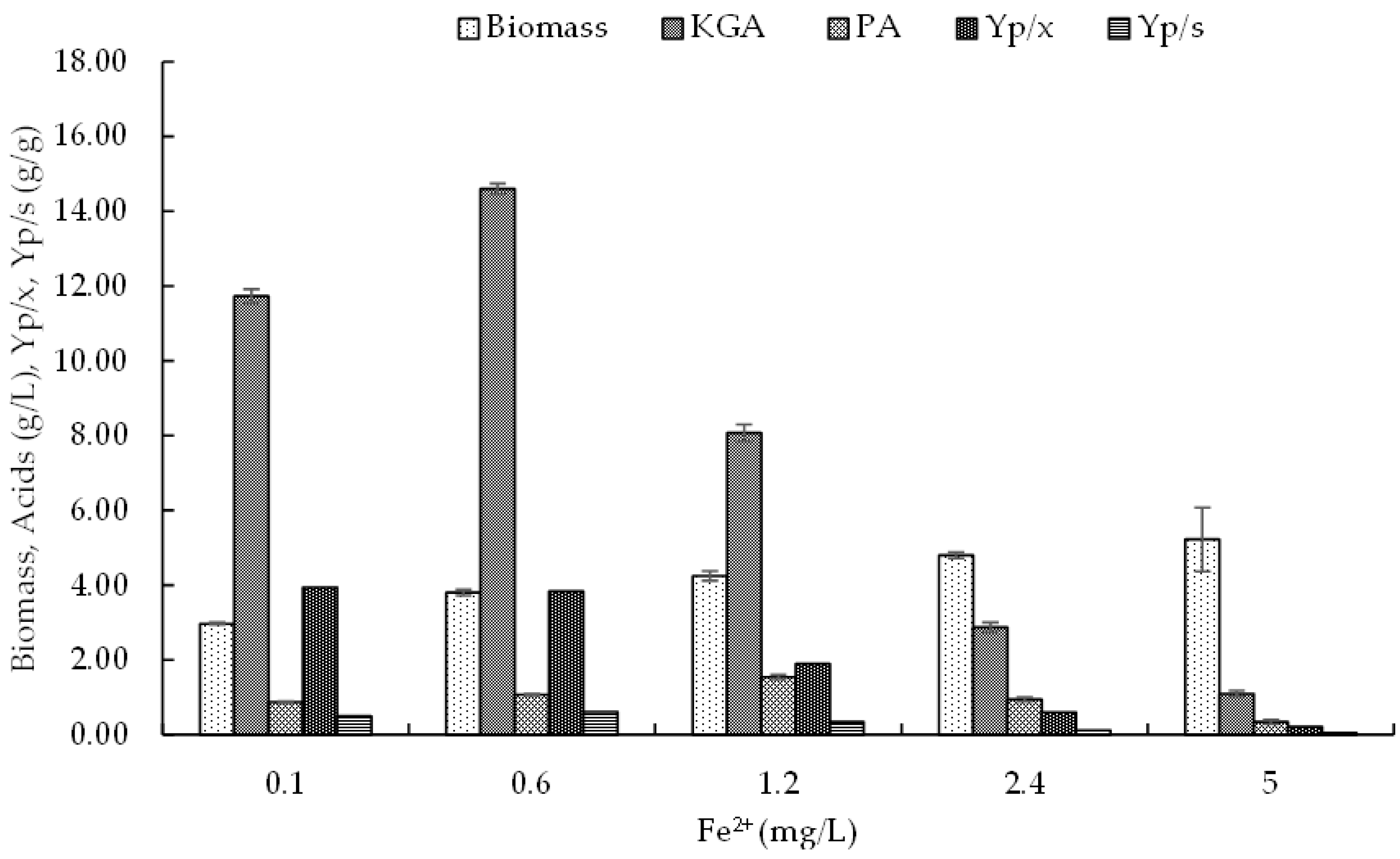
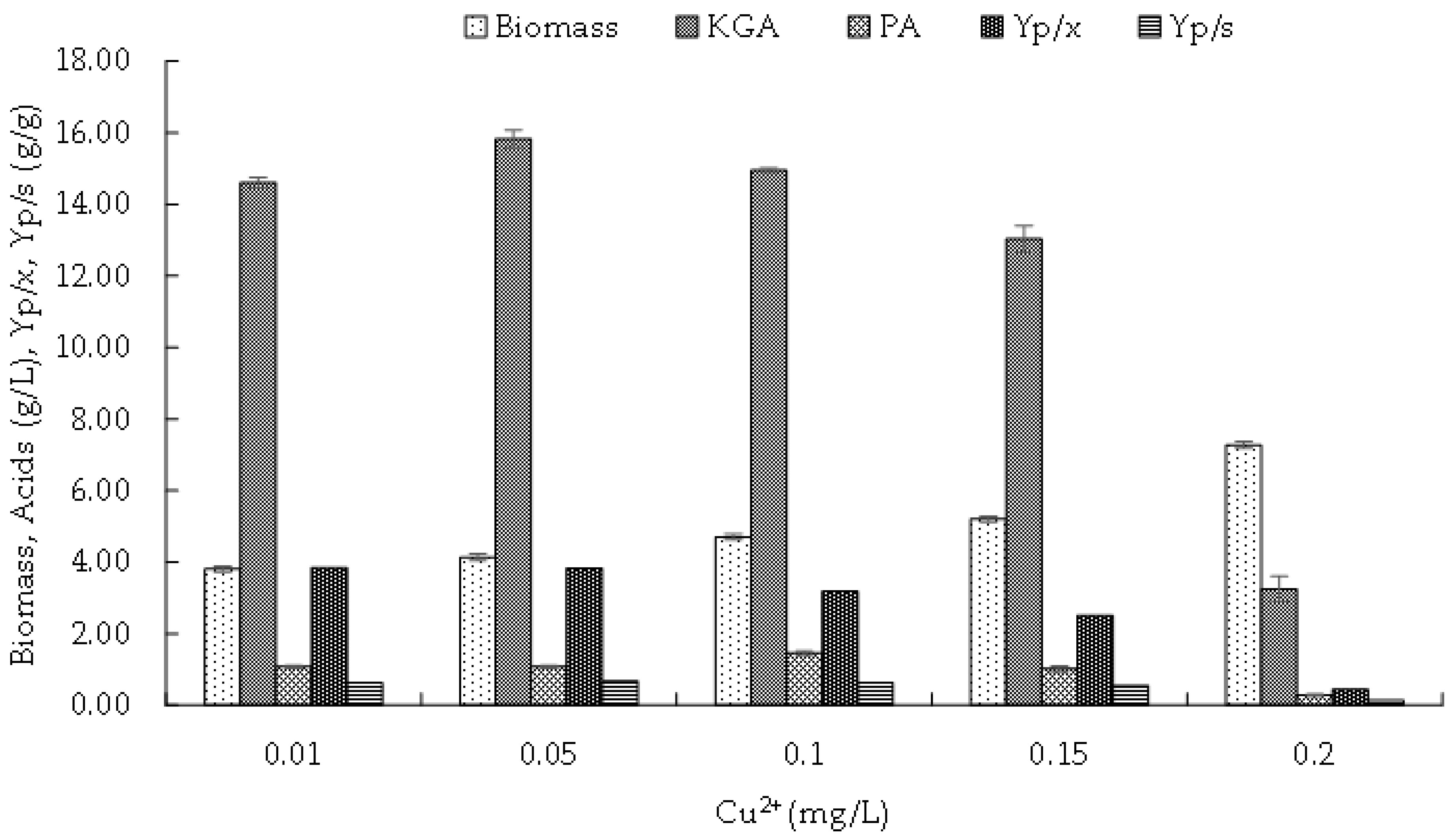
3.4. Effect of Microelements
3.4.1. Zinc
3.4.2. Iron
3.4.3. Copper
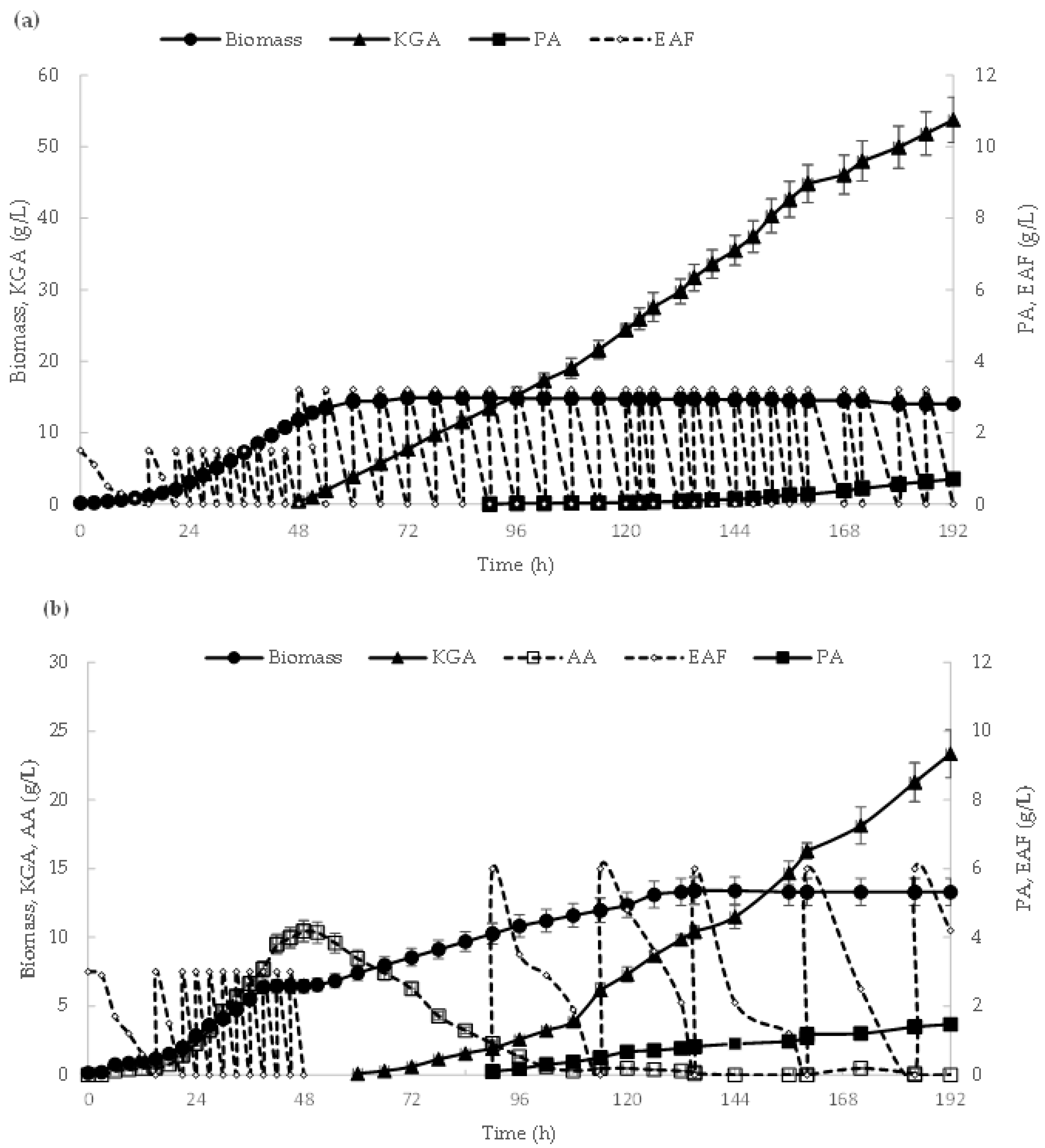
3.5. Effect of EAFs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Worldwide Production of Ethanol. Available online: https://afdc.energy.gov/data/ (accessed on 1 June 2022).
- Tsygankov, P.S.; Tsygankov, S.P. Handbook on Alcohol Rectification; Pishchepromizdat: Moscow, Russia, 2002; p. 400. (In Russian) [Google Scholar]
- Diamantis, I.; Melanouri, E.-M.; Dedousi, M.; Panagopoulou, I.; Papanikolaou, S.; Stoforos, N.G.; Diamantopoulou, P. Sustainable and Eco-Friendly Conversions of Olive Mill Wastewater-Based Media by Pleurotus pulmonarius Cultures. Fermentation 2022, 8, 129. [Google Scholar] [CrossRef]
- Cetecioglu, Z.; Atasoy, M.; Cenian, A.; Sołowski, G.; Trček, J.; Ugurlu, A.; Sedlakova-Kadukova, J. Bio-Based Processes for Material and Energy Production from Waste Streams under Acidic Conditions. Fermentation 2022, 8, 115. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Samoilenko, V.A.; Lunina, J.N.; Morgunov, I.G. Isocitric Acid Production from Ethanol Industry Waste by Yarrowia lipolytica. Fermentation 2021, 7, 146. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Karpukhina, O.V.; Kamzolova, S.V.; Samoilenko, V.A.; Inozemtsev, A.N. Investigation of the effect of biologically active threo-Ds-isocitric acid on oxidative stress in Paramecium caudatum. Prep. Biochem. Biotechnol. 2018, 48, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Morgunov, I.G.; Kamzolova, S.V.; Karpukhina, O.V.; Bokieva, S.B.; Lunina, J.N.; Inozemtsev, A.N. Microbiological production of isocitric acid from biodiesel waste and its effect on spatial memory. Microorganisms 2020, 8, 462. [Google Scholar] [CrossRef] [Green Version]
- Finogenova, T.V.; Morgunov, I.G.; Kamzolova, S.V.; Chernyavskaya, O.G. Organic acid production by the yeast Yarrowia lipolytica: A review of prospects. Appl. Biochem. Microbiol. 2005, 41, 418–425. [Google Scholar] [CrossRef]
- Stottmeister, U.; Aurich, A.; Wilde, H.; Andersch, J.; Schmidt, S.; Sicker, D. White biotechnology for green chemistry: Fermentative 2-oxocarboxylic acids as novel building blocks for subsequent chemical syntheses. J. Ind. Microbiol. Biotech. 2005, 32, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Yovkova, V.; Barth, G. Overproduction and secretion of α-ketoglutaric acid by microorganisms. Appl. Microbiol. Biotechnol. 2011, 92, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Aurich, A.; Specht, R.; Müller, R.A.; Stottmeister, U.; Yovkova, V.; Otto, C.; Holz, M.; Barth, G.; Heretsch, P.; Thomas, F.A.; et al. Microbiologically produced carboxylic acids used as building blocks in organic synthesis. In Reprogramming Microbial Metabolic Pathways; Wang, X., Chen, J., Quinn, P., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2012; Volume 64, pp. 391–423. [Google Scholar]
- Song, Y.; Li, J.; Shin, H.; Liu, L.; Du, G.; Chen, J. Biotechnological production of alpha-keto acids: Current status and perspectives. Bioresour. Technol. 2016, 219, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Su, S.; Madzak, C.; Zhou, J.; Chen, H.; Chen, G. Applying pathway engineering to enhance production of alpha-ketoglutarate in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2016, 100, 9875–9884. [Google Scholar] [CrossRef]
- Liu, S.; He, L.; Yao, K. The antioxidative function of alpha-ketoglutarate and its applications. BioMed Res. Int. 2018, 2018, 3408467. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Cheng, H.; Sze Ki Lin, C. Sugar Alcohols and Organic Acids Synthesis in Yarrowia lipolytica: Where Are We? Microorganisms 2020, 8, 574. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liu, H.; Peng, L.; He, W.; Li, S. Potential clinical applications of alpha-ketoglutaric acid in diseases. Mol. Med. Rep. 2022, 25, 151. [Google Scholar] [CrossRef]
- Luo, Z.; Yu, S.; Zeng, W.; Zhou, J. Comparative analysis of the chemical and biochemical synthesis of keto acids. Biotechnol. Adv. 2021, 47, 107706. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhou, H.; Du, G.; Liu, L.; Chen, J. Screening of a thiamine-auxotrophic yeast for alpha-ketoglutaric acid overproduction. Lett. Appl. Microbiol. 2010, 51, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Holz, M.; Otto, C.; Kretzschmar, A.; Yovkova, V.; Aurich, A.; Pötter, M.; Marx, A.; Barth, G. Overexpression of alpha-ketoglutarate dehydrogenase in Yarrowia lipolytica and its effect on production of organic acids. Appl. Microbiol. Biotechnol. 2011, 89, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yin, X.; Madzak, C.; Du, G.; Chen, J. Enhanced α-ketoglutarate production in Yarrowia lipolytica WSH-Z06 by alteration of the acetyl-CoA metabolism. J. Biotechnol. 2012, 161, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Enhanced alpha-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by regulation of the pyruvate carboxylation pathway. Appl. Microbiol, Biotechnol. 2012, 96, 1527–1537. [Google Scholar] [CrossRef]
- Yu, Z.; Du, G.; Zhou, J.; Chen, J. Enhanced α-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by an improved integrated fed-batch strategy. Bioresour. Technol. 2012, 114, 597–602. [Google Scholar] [CrossRef]
- Guo, H.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Effects of pyruvate dehydrogenase subunits overexpression on the α-ketoglutarate production in Yarrowia lipolytica WSH-Z06. Appl. Microbiol. Biotechnol. 2014, 98, 7003–7012. [Google Scholar] [CrossRef]
- Guo, H.; Liu, P.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Identification and application of keto acids transporters in Yarrowia lipolytica. Sci. Rep. 2015, 5, 8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Du, G.; Chen, J.; Li, J.; Zhou, J. A high-throughput screening procedure for enhancing α-ketoglutaric acid production in Yarrowia lipolytica by random mutagenesis. Process Biochem. 2015, 50, 1516–1522. [Google Scholar] [CrossRef]
- Otto, C.; Yovkova, V.; Aurich, A.; Mauersberger, S.; Barth, G. Variation of the by-product spectrum during α-ketoglutaric acid production from raw glycerol by overexpression of fumarase and pyruvate carboxylase genes in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 95, 905–917. [Google Scholar] [CrossRef]
- Yovkova, V.; Otto, C.; Aurich, A.; Mauersberger, S.; Barth, G. Engineering the α-ketoglutarate overproduction from raw glycerol by overexpression of the genes encoding NADP+-dependent isocitrate dehydrogenase and pyruvate carboxylase in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2014, 98, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Fang, F.; Liu, S.; Du, G.; Chen, J.; Zhou, J. Comparative genomics analysis of a series of Yarrowia lipolytica WSH-Z06 mutants with varied capacity for α-ketoglutarate production. J. Biotechnol. 2016, 239, 76–82. [Google Scholar] [CrossRef]
- Zeng, W.; Zhang, H.; Xu, S.; Fang, F.; Zhou, J. Biosynthesis of keto acids by fed-batch culture of Yarrowia lipolytica WSH-Z06. Bioresour. Technol. 2017, 243, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Morgunov, I.G. Optimization of medium composition and fermentation conditions for α-ketoglutaric acid production from biodiesel waste by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2020, 104, 7979–7989. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Samoilenko, V.A. Enhanced α-ketoglutaric acid production and recovery in Yarrowia lipolytica yeast by effective pH controlling. Appl. Microbiol. Biotechnol. 2013, 97, 8711–8718. [Google Scholar] [CrossRef]
- Cybulski, K.; Tomaszewska-Hetman, L.; Rakicka, M.; Laba, W.; Rymowicz, W.; Rywinska, A. The bioconversion of waste products from rapeseed processing into keto acids by Yarrowia lipolytica. Ind. Crops Prod. 2018, 119, 102–110. [Google Scholar]
- Rywińska, A.; Tomaszewska-Hetman, L.; Rakicka-Pustułka, M.; Juszczyk, P.; Rymowicz, W. Alpha-ketoglutaric acid production from a mixture of glycerol and rapeseed oil by Yarrowia lipolytica using different substrate feeding strategies. Sustainability 2020, 12, 6109. [Google Scholar] [CrossRef]
- Tomaszewska-Hetman, L.; Rywińska, A.; Lazar, Z.; Juszczyk, P.; Rakicka-Pustułka, M.; Janek, T.; Kuźmińska-Bajor, M.; Rymowicz, W. Application of a New Engineered Strain of Yarrowia lipolytica for Effective Production of Calcium Ketoglutarate Dietary Supplements. Int. J. Mol. Sci. 2021, 22, 7577. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Labbé, N.; Trinh, C.T. Simultaneous saccharification and fermentation of cellulose in ionic liquid for efficient production of α-ketoglutaric acid by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2015, 99, 4237–4244. [Google Scholar] [CrossRef] [PubMed]
- Chernyavskaya, O.G.; Shishkanova, N.V.; Il’chenko, A.P.; Finogenova, T.V. Synthesis of alpha ketoglutaric acid by Yarrowia lipolytica yeast grown on ethanol. Appl. Microbiol. Biotechnol. 2000, 53, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Chiglintseva, M.N.; Lunina, J.N.; Morgunov, I.G. α-Ketoglutaric acid production by Yarrowia lipolytica and its regulation. Appl. Microbiol. Biotechnol. 2012, 96, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, I.T.; Müller, P.; Finogenova, T.V.; Lozinov, A.B. Kinetics of Candida lipolytica yeast growth and biosynthesis of alpha-keto acids with thiamine deficiency in media with different carbon sources. Microbiologiia 1979, 48, 849–853. [Google Scholar]
- Finogenova, T.V. Biosynthesis of Organic Acids by Yeast Organisms and Its Regulation. Ph.D. Thesis, USSR Academy of Sciences, Institute of Biochemistry and Physiology of Microorganisms, Pushchino, Russia, 1982. (In Russian). [Google Scholar]
- Ermakova, I.T.; Finogenova, T.V.; Lozinov, A.B. Effect of cultivation conditions on the growth of yeast C. lipolytica and the biosynthesis of α-ketoglutaric acids under conditions of thiamine deficiency. Microbiologiia 1979, 48, 1001–1010. [Google Scholar]
- Ito, H.; Inouhe, M.; Tohoyama, H.; Joho, M. Characteristics of copper tolerance in Yarrowia lipolytica. BioMetals 2007, 20, 773–780. [Google Scholar] [CrossRef]
- Karasu-Yalcin, S.; Tijen Bozdemir, M.; Yesim Ozbas, Z. Effects of different fermentation conditions on growth and citric acid production kinetics of two Yarrowia lipolytica strains. Chem. Biochem. Eng. Q. 2010, 24, 347–360. [Google Scholar]
- Tomaszewska, L.; Rymowicz, W.; Rywinska, A. Mineral supplementation increases erythrose reductase activity in erythritol biosynthesis from glycerol by Yarrowia lipolytica. Appl. Biochem. Biotechnol. 2014, 172, 3069–3078. [Google Scholar] [CrossRef] [Green Version]
- Jach, M.E.; Malm, A. Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans. Molecules 2022, 27, 2300. [Google Scholar] [CrossRef]
- Mackenzie, K.F.; Eddy, C.K.; Ingram, L.O. Modulation of alcohol dehydrogenase isoenzyme levels in Zymomonas mobilis by iron and zinc. J. Bacteriol. 1989, 171, 1063–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, G.-H.; Hsieh, M.-C.; Shu, H.-Y. Role of Iron-Containing Alcohol Dehydrogenases in Acinetobacter baumannii ATCC 19606 Stress Resistance and Virulence. Int. J. Mol. Sci. 2021, 22, 9921. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.C.; Dawson, P.S.S. Relationship between iron-limited growth and energy limitation during phased cultivation of Candida utilis. Can. J. Microbiol. 1978, 24, 440–447. [Google Scholar] [CrossRef]
- Hoffmann, A.; Kupsch, C.; Walther, T.; Löser, C. Synthesis of ethyl acetate from glucose by Kluyveromyces marxianus, Cyberlindnera jadinii and Wickerhamomyces anomalus depending on the induction mode. Eng. Life Sci. 2021, 21, 154–168. [Google Scholar] [CrossRef]
- Il’chenko, A.P.; Chernyavskaya, O.G.; Finogenova, T.V. Ethanol metabolism in the yeasts Yarrowia and Torulopsis: A Review. Appl. Biochem. Microbiol. 2005, 41, 426–432. [Google Scholar] [CrossRef]
- Rodrigues, G.; Pais, C. The influence of acetic and other weak carboxylic acids on growth and cellular death of the yeast Yarrowia lipolytica. Food Technol. Biotechnol. 2000, 38, 27–32. [Google Scholar]
- Augstein, A.; Barth, K.; Gentsch, M.; Kohlwein, S.D.; Barth, G. Characterization, localization and functional analysis of Gpr1p, a protein affecting sensitivity to acetic acid in the yeast Yarrowia lipolytica. Microbiology 2003, 149, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Chatzifragkou, A.; Fakas, S.; Galiotou-Panayotou, M.; Komaitis, M.; Nicaud, J.M.; Aggelis, G. Biosynthesis of lipids and organic acids by Yarrowia lipolytica strains cultivated on glucose. Eur. J. Lipid Sci. Technol. 2009, 111, 1221–1232. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Makri, A.; Belka, A.; Bellou, S.; Mavrou, M.; Mastoridou, M.; Mystrioti, P.; Onjaro, G.; Aggelis, G.; Papanikolaou, S. Biotechnological conversions of biodiesel derived waste glycerol by yeast and fungal species. Energy 2011, 36, 1097–1108. [Google Scholar] [CrossRef]
- Tovilla-Coutiño, D.B.; Momany, C.; Eiteman, M.A. Engineered citrate synthase alters acetate accumulation in Escherichia coli. Metab. Eng. 2020, 61, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Zinjarde, S.S. Food-related applications of Yarrowia lipolytica. Food Chem. 2014, 152, 1–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Thiamine (µg/L) | Biotin (µg/L) | Biomass (g/L) | KGA (g/L) | PA (g/L) | KGA (% from Total Acids) | Yp/x (g/g) | Yp/s (g/g) |
|---|---|---|---|---|---|---|---|---|
| Ambrosiozyma angophorae VKM Y-2218 | 0.5 | 20 | 0.95 ± 0.07 | 0.49 ± 0.02 | 0.04 ± 0.0 | 92.59 | 0.51 | 0.02 |
| Babjeviella inositovora VKM Y-2494 | 0.5 | 20 | 5.42 ± 0.4 | 5.31 ± 0.83 | 0.63 ± 0.1 | 89.45 | 0.98 | 0.22 |
| Candida tropicalis (Castellani) 303 | - | 20 | 9.41 ± 0.7 | 0 | 0 | nd | nd | nd |
| C. maltosa | - | 20 | 9.50 ± 0.71 | 0 | 0 | nd | nd | nd |
| C. valida | - | - | 9.31 ± 0.69 | 0 | 0 | nd | nd | nd |
| Debaryomyces tamarii VKM Y-2626 | 0.5 | 20 | 7.89 ± 0.59 | 0.10 ± 0.0 | 0.10 ± 0.0 | 50.00 | 0.01 | nd |
| Diutina catenulata VKM Y-5 | 0.5 | 20 | 7.13 ± 0.53 | 2.31 ± 0.27 | 1.08 ± 0.17 | 68.17 | 0.32 | 0.10 |
| D. catenulata VKM Y-65 | 0.5 | 20 | 7.79 ± 0.58 | 2.83 ± 0.10 | 0.15 ± 0.01 | 95.08 | 0.36 | 0.12 |
| D.rugosa VKM Y-67 | 0.5 | 20 | 8.10 ± 1.27 | 2.45 ± 0.49 | 0.20 ± 0.01 | 92.63 | 0.30 | 0.10 |
| Moesziomyces antarcticus VKM Y-2604 | 0.5 | - | 8.55 ± 0.64 | 4.00 ± 0.14 | 0.20 ± 0.01 | 95.35 | 0.47 | 0.17 |
| Moesziomyces aphidis VKM Y-2090 | 0.5 | - | 1.81 ± 0.13 | 1.17 ± 0.04 | 0.59 ± 0.02 | 66.67 | 0.65 | 0.05 |
| Pichia besseyi VKM Y-2084 | 0.5 | - | 8.10 ± 1.13 | 5.07 ± 0.18 | 0.20 ± 0.01 | 96.30 | 0.63 | 0.21 |
| Pichia fermentans VKM Y-293 | 0.5 | - | 3.64 ± 0.51 | 0.88 ± 0.03 | 0.20 ± 0.01 | 81.82 | 0.24 | 0.04 |
| Pichia haplophila VKM Y-1286 | 0.5 | 20 | 2.37 ± 0.33 | 0.06 ± 0.00 | 0.07 ± 0.00 | 46.15 | 0.02 | nd |
| Pichia media VKM Y-1381 | 0.5 | 20 | 7.41 ± 0.55 | 4.97 ± 0.18 | 0.24 ± 0.01 | 95.33 | 0.67 | 0.21 |
| Saturnispora dispora VKM Y-418 | 0.5 | - | 7.64 ± 1.07 | 5.50 ± 0.71 | 0.20 ± 0.01 | 96.58 | 0.72 | 0.23 |
| Sugiyamaella paludigena VKM Y-2443 | 0.5 | - | 5.99 ± 0.45 | 4.00 ± 0.14 | 1.56 ± 0.06 | 71.93 | 0.67 | 0.17 |
| Yarrowia lipolytica VKM Y-2373 | 0.5 | - | 4.00 ± 0.56 | 4.97 ± 0.18 | 0.59± 0.02 | 89.47 | 1.24 | 0.21 |
| Y. lipolytica VKM Y-2412 | 0.5 | - | 3.42 ± 0.25 | 8.00 ± 0.29 | 0.78 ± 0.03 | 91.11 | 2.34 | 0.34 |
| Y. lipolytica 47 | 0.5 | - | 3.02 ± 0.42 | 5.45 ± 0.20 | 0.20 ± 0.01 | 96.55 | 1.80 | 0.23 |
| Y. lipolytica 69 | 0.5 | - | 2.73 ± 0.38 | 3.32 ± 0.12 | 0.07± 0.00 | 97.98 | 1.21 | 0.14 |
| Y. lipolytica 374 | 0.5 | - | 2.74 ± 0.20 | 3.67 ± 0.13 | 0.20 ± 0.01 | 94.95 | 1.34 | 0.15 |
| Y. lipolytica 387 | 0.5 | - | 2.85 ± 0.40 | 3.59 ± 0.13 | 0.29 ± 0.01 | 92.46 | 1.26 | 0.15 |
| Y. lipolytica 581 | 0.5 | - | 3.33 ± 0.25 | 4.76 ± 0.17 | 0 | 100.00 | 1.43 | 0.20 |
| Y. lipolytica 607 | 0.5 | - | 3.56 ± 0.27 | 3.39 ± 0.12 | 0.20 ± 0.01 | 94.57 | 0.95 | 0.14 |
| Y. lipolytica 645 | 0.5 | - | 3.98 ± 0.30 | 2.95 ± 0.11 | 0.07 ± 0.00 | 97.74 | 0.74 | 0.12 |
| Y. lipolytica 681 | 0.5 | - | 3.41 ± 0.48 | 2.67 ± 0.10 | 0.59 ± 0.02 | 82.04 | 0.78 | 0.11 |
| Zygoascus hellenicus VKM Y-2154 | 0.5 | - | 1.09 ± 0.15 | 2.24 ± 0.08 | 0.29 ± 0.01 | 88.46 | 2.05 | 0.09 |
| Thiamine (µg/L) | Biomass (g/L) | KGA (g/L) | PA (g/L) | KGA (% from Total Acids) | Yp/x (g/g) | Yp/s (g/g) |
|---|---|---|---|---|---|---|
| 0 | 1.70 ± 0.08 | 4.21 ± 0.20 | 0.00 ± 0.01 | 100.00 | 2.48 | 0.18 |
| 0.1 | 2.10 ± 0.08 | 5.84 ± 0.15 | 0.13 ± 0.01 | 99.90 | 2.78 | 0.25 |
| 0.3 | 2.40 ± 0.17 | 8.95 ± 0.10 | 0.76 ± 0.04 | 92.18 | 3.73 | 0.38 |
| 0.5 | 3.55 ± 0.27 | 8.07 ± 0.19 | 0.81 ± 0.05 | 90.85 | 2.27 | 0.34 |
| 1.0 | 4.21 ± 0.17 | 4.38 ± 0.10 | 0.43 ± 0.04 | 91.05 | 1.04 | 0.18 |
| 1.5 | 4.87 ± 0.29 | 3.04 ± 0.31 | 0.31 ± 0.02 | 90.81 | 0.62 | 0.13 |
| 2.0 | 6.18 ± 0.19 | 1.70 ± 0.17 | 0.15 ± 0.02 | 91.77 | 0.27 | 0.07 |
| 2.5 | 6.47 ± 0.37 | 0.51 ± 0.03 | 0.03 ± 0.03 | 95.31 | 0.08 | 0.02 |
| 5.0 | 6.94 ± 0.27 | 0 | 0 | nd | nd | nd |
| 10.0 | 8.37 ± 0.30 | 0 | 0 | nd | nd | nd |
| (NH4)2SO4 (g/L) | Nitrogen (g/L) | C/N Ratio | Biomass (g/L) | KGA (g/L) | PA (g/L) | N content (mg/g) | KGA (% from Total Acids) | Yp/x (g/g) | Yp/s (g/g) |
|---|---|---|---|---|---|---|---|---|---|
| 0.3 | 0.06 | 197 | 2.16 ± 0.14 | 3.49 ± 0.03 | 0.04 ± 0.01 | 30 | 92.02 | 0.23 | 0.02 |
| 0.5 | 0.11 | 119 | 2.29 ± 0.17 | 3.37 ± 0.26 | 0.38 ± 0.05 | 50 | 89.98 | 1.47 | 0.14 |
| 1.0 | 0.21 | 59 | 2.35 ± 0.13 | 8.53 ± 0.28 | 0.68 ± 0.06 | 90 | 92.66 | 3.63 | 0.35 |
| 2.0 | 0.42 | 30 | 2.36 ± 0.11 | 8.81 ± 0.14 | 0.70 ± 0.04 | 100 | 92.62 | 3.73 | 0.37 |
| 3.0 | 0.64 | 20 | 2.40 ± 0.17 | 8.95 ± 0.10 | 0.76 ± 0.04 | 110 | 92.18 | 3.73 | 0.37 |
| 4.0 | 0.85 | 15 | 2.88 ± 0.13 | 6.89 ± 0.35 | 0.52 ± 0.09 | 220 | 93.01 | 2.40 | 0.29 |
| 6.0 | 1.27 | 10 | 3.15 ± 0.13 | 3.31 ± 0.21 | 0.27 ± 0.07 | 340 | 92.51 | 1.05 | 0.14 |
| 8.0 | 1.70 | 7 | 2.25 ± 0.14 | 2.68 ± 0.10 | 0.27 ± 0.05 | 680 | 90.75 | 1.19 | 0.11 |
| 10.0 | 2.12 | 6 | 2.03 ± 0.13 | 2.53 ± 0.12 | 0.23 ± 0.07 | 970 | 91.73 | 1.25 | 0.10 |
| Type of Feeding | Yp/x (g/g) | Yp/s (g/g) | Qp (g/L·h) |
|---|---|---|---|
| Small-dose feeding | 3.84 | 0.68 | 0.38 |
| Large-dose feeding | 1.76 | 0.52 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamzolova, S.V.; Morgunov, I.G. Selection of Producer of α-Ketoglutaric Acid from Ethanol-Containing Wastes and Impact of Cultivation Conditions. Fermentation 2022, 8, 362. https://doi.org/10.3390/fermentation8080362
Kamzolova SV, Morgunov IG. Selection of Producer of α-Ketoglutaric Acid from Ethanol-Containing Wastes and Impact of Cultivation Conditions. Fermentation. 2022; 8(8):362. https://doi.org/10.3390/fermentation8080362
Chicago/Turabian StyleKamzolova, Svetlana V., and Igor G. Morgunov. 2022. "Selection of Producer of α-Ketoglutaric Acid from Ethanol-Containing Wastes and Impact of Cultivation Conditions" Fermentation 8, no. 8: 362. https://doi.org/10.3390/fermentation8080362