
Algicidal Properties of Microbial Fermentation Products on Inhibiting the Growth of Harmful Dinoflagellate Species

Abstract
:1. Introduction
2. Experimental Methods
2.1. Cultivation of the Dinoflagellate Species
2.2. Microbial Culture and Fermentation Broth Preparation
2.3. Inhibition Experiment of Fermentation Liquid
2.4. Calculation Method of Algae Removal Rate
2.5. Crude Extraction of Algal Inhibitory Substances in Fermentation Liquid
2.6. Analysis of Molecular Weight of Algicidal Substances in Dialysis Bags
3. Isolation of Macroporous Resin from Algicidal Substances of Strain Ba3
3.1. Crude Extract of Algicidal Substance in Fermentation Broth of Strain Ba3
3.2. Selection of Macroporous Resin
3.3. Separation of Macroporous Lipid, an Algicidal Substance
3.4. LC-MS Analysis of Algal Inhibition Substances
4. Results
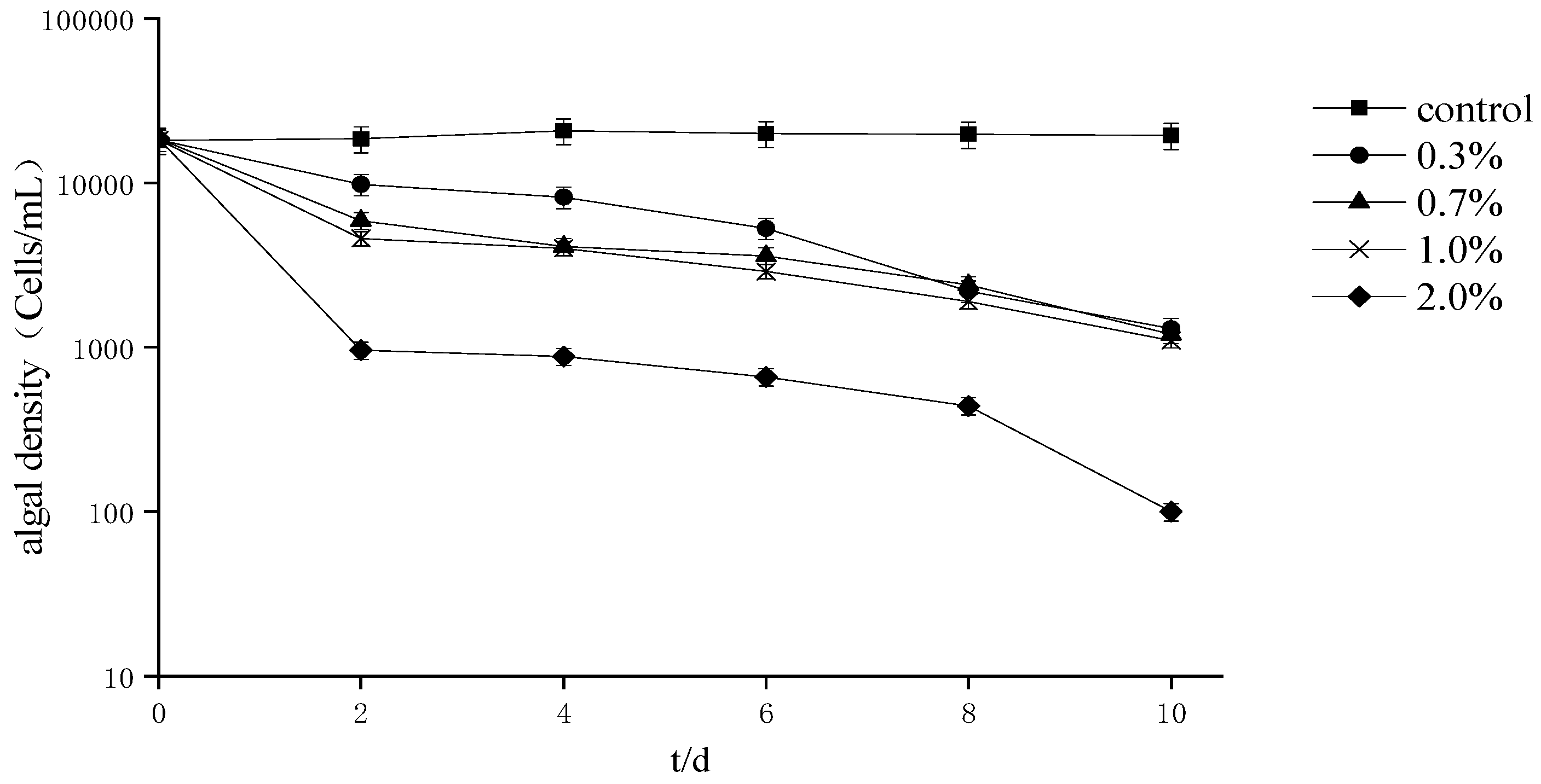
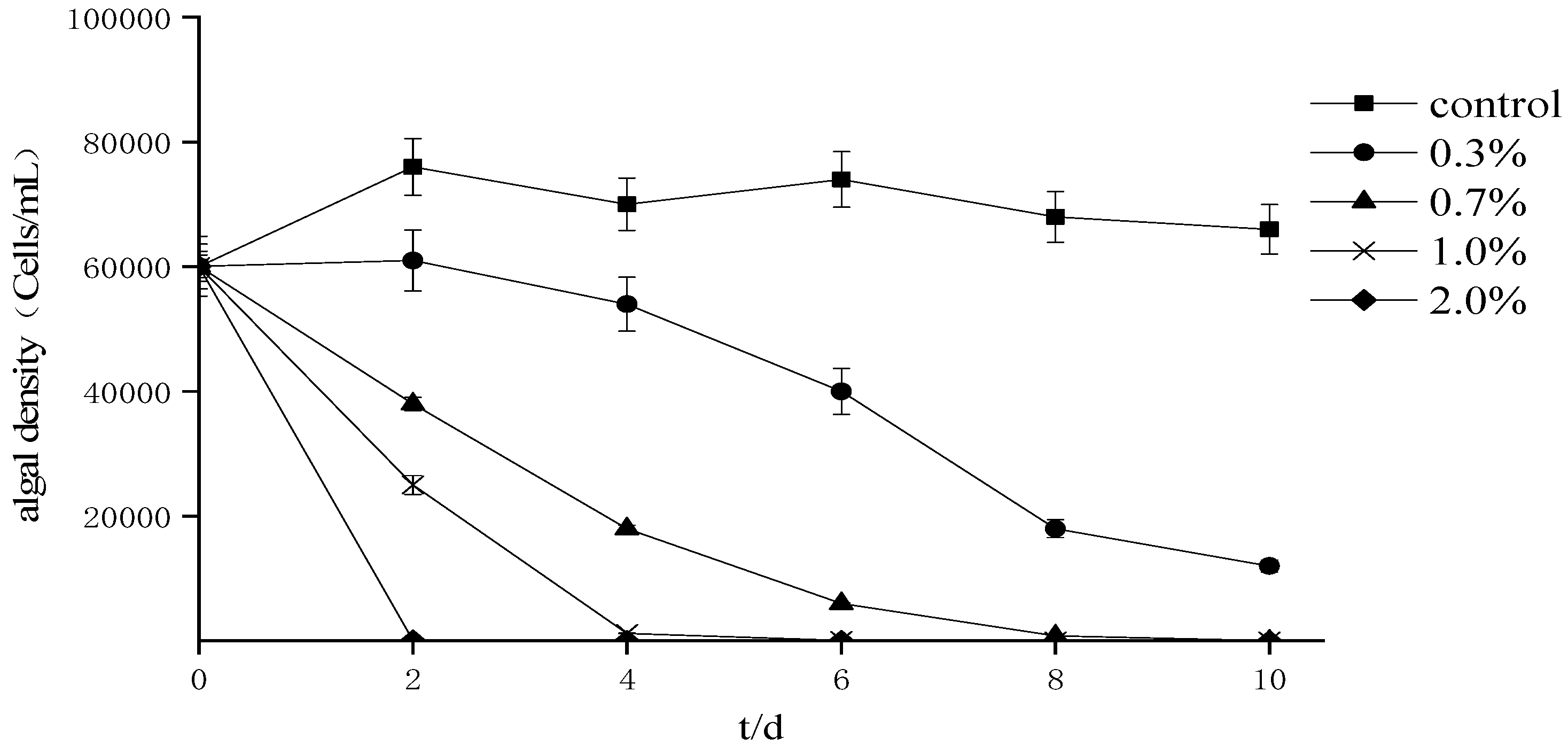
4.1. Effects of Ba3 Strain Fermentation Broth on the Growth of Two Kinds Red Tide Algae
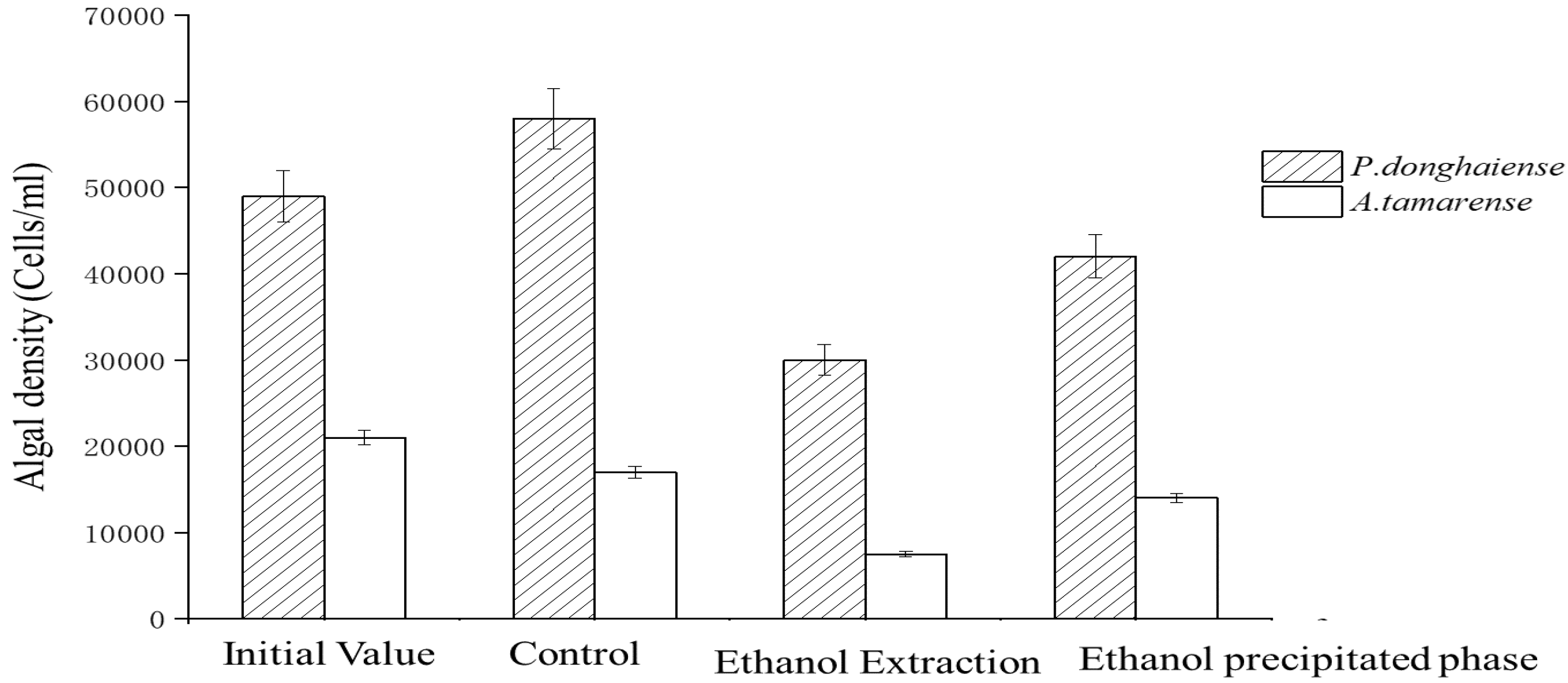
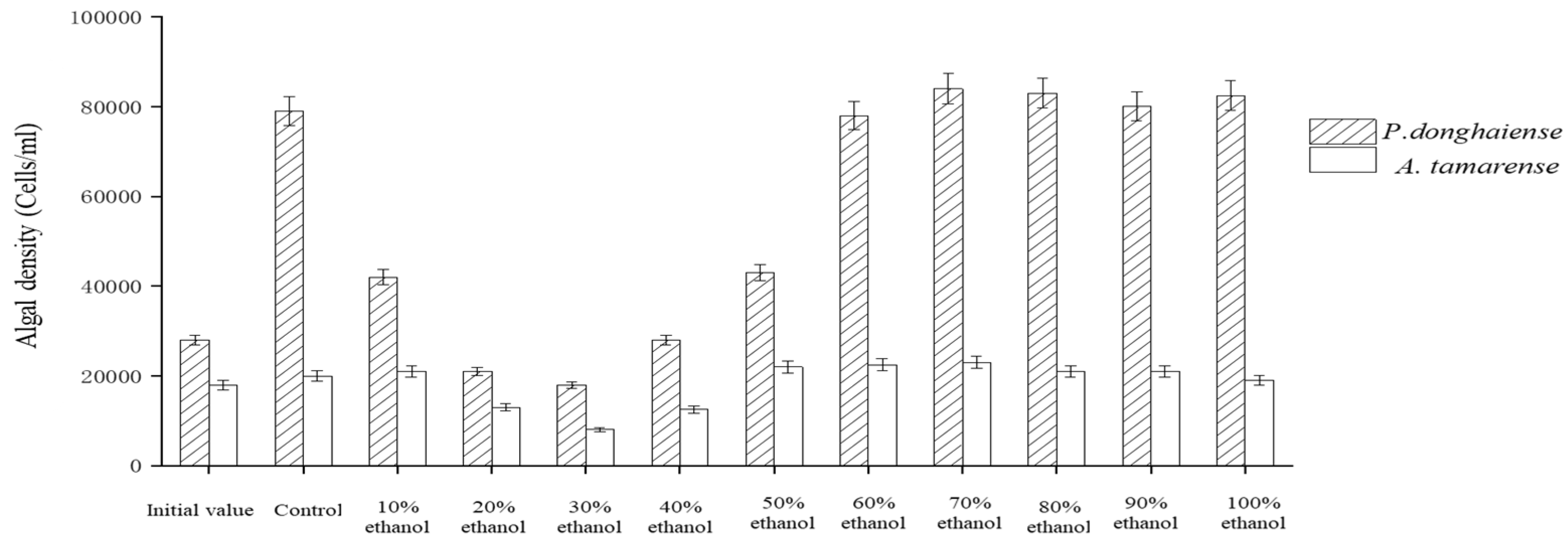
4.2. Effects of Ethanol Crude Extract of Fermentation Broth on the Growth of Red Tide Algae
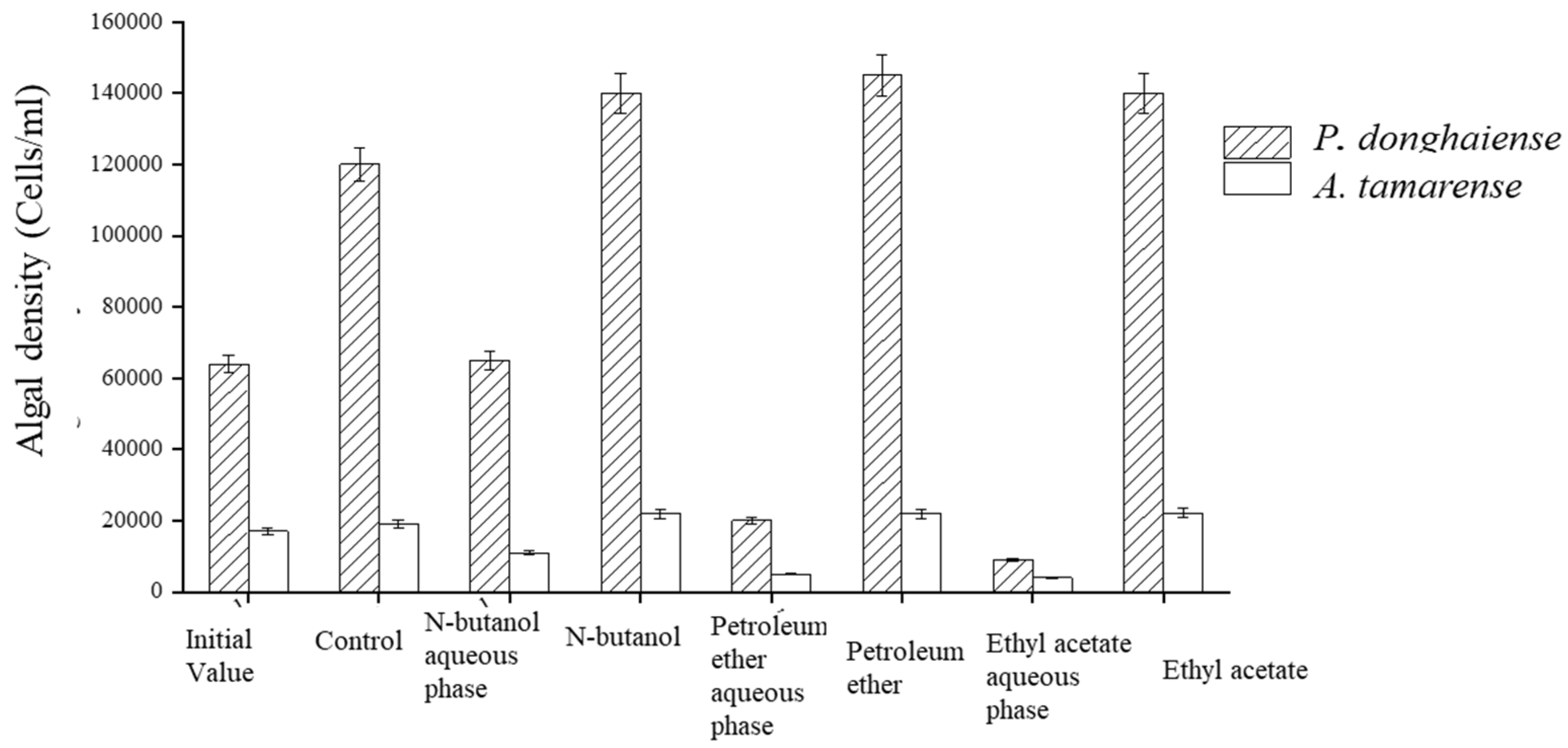
4.3. Effects of Different Organic Solvents Extracted from Fermentation Broth on the Growth of Red Tide Algae
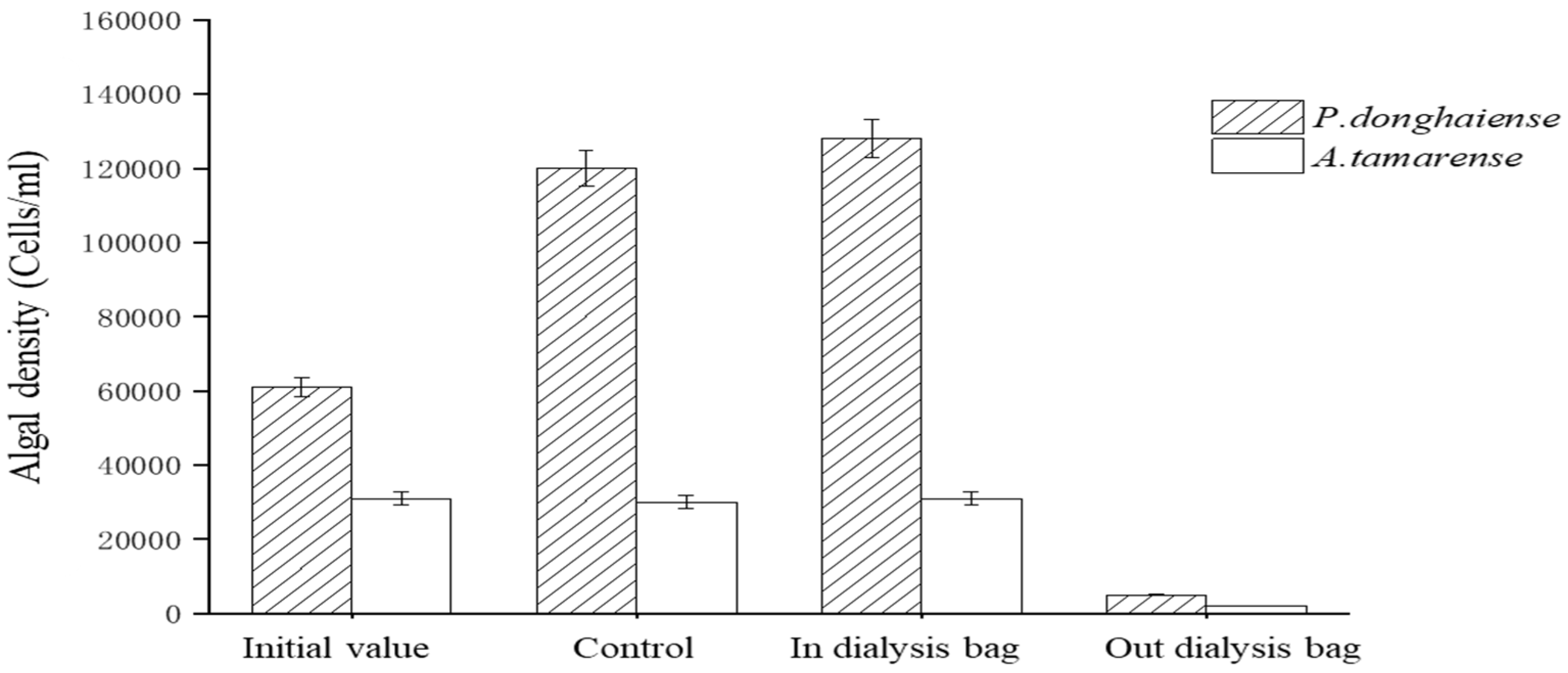
4.4. Molecular Weight Analysis of Dialysis Bags for Algal Inhibition Substances
4.5. Selection of Macroporous Resin
4.6. Inhibition Effect of Ethanol Elution of HZ830 Macroporous Resin
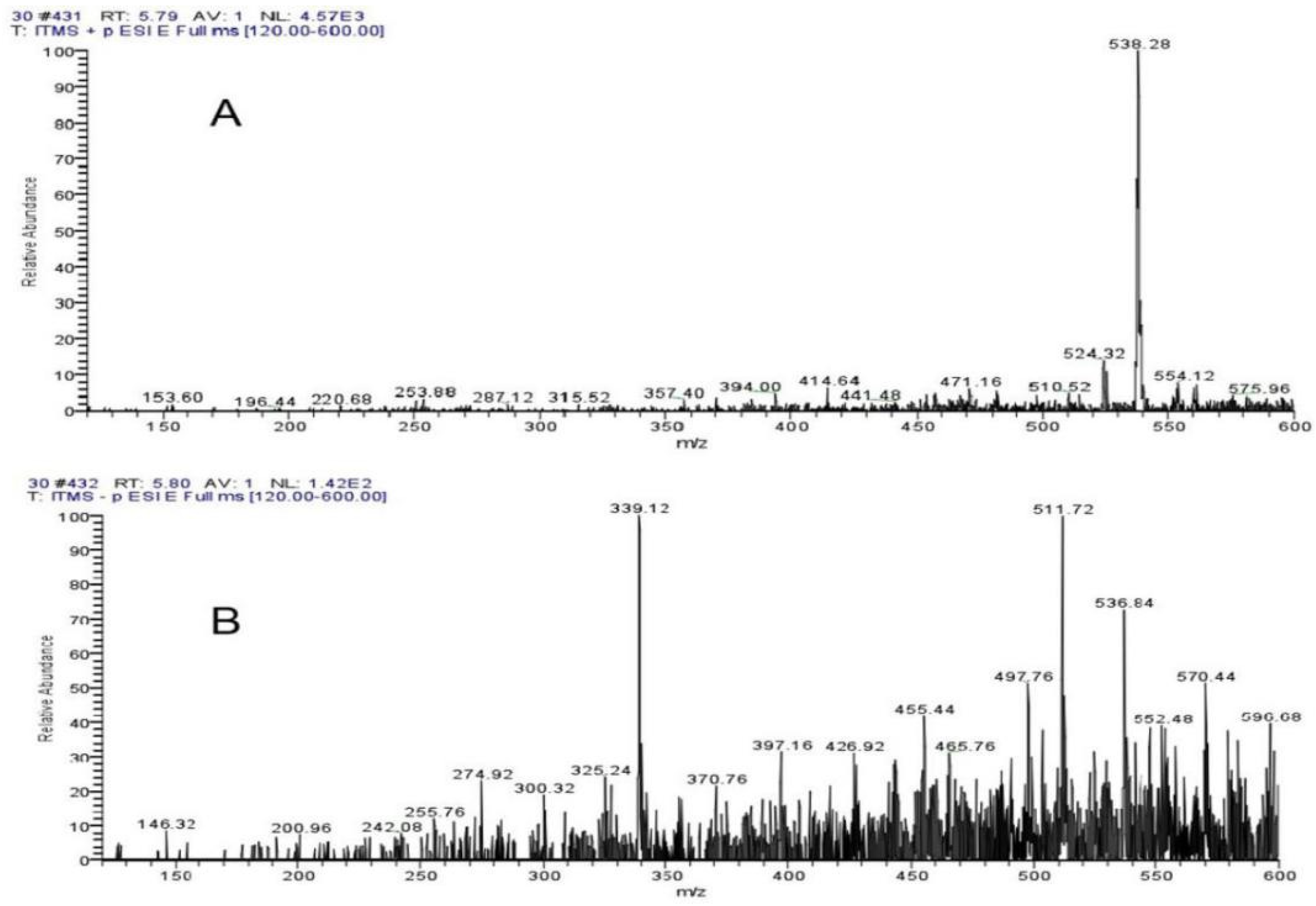
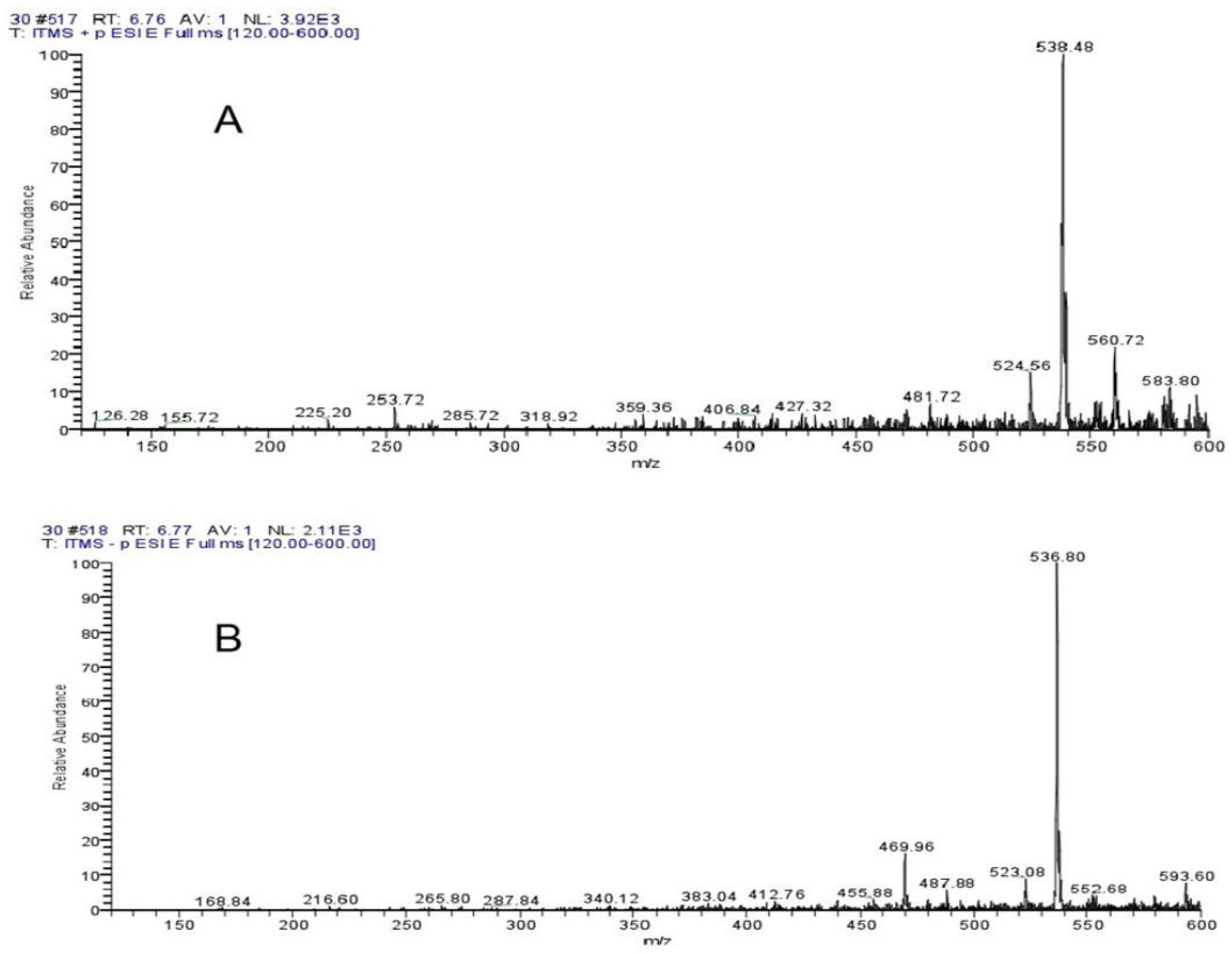
4.7. Liquid Mass Spectrometry Analysis of Algal Inhibition Substances
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2019, 91, 101590. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M.; Al-Azri, A.; Icarus Allen, J.; Bouwman, A.F.; Beusen, A.H.W.; Burford, M.A.; Harrison, P.J.; Zhou, M. Key questions and recent research advances on harmful algal blooms in relation to nutrients and eutrophication. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 229–259. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, T.; Zhou, J. Historical Occurrence of Algal Blooms in the Northern Beibu Gulf of China and Implications for Future Trends. Front. Microbiol. 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coast wide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.P.; Kudela, R.M.; Birch, J.M.; Blum, M.; Bowers, H.A.; Chavez, F.P.; Doucette, G.J.; Hayashi, K.; Marin, R.; Mikulski, C.M.; et al. Causality of an extreme harmful algal bloom in Monterey Bay, California, during the 2014–2016 northeast Pacific warm anomaly: Extreme HAB Causality in Monterey Bay. Geophys. Res. Lett. 2017, 44, 5571–5579. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Yang, C.Y.; Chi, Y.X.; Wu, D.H.; Dai, X.Z.; Zhang, X.H.; Igarashi, Y.; Luo, F. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake. J. Basic. Microb. 2019, 59, 1112–1124. [Google Scholar] [CrossRef]
- Bigalke, A.; Meyer, N.; Papanikolopoulou, L.A.; Wiltshire, K.H.; Pohnert, G. The algicidal bacterium Kordia algicida shapes a natural plankton community. Appl. Environ. Microbiol. 2019, 85, e02779-18. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Bigalke, A.; Kaulfuss, A.; Pohnert, G. Strategies and ecological roles of algicidal bacteria. FEMS Microbiol. Rev. 2017, 41, 880–899. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Chen, Q.; Zhang, D.; Zhang, H.; Lei, X.; Chen, Z.; Li, Y.; Hong, Y.; Ma, X.; Zheng, W.; et al. The algicidal mechanism of prodigiosin from Hahella sp. KA22 against Microcystis aeruginosa. Sci. Rep. 2017, 7, 7750. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Ye, Q.; Zhang, F.; Shao, X.; Fan, Y.; Zhu, X.; Li, Y.; Yao, L.; Tian, Y.; Zheng, T.; et al. Flocculating properties and potential of Halobacillus sp. strain H9 for the mitigation of Microcystis aeruginosa blooms. Chemosphere 2019, 218, 138–146. [Google Scholar] [CrossRef]
- Pokrzywinski, K.L.; Tilney, C.L.; Warner, M.E.; Coyne, K.J. Cell cycle arrest and biochemical changes accompanying cell death in harmful dinoflagellates following exposure to bacterial algicide IRI-160AA. Sci. Rep. 2017, 7, 45102. [Google Scholar] [CrossRef]
- Wang, M.; Chen, S.; Zhou, W.; Yuan, W.; Wang, D. Algal cell lysis by bacteria: A review and comparison to conventional methods. Algal Res. 2020, 46, 101794. [Google Scholar] [CrossRef]
- Ouyang, L.; Liu, Y.; Chen, H.; Zaynab, M.; Yang, X.; Wang, S.; Li, S. Encapsulation and Algicidal Properties of Fermentation Products From Vibrio brasiliensis H115. Front. Mar. Sci. 2021, 8, 676913. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, W.; Gu, X.; Guo, Z.; Song, J.; Zhu, D. Stabilizing lactate production through repeated batch fermentation of food waste and waste activated sludge. Bioresour. Technol. 2020, 300, 122709. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, Y.; Wang, Y.; Shi, F.; Nie, Y.; Liu, T. Effects of concentration variation on the physical properties of alginate-based substrates and cell behavior in culture. Int. J. Biol. Macromol. 2019, 128, 184–195. [Google Scholar] [CrossRef]
- Hu, X.; Su, H.; Xu, Y.; Xu, W.; Li, S.; Huang, X. Algicidal properties of fermentation products from Bacillus cereus strain JZBC1 dissolving dominant dinoflagellate species Scrippsiella trochoidea, Prorocentrum micans, and Peridinium umbonatum. Biologia 2020, 75, 2015–2024. [Google Scholar] [CrossRef]
- Balaji Prasath, B.; Wang, Y.; Su, Y.; Zheng, W.; Lin, H.; Yang, H. Coagulant Plus Bacillus nitratireducens Fermentation Broth Technique Provides a Rapid Algicidal Effect of Toxic Red Tide Dinoflagellate. J. Mar. Sci. Eng. 2021, 9, 395. [Google Scholar] [CrossRef]
- Lu, X.; Zhou, B.; Xu, L.; Liu, L.; Wang, G.; Liu, X. A marine algicidal Thalassospira and its active substance against the harmful algal bloom species Karenia mikimotoi. Appl. Microbiol. Biotechnol. 2016, 100, 5131–5139. [Google Scholar] [CrossRef]
- Sun, H.-Y.; Zhang, Y.; Chen, H.-R.; Hu, C.-X.; Li, H.; Hu, Z.-L. Isolation and characterization of the marine algicidal bacterium Pseudoalteromonas S1 against the harmful alga Akashiwo sanguinea. Mar. Biol. 2016, 163, 1–8. [Google Scholar] [CrossRef]
- Zhang, B.; Cai, G.; Wang, H.; Li, D.; Yang, X.; An, X. Streptomyces alboflavus RPS and its novel and high algicidal activity against harmful algal bloom species Phaeocystis globosa. PLoS ONE 2014, 9, e92907. [Google Scholar] [CrossRef]
- Bai, S.J.; Huang, L.P.; Su, J.Q.; Tian, Y.; Zheng, T.L. Algicidal effects of a novel marine actinomycete on the toxic dinoflagellate Alexandrium tamarense. Curr. Microbiol. 2011, 62, 1774–1781. [Google Scholar] [CrossRef]
- Cao, Y.C.; Wen, G.L.; Li, Z.J.; Hu, X.J.; Xi, J.Y.; Li, S.S.; Huang, H.H. Bacillus Cereus Strain JZBC1 Dissolving Dominant Dinoflagellate of Pond-Scrippsiella Trochoidea and Its Application. China Patent ZL201610209371.X, 5 February 2019. [Google Scholar]
- Guan, C.; Guo, X.; Cai, G.; Zhang, H.; Li, Y.; Zheng, W.; Zheng, T. Novel algicidal evidence of a bacterium Bacillus sp. LP-10 killing Phaeocystis globosa, a harmful algal bloom causing species. Biol. Control 2014, 76, 79–86. [Google Scholar] [CrossRef]
- Rani, A.S.; Jetty, A.; Ramakrishna, S.V. Kinetic studies of penicillin production during batch and repeated batch in fluidized bed bioreactor with agar immobilized P. chrysogenum cells. Indian J. Biotechnol. 2004, 3, 394–399. [Google Scholar]
- Meleigy, S.A.; Khalaf, M.A. Biosynthesis of gibberellic acid from milk permeate in repeated batch operation by a mutant Fusarium moniliforme cells immobilized on loofa sponge. Bioresour. Technol. 2009, 100, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Suvase, S.A.; Annapure, U.S.; Singhal, R.S. Gellan gum as an immobilization matrix for the production of cyclosporin A. J. Microbiol. Biotechnol. 2010, 20, 1086–1091. [Google Scholar] [CrossRef] [PubMed]
- Porcel, E.M.R.; López, J.L.C.; Pérez, J.A.S.; Chisti, Y. Lovastatin production by Aspergillus terreus in a two-staged feeding operation. J. Chem. Technol. Biotechnol. 2008, 83, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Perzborn, M.; Syldatk, C.; Rudat, J. Enzymatical and microbial degradation of cyclic dipeptides (diketopiperazines). AMB Express 2013, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Chen, L.; Yin, P. Algicidal metabolites produced by Bacillus sp strain B1 against Phaeocystis globosa. J. Ind. Microbiol. Biotechnol. 2014, 41, 593–599. [Google Scholar] [CrossRef]
- Ma, F.; Sun, Y.; Li, A.; Zhang, X.; Yang, J. Activation of accumulated nitrite reduction by immobilized Pseudomonas stutzeri T13 during aerobic denitrification. Bioresour. Technol. 2015, 187, 30–36. [Google Scholar] [CrossRef]
- Wang, J.; Hong, G.; Zhang, J. Study on the algal-lysis mechanism of a Microcystis aeruginosa alginolytic bacteria. J. Anhui Univ. Archit. Technol. Nat. Sci. Ed. 2017, 25, 19–23. (In Chinese) [Google Scholar]
- Yu, F. Isolation and Identification of Two High-Efficiency Actinomycetes of Heteroflexus Erythrolytica and Preliminary Research on Alginolytic Active Substances. Master’s Thesis, Central China Normal University, Shanghai, China, 2011. (In Chinese). [Google Scholar]
- Wang, J.X.; De, X.H.; Liu, J. Isolation and characterization of extracellular algic substances from algae-soluble bacteria. J. Inn. Mong. Agric. Univ. (Nat. Sci. Ed.) 2014, 35, 67–71. (In Chinese) [Google Scholar]
- Chen, M. Isolation and Identification of Extracellular Active Substances from Algae-Soluble Bacteria Purified by Glucose Gel. Master’s Thesis, Jinan University, Jinan, China, 2013. (In Chinese). [Google Scholar]
- Li, Y.; Pan, W.; Yang, L. Study on the separation characteristics of extracellular alginolytic active substances from three algicidal bacteria. Bull. Microbiol. 2008, 35, 7. (In Chinese) [Google Scholar]
- Tang, S.; Lin, W.; Li, J. Isolation of algae-lysing substances from Actinomyces L74 and their algae-lytic properties. Bull. Microbiol. 2011, 38, 6. (In Chinese) [Google Scholar]
- Wang, S.; Zou, R.; Wu, F. HPLC–MS/MS Analysis and Study on the Adsorption/Desorption Characteristics of Ginsenosides on Anion-Exchange Macroporous Resins. Chromatographia 2021, 84, 441–453. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Hu, G.; Huang, J.; Chen, L.; Yin, Y.; Cai, Y.; Shen, X.; Ji, N. Isolation and characterization of an algicidal bacterium against the bloom-forming algae raphidophyte Heterosigma akashiwo. Environ. Technol. 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Wen, G.L.; Xu, W.J.; Xu, Y.; Su, H.C.; Yang, K.; Xu, Y.N.; Li, Z.J.; Cao, Y.C. Effects of the algicidal bacterium CZBC1 on microalgal and bacterial communities in shrimp culture. Aquac. Environ. Interact. 2019, 11, 279–290. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Add the Amount | P. Donghaiense | A. Tamarense | ||
|---|---|---|---|---|---|
| Time | Pearson correlation Sig. (double tail) | 1 | 0.000 1.000 | −0.516 ** 0.000 | −0.164 0.122 |
| Add the amount | Pearson correlation Sig. (double tail) | 0.000 1.000 | 1 | −0.672 ** 0.000 | −0.634 ** 0.000 |
| P. donghaiense | Pearson correlation Sig. (double tail) | −0.516 ** 0.000 | −0.672 ** 0.000 | 1 | 0.814 ** 0.000 |
| A. tamarense | Pearson correlation Sig. (double tail) | −0.164 0.122 | 0.634 ** 0.000 | 0.814 ** 0.000 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balaji-Prasath, B.; Wang, Y.; Su, Y.; Chen, M.; Zheng, Y. Algicidal Properties of Microbial Fermentation Products on Inhibiting the Growth of Harmful Dinoflagellate Species. Fermentation 2022, 8, 176. https://doi.org/10.3390/fermentation8040176
Balaji-Prasath B, Wang Y, Su Y, Chen M, Zheng Y. Algicidal Properties of Microbial Fermentation Products on Inhibiting the Growth of Harmful Dinoflagellate Species. Fermentation. 2022; 8(4):176. https://doi.org/10.3390/fermentation8040176
Chicago/Turabian StyleBalaji-Prasath, Barathan, Ying Wang, Yuping Su, Minghua Chen, and Yi Zheng. 2022. "Algicidal Properties of Microbial Fermentation Products on Inhibiting the Growth of Harmful Dinoflagellate Species" Fermentation 8, no. 4: 176. https://doi.org/10.3390/fermentation8040176