
Halophytes as Feedstock for Biogas Production: Composition Analysis and Biomethane Potential of Salicornia spp. Plant Material from Hydroponic and Seawater Irrigation Systems

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
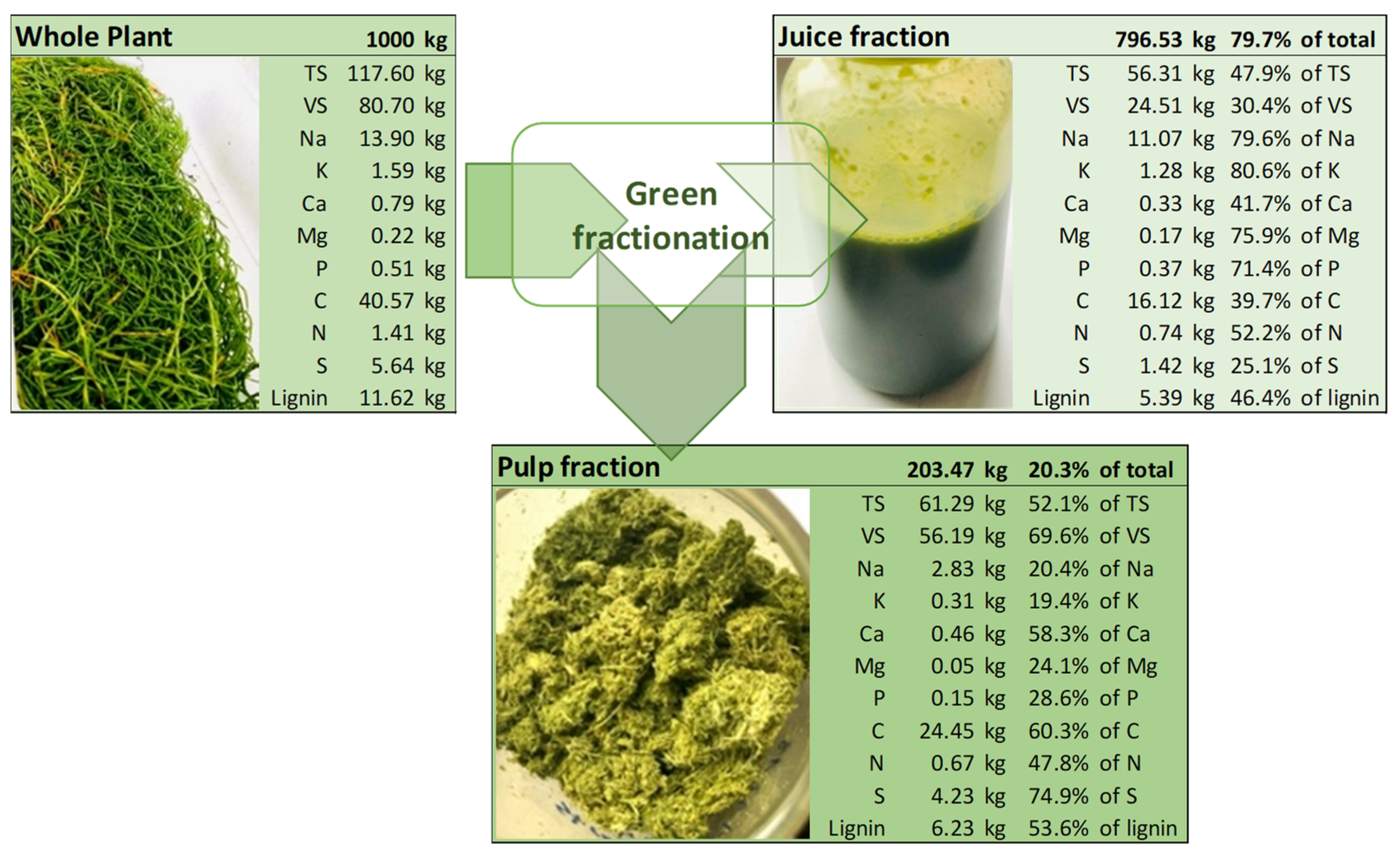
2.1. Halophyte Plant Material and Residues Used for the Biogas Potential Tests
2.2. Biomass Composition Analysis
2.2.1. Content of Carbohydrates, Lignin, and Proteins
2.2.2. Elemental Analysis
2.2.3. Mineral Content in Substrates and Inoculum
2.3. Theoretical BMP and Biodegradability
- optimal microbial condition, constant temperature, and ideal mixing, leading to complete anaerobic degradation of the organic matter content (no differentiation of biodegradable and non-biodegradable compounds);
- input substrate organic matter consists only of the elements C, H, O, N and S, and gaseous products resulting from anaerobic digestion are CH4, CO2, NH3, and H2S.
2.4. Experimental Biomethane Potential (BMP)
3. Results and Discussion
3.1. Salt Content of Halophyte Plant Material
3.2. Content of Carbohydrates, Lignin and Proteins
3.3. Elemental Composition of Organic Matter
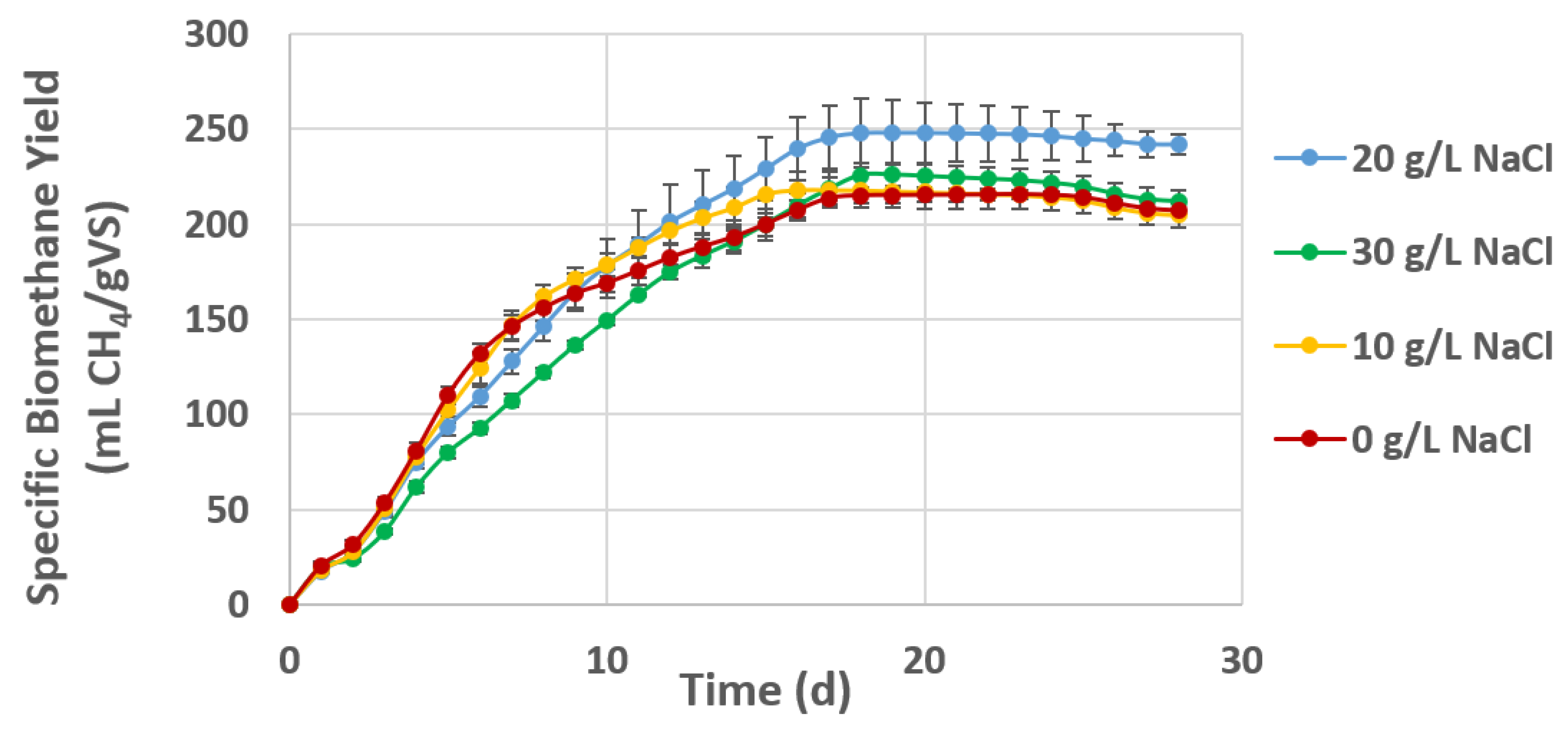
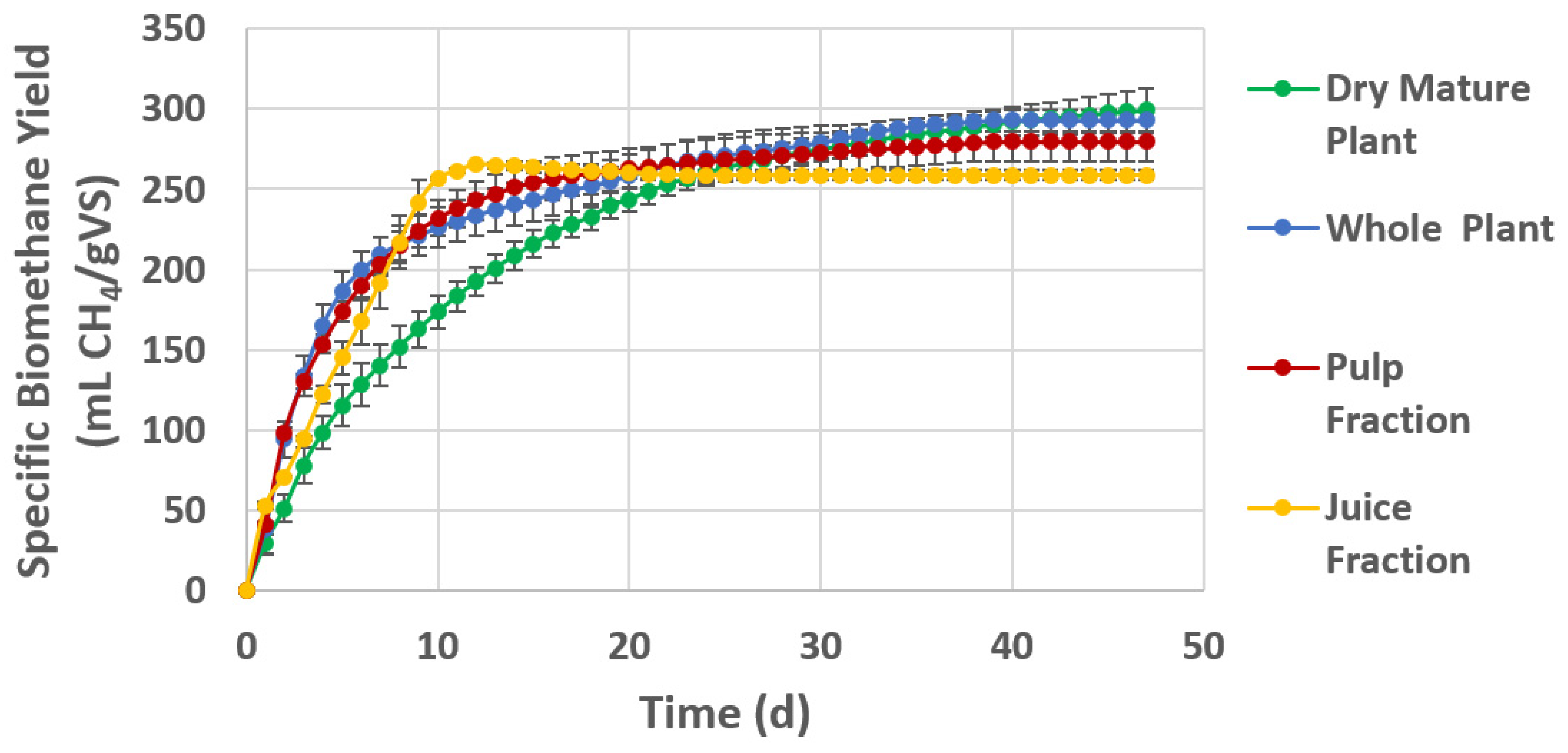
3.4. Biomethane Potential of Halophytic Substrates
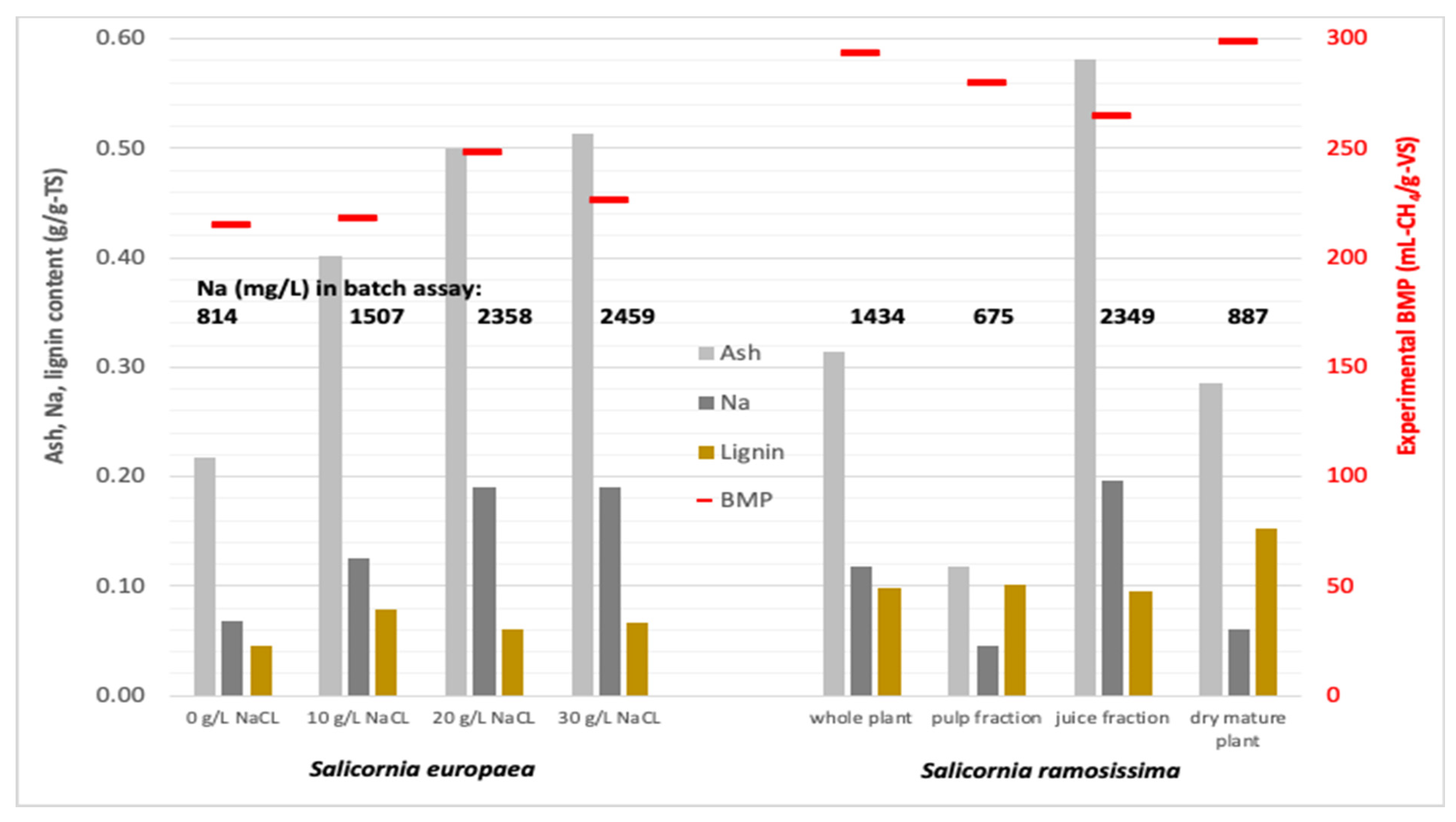
3.5. Mineral Concentration in Batch Tests
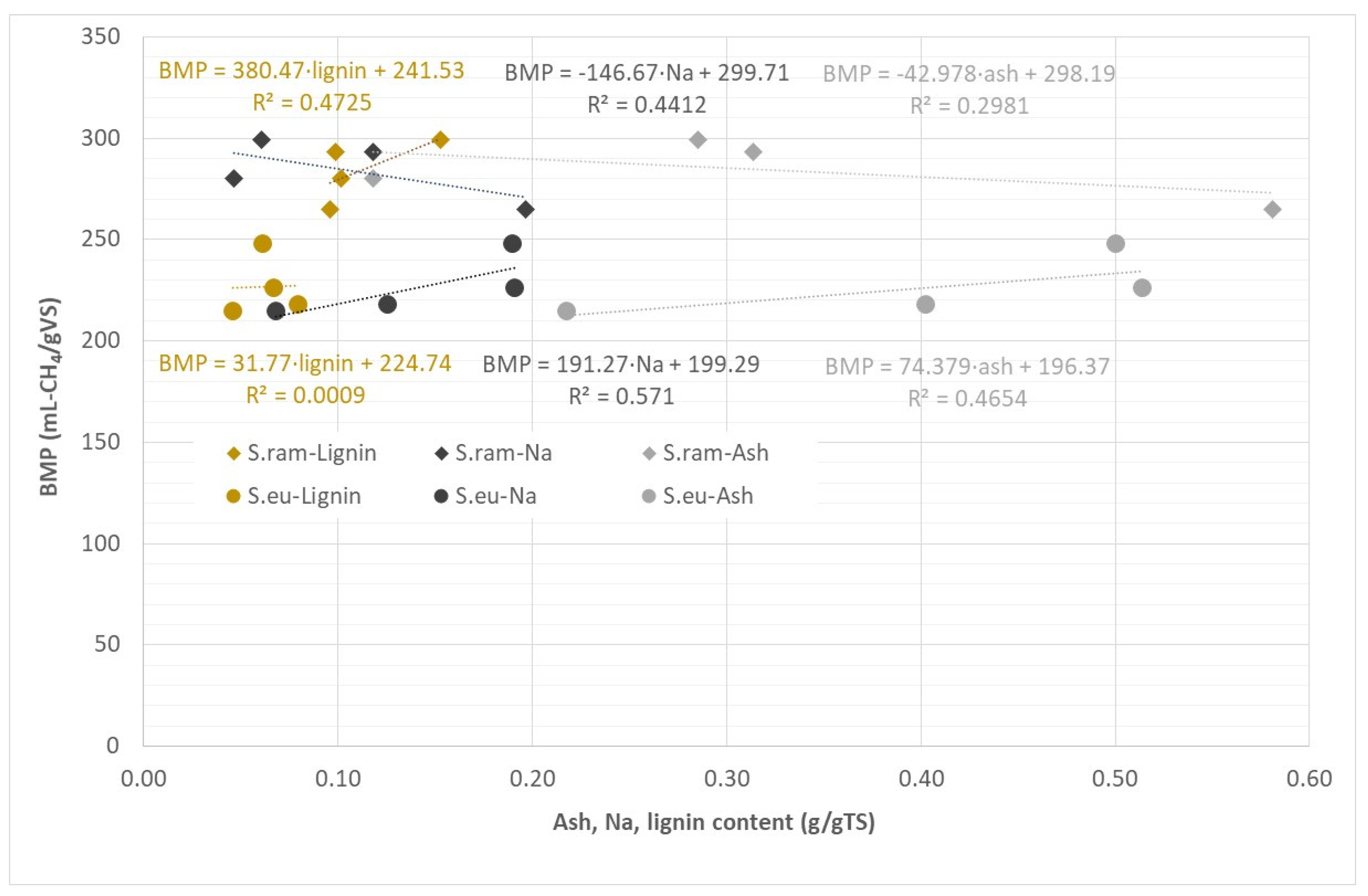
3.6. Correlation of Mineral and Lignin Content and BMP
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turcios, A.; Cayenne, A.; Uellendahl, H.; Papenbrock, J. Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Appl. Sci. 2021, 11, 2746. [Google Scholar] [CrossRef]
- Kamel, M.; Hammad, S.; Khalaphallah, R.; Elazeem, M.A. Halophytes and Salt Tolerant Wild Plants as a Feedstock for Biogas Production. J. BioSci. Biotechnol. 2019, 8, 151–159. [Google Scholar]
- Abideen, Z.; Ansari, R.; Khan, M.A. Halophytes: Potential Source of Ligno-Cellulosic Biomass for Ethanol Production. Biomass Bioenergy 2011, 35, 1818–1822. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt Tolerance and Crop Potential of Halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Deublein, D.; Angelika, S. (Eds.) Biogas from Waste and Renewable Resources—An Introduction; 3. Nachdr.; Wiley-VCH-Verl: Weinheim, Germany, 2010; ISBN 9783527318414. [Google Scholar]
- Al Seadi, T.; Rutz, D.; Prassl, H.; Köttner, M.; Finsterwalder, T.; Volk, S.; Janssen, R. Biogas Handbook; University of Southern Denmark Esbjerg: Esbjerg, Denmark, 2008; ISBN 9788799296200. [Google Scholar]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Machmüller, A.; Hopfner-Sixt, K.; Bodiroza, V.; Hrbek, R.; Friedel, J.; Pötsch, E.; Wagentristl, H.; et al. Methane production through anaerobic digestion of various energy crops grown in sustainable crop rotations. Bioresour. Technol. 2007, 98, 3204–3212. [Google Scholar] [CrossRef]
- Murphy, J.; Braun, R.; Weiland, P.; Wellinger, A. Biogas from crop digestion. IEA Bioenergy Task 2011, 37, 1–23. [Google Scholar]
- Yang, S.; Li, J.; Zheng, Z.; Meng, Z. Characterization of Spartina alterniflora as Feedstock for Anaerobic Digestion. Biomass Bioenergy 2009, 33, 597–602. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- McCarty, P.L. Anaerobic Waste Treatment Fundamentals—Part Four—Process Design. Public Works 1964, 95, 95–99. [Google Scholar]
- Kugelman, I.J.; McCarty, P.L. Cation Toxicity and Stimulation in Anaerobic Waste Treatment. J. Water Pollut. Control Fed. 1965, 37, 97–116. [Google Scholar]
- Oleszek, M.; Kowalska, I.; Oleszek, W. Phytochemicals in bioenergy crops. Phytochem. Rev. 2019, 18, 893–927. [Google Scholar] [CrossRef] [Green Version]
- Li, H.L.; Zhang, X.M.; Deng, F.D.; Han, X.G.; Xiao, C.W.; Han, S.J.; Wang, Z.P. Microbial methane production is affected by secondary metabolites in the heartwood of living trees in upland forests. Trees 2020, 34, 243–254. [Google Scholar] [CrossRef]
- Cybulska, I.; Brudecki, G.; Alassali, A.; Alassali, M.; Brown, J.J. Phytochemical composition of some common coastal halophytes of the United Arab Emirates. Emir. J. Food Agric. 2014, 26, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Isca, V.M.S.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. An overview of Salicornia genus: The phytochemical and pharmacological profile. In Natural Products: Research Reviews; Gupta, V.K., Ed.; Daya Publishing House: New Delhi, India, 2014; Volume 2, pp. 145–176. [Google Scholar]
- Cybulska, I.; Chaturvedi, T.; Alassali, A.; Brudecki, G.P.; Brown, J.J.; Sgouridi, S.; Thomsen, M.H. Characterization of the chemical composition of the halophyte Salicornia bigelovii under cultivation. Energy Fuels 2014, 28, 3873–3883. [Google Scholar] [CrossRef]
- DIN EN 12880; Characterization of Sludges e Determination of Dry Residue and Water Content; German Version. Beuth Verlag: Berlin, Germany, 2000.
- DIN EN 12879; Characterization of Sludges e Determination of the Loss on Ignition of Dry Mass; German Version. Beuth Verlag: Berlin, Germany, 2000.
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass; National Renewable Laboratory (NREL): Golden, CO, USA, 2008; pp. 1–15. [Google Scholar]
- Raposo, F.; Fernández-Cegrí, V.; De la Rubia, M.A.; Borja, R.; Béline, F.; Cavinato, C.; Demirer, G.; Fernández, B.; Fernández-Polanco, M.; Frigon, J.C.; et al. Biochemical methane potential (BMP) of solid organic substrates: Evaluation of anaerobic biodegradability using data from an international interlaboratory study. J. Chem. Technol. Biotechnol. 2011, 86, 1088–1098. [Google Scholar] [CrossRef]
- Rincón, B.; Heaven, S.; Banks, C.J.; Zhang, Y. Anaerobic Digestion of Whole-Crop Winter Wheat Silage for Renewable Energy Production. Energy Fuels 2012, 26, 2357–2364. [Google Scholar] [CrossRef] [Green Version]
- Ugwu, S.; Enweremadu, C. Biodegradability and kinetic studies on biomethane production from okra (Abelmoschus esculentus) waste. S. Afr. J. Sci. 2019, 115, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Weese, A.; Pallmann, P.; Papenbrock, J.; Riemenschneider, A. Brassica napus L. cultivars show a broad variability in their morphology, physiology and metabolite levels in response to sulfur limitations and to pathogen attack. Front. Plant Sci. 2015, 6, 9. [Google Scholar] [CrossRef]
- Boyle, W.C. Energy recovery from sanitary landfills—A review. Micro Energy Conv. 1977, 119–138. [Google Scholar] [CrossRef]
- Achinas, S.; Euverink, G. Theoretical analysis of biogas potential prediction from agricultural waste. Resour. Effic. Technol. 2016, 2, 143–147. [Google Scholar] [CrossRef] [Green Version]
- VDI 4630; VDI Guideline: Fermentation of Organic Materials. Characterisation of the Substrate, Sampling, Collection of Material Data, Fermentation Tests. Beuth Verlag: Berlin, Germany, 2016.
- Akinshina, N.; Naka, D.; Toderich, K.; Azizov, A.; Yasui, H. Anaerobic degradation of halophyte biomass for biogas production. J. Arid Land Stud. 2012, 22, 227–230. [Google Scholar]
- Turcios, A.; Weichgrebe, D.; Papenbrock, J. Effect of salt and sodium concentration on the anaerobic methanisation of the halophyte Tripolium pannonicum. Biomass Bioenergy 2016, 87, 69–77. [Google Scholar] [CrossRef]
- Turcios, A.; Weichgrebe, D.; Papenbrock, J. Potential use of the facultative halophyte Chenopodium quinoa Willd. as substrate for biogas production cultivated with different concentrations of sodium chloride under hydroponic conditions. Bioresour. Technol. 2016, 203, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Cybulska, I.; Chaturvedi, T.; Brudecki, G.P.; Kádár, Z.; Meyer, A.S.; Baldwin, R.M.; Thomsen, M.H. Chemical characterization and hydrothermal pretreatment of Salicornia bigelovii straw for enhanced enzymatic hydrolysis and bioethanol potential. Bioresour. Technol. 2014, 153, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.; Colmer, T. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Santamaria-Fernandez, M.; Ytting, N.K.; Lübeck, M.; Uellendahl, H. Potential Nutrient Recovery in a Green Biorefinery for Production of Feed, Fuel and Fertilizer for Organic Farming. Waste Biomass Valoriz. 2019, 11, 5901–5911. [Google Scholar] [CrossRef]
- Brown, J.J.; Cybulska, I.; Chaturvedi, T.; Thomsen, M.H. Halophytes for the Production of Liquid Biofuels. In Sabkha Ecosystems: Volume IV: Cash Crop Halophyte and Biodiversity Conservation; Springer: Dordrecht, The Netherlands, 2014; pp. 67–72. [Google Scholar]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Zollitsch, W.; Mayer, K.; Gruber, L. Biogas production from maize and dairy cattle manure—Influence of biomass composition on the methane yield. Agric. Ecosyst. Environ. 2007, 118, 173–182. [Google Scholar] [CrossRef]
- Ward, A.J.; Lewis, D.M.; Green, F.B. Anaerobic Digestion of Algae Biomass: A Review. Algal Res. 2014, 5, 204–214. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, Z.; Yang, S.; Fang, C.; Zou, X.; Zhang, J. Improving conversion of Spartina alterniflora into biogas by co-digestion with cow feces. Fuel Process. Technol. 2010, 91, 1416–1421. [Google Scholar] [CrossRef]
- Jackson-Moss, C.A.; Duncan, J.R.; Cooper, D.R. The effect of calcium on anaerobic digestion. Biotechnol. Lett. 1989, 11, 219–224. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Halophyte Species | TS | VS | VS | Ash Content |
|---|---|---|---|---|
| wt% FM | wt% FM | wt% TS | wt% TS | |
| Salicornia europaea | ||||
| 0 g/L NaCl | 97.2 (0.02) | 76.1 (4.09) | 78.3 (4.2) | 21.7 (4.2) |
| 10 g/L NaCl | 98.2 (0.1) | 58.7 (1.46) | 59.8 (1.49) | 40.2 (1.49) |
| 20 g/L NaCl | 98.6 (0.14) | 49.3 (1.3) | 50.0 (1.36) | 50.0 (1.36) |
| 30 g/L NaCl | 98.7 (0.18) | 48.0 (2.97) | 48.6 (3.10) | 51.4 (3.1) |
| Salicornia ramosissima | ||||
| Whole (green) plant | 11.8 (0.55) | 8.1 (0.39) | 68.6 (0.31) | 31.4 (0.31) |
| Pulp fraction | 30.1 (1.45) | 26.6 (1.47) | 88.2 (0.65) | 11.8 (0.65) |
| Juice fraction | 7.1 (0.03) | 3.0 (0.03) | 41.9 (0.30) | 58.1 (0.30) |
| Dry mature plant | 69.6 (0.9) | 49.8 (0.94) | 71.5 (0.85) | 28.5 (0.85) |
| Halophyte Species | Na | K | Ca | Mg | P |
|---|---|---|---|---|---|
| mg/gTS | mg/gTS | mg/gTS | mg/gTS | mg/gTS | |
| Salicornia europaea | |||||
| 0 g/L NaCl | 68.0 (0.27) | 36.0 (0.28) | 36.0 (0.21) | 2.1 (0.01) | 4.7 (0.06) |
| 10 g/L NaCl | 126 (0.87) | 24.0 (0.03) | 23.1 (0.10) | 1.7 (0.01) | 4.6 (0.13) |
| 20 g/L NaCl | 190 (0.85) | 15.8 (0.07) | 5.3 (0.03) | 0.8 (0.01) | 3.1 (0.03) |
| 30 g/L NaCl | 191 (7.16) | 15.7 (0.17) | 4.5 (0.04) | 0.7 (0.01) | 2.9 (0.02) |
| Salicornia ramosissima | |||||
| Whole (green) plant | 118 (2.42) | 13.5 (0.11) | 6.7 (0.03) | 1.9 (0.01) | 4.4 (0.08) |
| Pulp fraction | 46.2 (0.93) | 5.0 (0.08) | 7.5 (0.04) | 0.9 (0.01) | 2.4 (0.04) |
| Juice fraction | 197 * | 22.7 * | 5.9 * | 3.0 * | 6.5 * |
| Dry mature plant | 60.5 (0.23) | 27.8 (0.05) | 3.8 (0.01) | 1.5 (0.01) | 4.8 (0.02) |
| Halophyte Species | Total Sugars | Klason Lignin | Crude Protein |
|---|---|---|---|
| g/100 gTS | g/100 gTS | g/100 gTS | |
| Salicornia europaea | |||
| 0 g/L NaCl | 9.8 (0.21) | 4.6 (0.53) | 26.9 |
| 10 g/L NaCl | 10.6 (0.54) | 7.9 (0.31) | 24.0 |
| 20 g/L NaCl | 12.9 (0.61) | 6.1 (0.51) | 18.2 |
| 30 g/L NaCl | 8.7 (0.43) | 6.7 (0.25) | 18.9 |
| Salicornia ramosissima | |||
| Whole (green) plant | 45.1 (5.89) | 9.9 (0.54) | 7.2 |
| Pulp fraction | 58.6 (3.89) | 10.2 (0.32) | 7.0 |
| Juice fraction | 14.1 (0.21) | 9.6 * | nd |
| Dry mature plant | 45.9 (2.78) | 15.3 (0.46) | 5.5 |
| Halophyte Species | C | H | N | O | S | C/N Ratio | Elemental Composition |
|---|---|---|---|---|---|---|---|
| wt% TS | (wt/wt) | CcHhNnSsOo | |||||
| Salicornia europaea | |||||||
| 0 g/L NaCl | 29.3 (0.8) | 4.3 (0.1) | 4.3 (0.03) | 33.1 (0.9) | 5.6 (0.1) | 6.8 | C14H25N2S1O1 |
| 10 g/L NaCl | 28.1 (0.4) | 3.8 (0.1) | 3.8 (0.1) | 18.0 (0.5) | 5.1 (0.1) | 7.3 | C15H24N2S1O7 |
| 20 g/L NaCl | 22.0 (1.4) | 2.9 (0.2) | 2.9 (0.1) | 21.9 (0.9) | 3.9 (0.1) | 7.6 | C15H24N2S1O11 |
| 30 g/L NaCl | 20.9 (0.3) | 2.7 (0.1) | 3.0 (0.04) | 20.1 (0.2) | 3.8 (0.3) | 6.9 | C15H23N2S1O11 |
| Salicornia ramosissima | |||||||
| Whole (green) plant | 34.5 (0.5) | 4.1 (0.1) | 1.2 (0.1) | 23.6 (0.6) | 4.8 (0.1) | 30.0 | C35H50N1S2O18 |
| Pulp fraction | 39.9 (0.2) | 5.5 (0.1) | 1.1 (0.02) | 34.3 (0.2) | 6.9 (0.1) | 35.6 | C42H69N1S3O27 |
| Dry mature plant | 36.5 (0.2) | 5.1 (0.03) | 0.9 (0.04) | 22.1 (0.4) | 6.5 (0.1) | 41.4 | C39H63N1S3O22 |
| Halophyte Species | BMP | TBMP | BMP | BDCHONS |
|---|---|---|---|---|
| mL CH4/gVS | mL CH4/gVS | m3 CH4/tFM | % | |
| Salicornia europaea | ||||
| 0 g/L NaCl | 215 (6.3) | 316 | 164 | 68 |
| 10 g/L NaCl | 218 (9.8) | 466 | 128 | 47 |
| 20 g/L NaCl | 248 (14.3) | 347 | 122 | 71 |
| 30 g/L NaCl | 226 (6.3) | 348 | 109 | 65 |
| Salicornia ramosissima | ||||
| Whole (green) plant | 293 (6.7) | 497 | 23.6 | 62 |
| Pulp fraction | 280 (12.2) | 442 | 74.4 | 63 |
| Juice fraction | 265 (8.3) | nd | 7.8 | nd |
| Dry mature plant | 299 (13.8) | 547 | 149 | 55 |
| Halophyte Species | Na | K | Ca | P | Mg |
|---|---|---|---|---|---|
| (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | |
| Salicornia europaea | |||||
| 0 g/L NaCl | 814 (34.41) | 282 (17.87) | 433 (18.24) | 87.7 (2.45) | 24.1 (1.07) |
| 10 g/L NaCl | 1508 (63.49) | 255 (12.04) | 398 (11.96) | 97.5 (2.41) | 25.2 (0.87) |
| 20 g/L NaCl | 2358 (36.74) | 200 (3.06) | 240 (1.42) | 88.0 (0.71) | 18.3 (0.18) |
| 30 g/L NaCl | 2458 (96.67) | 207 (7.94) | 233 (2.66) | 87.3 (1.58) | 17.2 (0.37) |
| Salicornia ramosissima | |||||
| Whole (green) plant | 1434 (12.44) | 158 (1.41) | 246 (1.09) | 95.4 (0.56) | 27.1 (0.21) |
| Pulp fraction | 675 (7.46) | 69.4 (0.80) | 238 (1.51) | 72.2 (0.47) | 15.8 (0.15) |
| Juice fraction | 2349 * | 267 * | 245 * | 126 * | 40.0 * |
| Dry mature plant | 887 (8.89) | 282 (3.80) | 218 (0.93) | 97.8 (0.76) | 23.1 (0.22) |
| Inoculum | |||||
| Inoculum control | 344 | 33.3 | 184 | 55.1 | 9.5 |
| Halophyte Material | TS | Na | Limit of Material Added to 1 T Non-Saline Substrate |
|---|---|---|---|
| % FM | g/kg | kg | |
| Salicornia europaea | |||
| 0 g/L NaCl | 97.2 | 68 | 32.2 |
| 10 g/L NaCl | 98.2 | 126 | 17.0 |
| 20 g/L NaCl | 98.6 | 190 | 11.1 |
| 30 g/L NaCl | 98.7 | 191 | 11.0 |
| Salicornia ramosissima | |||
| Whole (green) plant | 11.8 | 118 | 178.2 |
| Pulp fraction | 30.1 | 46 | 178.1 |
| Juice fraction | 7.1 | 197 * | 178.3 |
| Dry mature plant | 69.6 | 61 | 52.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cayenne, A.; Turcios, A.E.; Thomsen, M.H.; Rocha, R.M.; Papenbrock, J.; Uellendahl, H. Halophytes as Feedstock for Biogas Production: Composition Analysis and Biomethane Potential of Salicornia spp. Plant Material from Hydroponic and Seawater Irrigation Systems. Fermentation 2022, 8, 189. https://doi.org/10.3390/fermentation8040189
Cayenne A, Turcios AE, Thomsen MH, Rocha RM, Papenbrock J, Uellendahl H. Halophytes as Feedstock for Biogas Production: Composition Analysis and Biomethane Potential of Salicornia spp. Plant Material from Hydroponic and Seawater Irrigation Systems. Fermentation. 2022; 8(4):189. https://doi.org/10.3390/fermentation8040189
Chicago/Turabian StyleCayenne, Aadila, Ariel E. Turcios, Mette Hedegaard Thomsen, Rui Miranda Rocha, Jutta Papenbrock, and Hinrich Uellendahl. 2022. "Halophytes as Feedstock for Biogas Production: Composition Analysis and Biomethane Potential of Salicornia spp. Plant Material from Hydroponic and Seawater Irrigation Systems" Fermentation 8, no. 4: 189. https://doi.org/10.3390/fermentation8040189