
Isolation and Characterization of a Cholesterol-Lowering Bacteria from Bubalus bubalis Raw Milk

 , ,
, ,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Sampling
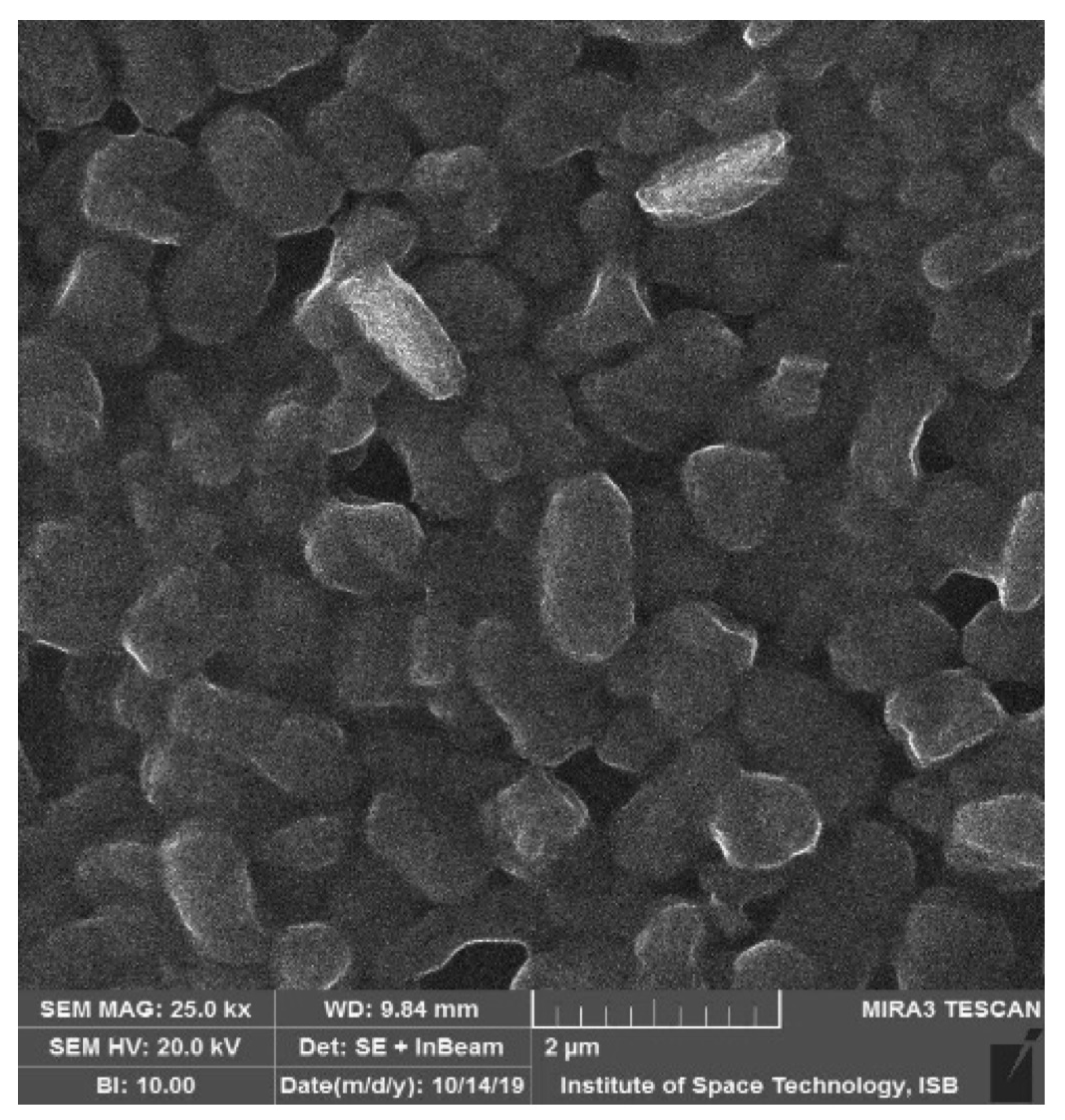
2.2. Bacterial Isolation and Phenotypic Characterization
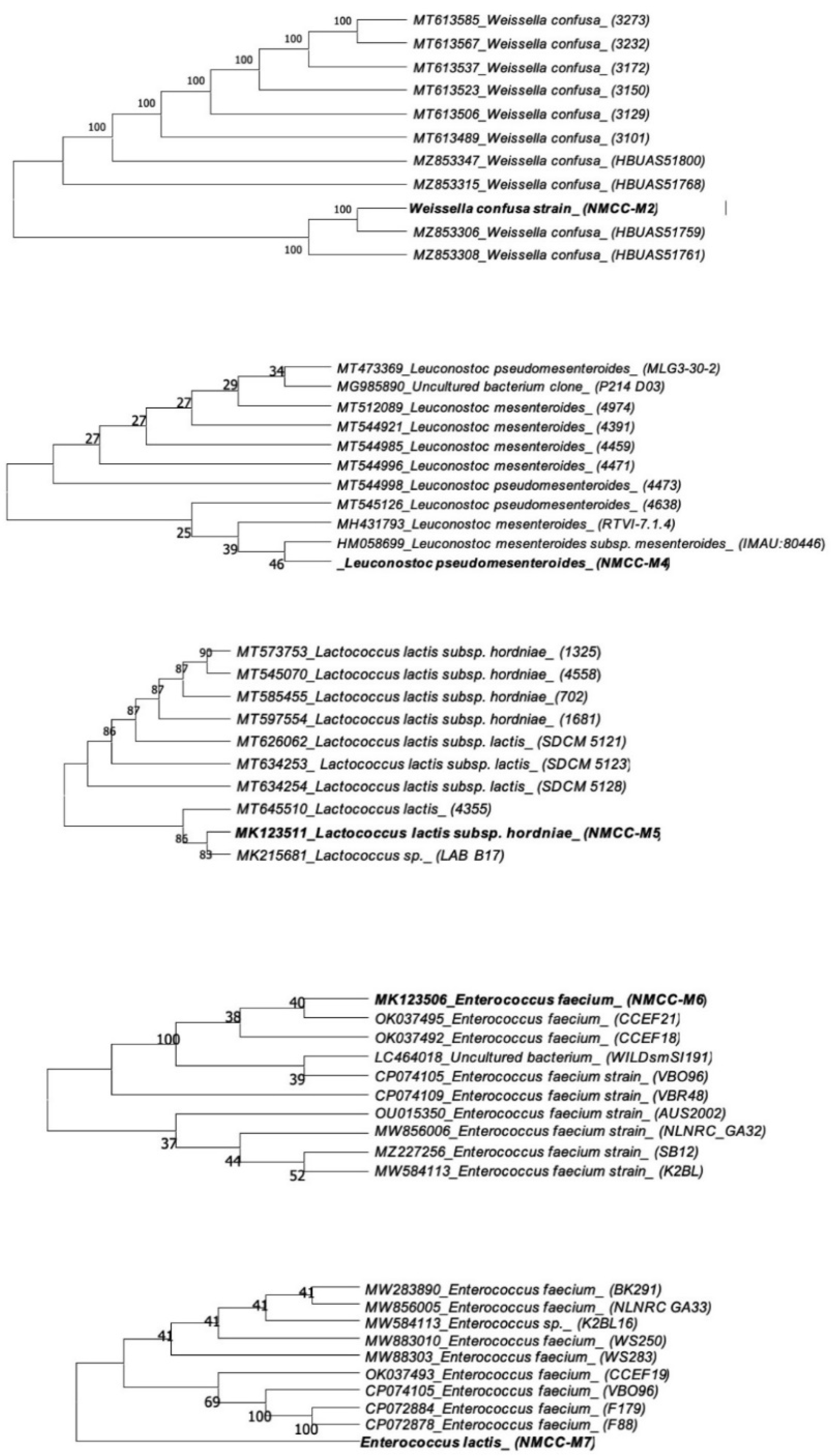
2.3. Bacterial Identification Based upon 16S rDNA Sequencing
2.4. Phylogenetic Analysis
2.5. Probiotic Characterization of Putative Probiotic Strains
2.5.1. Tolerance to Acid and Bile Salts
2.5.2. Determination of Antimicrobial Potential
2.5.3. Antibiotic Resistance
2.5.4. Screening of Proteolytic Activity
2.5.5. Screening of Lipolytic Activity
2.5.6. Screening of Amylolytic Activity
2.5.7. Screening of Hemolytic Activity
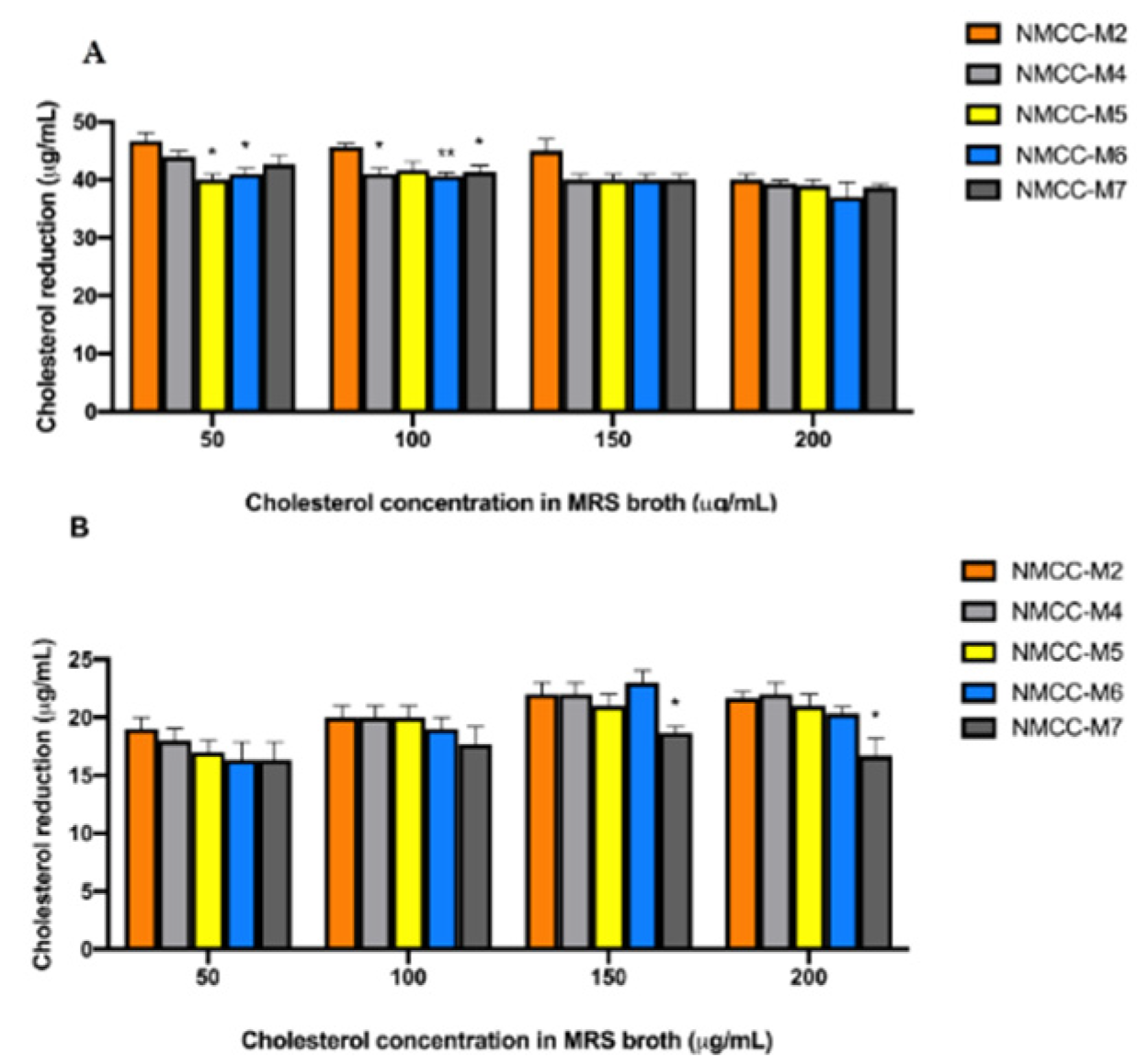
2.5.8. Cholesterol Reduction Assay
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Division, N. Health and Nutritional Properties and Guidelines for Evaluation—Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. FAO/WHO. 2006, p. 56. Available online: https://www.fao.org/publications/card/en/c/7c102d95-2fd5-5b22-8faf-f0b2e68dfbb6/ (accessed on 3 March 2022).
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and therapeutic effectiveness of lactobacilli. J. Clin. Pathol. 2016, 69, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Di Cerbo, A.; Canello, S.; Guidetti, G.; Laurino, C.; Palmieri, B. Unusual antibiotic presence in gym trained subjects with food intolerance; a case report. Nutr. Hosp. 2014, 30, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Anandharaj, M.; Sivasankari, B.; Santhanakaruppu, R.; Manimaran, M.; Rani, R.P.; Sivakumar, S. Determining the probiotic potential of cholesterol-reducing Lactobacillus and Weissella strains isolated from gherkins (fermented cucumber) and south Indian fermented koozh. Res. Microbiol. 2015, 166, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Di Cerbo, A.; Pezzuto, F.; Palmieri, L.; Rottigni, V.; Iannitti, T.; Palmieri, B. Clinical and experimental use of probiotic formulations for management of end-stage renal disease: An update. Int. Urol. Nephrol. 2013, 45, 1569–1576. [Google Scholar] [CrossRef]
- Blaiotta, G.; Murru, N.; Di Cerbo, A.; Succi, M.; Coppola, R.; Aponte, M. Commercially standardized process for probiotic “Italico” cheese production. LWT-Food Sci. Technol. 2017, 79, 601–608. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Pezzuto, F.; Palmieri, L.; Palmieri, B. The use of probiotics in the end-stage renal disease management. Minerva Biotecnol. 2012, 24, 155–170. [Google Scholar]
- Di Cerbo, A.; Palmieri, B. Lactobacillus Paracasei subsp. Paracasei F19; a farmacogenomic and clinical update. Nutr. Hosp. 2013, 28, 1842–1850. [Google Scholar] [CrossRef]
- Romano, A.; Blaiotta, G.; Di Cerbo, A.; Coppola, R.; Masi, P.; Aponte, M. Spray-dried chestnut extract containing Lactobacillus rhamnosus cells as novel ingredient for a probiotic chestnut mousse. J. Appl. Microbiol. 2014, 116, 1632–1641. [Google Scholar] [CrossRef] [Green Version]
- Blaiotta, G.; Murru, N.; Di Cerbo, A.; Romano, R.; Aponte, M. Production of probiotic bovine salami using Lactobacillus plantarum 299v as adjunct. J. Sci. Food Agric. 2018, 98, 2285–2294. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Barkat, R.A.; Gabr, A.A.; Foda, M.A.; Noreldin, A.E.; Khafaga, A.F.; El-Sabrout, K.; et al. Potential role of important nutraceuticals in poultry performance and health—A comprehensive review. Res. Vet. Sci. 2021, 137, 9–29. [Google Scholar] [CrossRef]
- Kanwal, H.; Di Cerbo, A.; Zulfiqar, F.; Sabia, C.; Nawaz, A.; Siddiqui, F.M.; Aqeel, M.; Ghazanfar, S. Probiotic Characterization and Population Diversity Analysis of Gut-Associated Pediococcus acidilactici for Its Potential Use in the Dairy Industry. Appl. Sci. 2021, 11, 9586. [Google Scholar] [CrossRef]
- Mangiaterra, S.; Schmidt-Kuntzel, A.; Marker, L.; Di Cerbo, A.; Piccinini, R.; Guadagnini, D.; Turba, M.E.; Berardi, S.; Galosi, L.; Preziuso, S.; et al. Effect of a Probiotic Mixture in Captive Cheetahs (Acinonyx Jubatus) with Gastrointestinal Symptoms-A Pilot Study. Animals 2022, 12, 395. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.L.; Lammers, K.; Brigidi, P.; Vitali, B.; Rizzello, F.; Gionchetti, P.; Campieri, M.; Kamm, M.A.; Knight, S.C.; Stagg, A.J. Modulation of human dendritic cell phenotype and function by probiotic bacteria. Gut 2004, 53, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Perdigon, G. The probiotic bacterium Lactobacillus casei induces activation of the gut mucosal immune system through innate immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saulnier, D.M.; Gibson, G.R.; Kolida, S. In vitro effects of selected synbiotics on the human faecal microbiota composition. FEMS Microbiol. Ecol. 2008, 66, 516–527. [Google Scholar] [CrossRef]
- Vanderpool, C.; Yan, F.; Polk, D.B. Mechanisms of probiotic action: Implications for therapeutic applications in inflammatory bowel diseases. Inflamm. Bowel Dis. 2008, 14, 1585–1596. [Google Scholar] [CrossRef]
- Chiang, S.S.; Pan, T.M. Beneficial effects of Lactobacillus paracasei subsp. paracasei NTU 101 and its fermented products. Appl. Microbiol. Biotechnol. 2012, 93, 903–916. [Google Scholar] [CrossRef] [Green Version]
- Di Cerbo, A.; Palmieri, B. Review: The market of probiotics. Pak. J. Pharm. Sci. 2015, 28, 2199–2206. [Google Scholar]
- Ricciardi, A.; Blaiotta, G.; Di Cerbo, A.; Succi, M.; Aponte, M. Behaviour of lactic acid bacteria populations in Pecorino di Carmasciano cheese samples submitted to environmental conditions prevailing in the gastrointestinal tract: Evaluation by means of a polyphasic approach. Int. J. Food Microbiol. 2014, 179, 64–71. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Aponte, M.; Esposito, R.; Bondi, M.; Palmieri, B. Comparison of the effects of hyaluronidase and hyaluronic acid on probiotics growth. BMC Microbiol. 2013, 13, 243. [Google Scholar] [CrossRef] [Green Version]
- WHO. Cardiovascular Diseases. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 31 July 2020).
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef] [PubMed]
- WHO. Diet, nutrition and the prevention of chronic diseases. World Health Organ. Tech. Rep. Ser. 2003, 916, 1–149. [Google Scholar]
- Heo, W.; Lee, E.S.; Cho, H.T.; Kim, J.H.; Lee, J.H.; Yoon, S.M.; Kwon, H.T.; Yang, S.; Kim, Y.J. Lactobacillus plantarum LRCC 5273 isolated from Kimchi ameliorates diet-induced hypercholesterolemia in C57BL/6 mice. Biosci. Biotechnol. Biochem. 2018, 82, 1964–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.A.; Kim, J. Effect of Probiotics on Blood Lipid Concentrations: A Meta-Analysis of Randomized Controlled Trials. Medicine 2015, 94, e1714. [Google Scholar] [CrossRef]
- Yildiz, G.; Öztürk, M.; Aslım, B.; Aslim, B. Identification of Lactobacillus strains from breast-fed infant and investigation of their cholesterol-reducing effects. World J. Microbiol. Biotechnol. 2011, 27, 2397–2406. [Google Scholar] [CrossRef]
- Sanders, M.E. Summary of probiotic activities of Bifidobacterium lactis HN019. J. Clin. Gastroenterol. 2006, 40, 776–783. [Google Scholar] [CrossRef]
- Naeem, M.; Ahmed, I.; Ahmed, S.; Ahmed, Z.; Riaz, M.N.; Ghazanfar, S. Screening of cattle gut associated Bacillus strains for their potential use as animal probiotic. Indian J. Anim. Res. 2018, B-948, 1–6. [Google Scholar] [CrossRef]
- Lee, K.W.; Park, J.Y.; Jeong, H.R.; Heo, H.J.; Han, N.S.; Kim, J.H. Probiotic properties of Weissella strains isolated from human faeces. Anaerobe 2012, 18, 96–102. [Google Scholar] [CrossRef]
- Aristimuno Ficoseco, C.; Mansilla, F.I.; Maldonado, N.C.; Miranda, H.; Fatima Nader-Macias, M.E.; Vignolo, G.M. Safety and Growth Optimization of Lactic Acid Bacteria Isolated from Feedlot Cattle for Probiotic Formula Design. Front. Microbiol. 2018, 9, 2220. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019; Volume 39. [Google Scholar]
- Pailin, T.; Kang, D.H.; Schmidt, K.; Fung, D.Y. Detection of extracellular bound proteinase in EPS-producing lactic acid bacteria cultures on skim milk agar. Lett. Appl. Microbiol. 2001, 33, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Bernfeld, P. Amylase α and β. Methods Enzymol. 1955, 1, 149–158. [Google Scholar] [CrossRef]
- Gilliland, S.E.; Nelson, C.R.; Maxwell, C. Assimilation of cholesterol by Lactobacillus acidophilus. Appl. Environ. Microbiol. 1985, 49, 377–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakra, A.K.; Domdi, L.; Hanjon, G.; Tilwani, Y.M.; Arul, V. Some probiotic potential of Weissella confusa MD1 and Weissella cibaria MD2 isolated from fermented batter. LWT 2020, 125, 109261. [Google Scholar] [CrossRef]
- Vijayakumar, M.; Ilavenil, S.; Kim, D.H.; Arasu, M.V.; Priya, K.; Choi, K.C. In-vitro assessment of the probiotic potential of Lactobacillus plantarum KCC-24 isolated from Italian rye-grass (Lolium multiflorum) forage. Anaerobe 2015, 32, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kandasamy, S.; Kavitake, D.; Shetty, P.H. Probiotic characterization and antioxidant properties of Weissella confusa KR780676, isolated from an Indian fermented food. LWT 2018, 97, 53–60. [Google Scholar] [CrossRef]
- Yu, H.-S.; Jang, H.J.; Lee, N.-K.; Paik, H.-D. Evaluation of the probiotic characteristics and prophylactic potential of Weissella cibaria strains isolated from kimchi. LWT 2019, 112, 108229. [Google Scholar] [CrossRef]
- Kimoto, H.; Ohmomo, S.; Okamoto, T. Cholesterol removal from media by lactococci. J. Dairy Sci. 2002, 85, 3182–3188. [Google Scholar] [CrossRef]
- Nami, Y.; Haghshenas, B.; Haghshenas, M.; Abdullah, N.; Yari Khosroushahi, A. The Prophylactic Effect of Probiotic Enterococcus lactis IW5 against Different Human Cancer Cells. Front. Microbiol. 2015, 6, 1317. [Google Scholar] [CrossRef] [Green Version]
- Assamoi, A.A.; Krabi, E.R.; Ehon, A.F.; N’guessan, G.A.; Niamké, L.S.; Thonart, P. Isolation and screening of Weissella strains for their potential use as starter during attiéké production. Biotechnol. Agron. Soc. Environ. 2016, 20, 355–362. [Google Scholar] [CrossRef]
- Tulini, F.L.; Hymery, N.; Haertle, T.; Le Blay, G.; De Martinis, E.C. Screening for antimicrobial and proteolytic activities of lactic acid bacteria isolated from cow, buffalo and goat milk and cheeses marketed in the southeast region of Brazil. J. Dairy Res. 2016, 83, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; El-Sayed, A.M.; Yeganeh, S.; Dadar, M.; Giri, S.S. Effect of Potential Probiotic Lactococcus lactis Subsp. lactis on Growth Performance, Intestinal Microbiota, Digestive Enzyme Activities, and Disease Resistance of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2017, 9, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Dincer, E.; Kivanc, M. Lipolytic Activity of Lactic Acid Bacteria Isolated from Turkish Pastırma. Anadolu Univ. J. Sci. Technol. C-Life Sci. Biotechnol. 2018, 7, 12–19. [Google Scholar] [CrossRef]
- Ghazanfar, S. Understanding the Mechanism of Action of Indigenous Target Probiotic Yeast: Linking the Manipulation of Gut Microbiota and Performance in Animals. In Saccharomyces; IntechOpen: London, UK, 2021. [Google Scholar]
- Ghazanfar, S.; Riaz, A.; Ali, G.M.; Naveed, S.; Arif, I.; Irshad, S.; Riaz, N.; Manzoor, K.N. Common Methods to Understand and Develop Indigenous Probiotics Yeast for Ruminant. In Yeasts in Biotechnology; IntechOpen: London, UK, 2019. [Google Scholar]
- Ghazanfar, S.; Khalid, N.; Ahmed, I.; Imran, M. Probiotic yeast: Mode of action and its effects on ruminant nutrition. In Yeast—Industrial Applications; IntechOpen: London, UK, 2017; pp. 179–202. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Strain ID | Catalase | Oxidase | Gram Staining | Morphology | Sugar Fermentation | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arabi | Fru | Gala | Mel | Rib | Xyl | Aes | Sal | Ram | |||||
| NMCC-M2 | − | − | + | Cocci/bacilli | + | + | + | + | + | − | + | + | − |
| NMCC-M4 | − | − | + | Cocci | + | + | + | + | + | − | + | − | − |
| NMCC-M5 | − | − | + | Cocci | + | − | + | + | + | - | + | + | − |
| NMCC-M6 | − | − | + | Cocci | + | − | + | + | + | + | + | − | − |
| NMCC-M7 | − | − | + | Cocci | + | + | + | + | + | − | + | + | − |
| Strain ID | Survival Rate pH (1.0) | Survival Rate pH (2.0) | Survival Rate pH (3.0) | p Value |
|---|---|---|---|---|
| NMCC-M2 | 28.06 ± 0.19 a | 40.68 ± 1.38 b | 82.26 ± 2.27 c | p < 0.0001 |
| NMCC-M4 | 27.81 ± 1.92 a | 36.56 ± 2.05 b | 67.99 ± 1.88 c | p < 0.0001 |
| NMCC-M5 | 24.69 ± 2.19 a | 32.80 ± 1.02 b | 68.25 ± 3.67 c | p < 0.001; p < 0.0001 |
| NMCC-M6 | 24.13 ± 0.69 a | 35.51 ± 0.53 b | 71.72 ± 5.17 c | p < 0.0001 |
| NMCC-M7 | 23.92 ± 1.80 a | 33.37 ± 1.14 b | 71.70 ± 1.61 c | p < 0.0001 |
| Strain ID | 3 h | 5 h | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | 0.3% | 0.5% | p Value | Control a | 0.3% | 0.5% | p Value | |
| NMCC-M2 | 7.21 ± 0.01 | 6.44 ± 0.10 a | 3.24 ± 0.06 b | p < 0.001 | 7.37 ± 0.06 a | 5.29 ± 0.06 a | 1.61 ± 0.06 a | p < 0.001 |
| NMCC-M4 | 6.41 ± 0.04 | 5.42 ± 0.07 a | 2.33 ± 0.02 b | p < 0.001 | 6.54 ± 0.01 a | 4.48 ± 0.06 b | 1.22 ± 0.02 a | p < 0.001 |
| NMCC-M5 | 5.37 ± 0.06 | 4.55 ± 0.21 a | 1.54 ± 0.11 b | p < 0.001 | 6.20 ± 0.02 a | 4.40 ± 0.12 b | 1.11 ± 0.01 a | p < 0.001 |
| NMCC-M6 | 5.49 ± 0.06 | 5.04 ± 0.61 a | 1.26 ± 0.07 b | p < 0.05; p < 0.001 | 5.58 ± 0.13 a | 3.47 ± 0.06 c | 1.03 ± 0.02 a | p < 0.001 |
| NMCC-M7 | 4.31 ± 0.11 | 3.30 ± 0.11 a | 1.37 ± 0.05 b | p < 0.001 | 5.36 ± 0.13 a | 2.47 ± 0.16 d | 4.42 ± 0.08 a | p < 0.001 |
| Strain ID | Salmonella Typhimurium | Bacillus cereus | Staphylococcus aureus | Escherichia coli | p Value |
|---|---|---|---|---|---|
| NMCC-M2 | 11.66 ± 0.57 | 10.12 ± 0.01 | 12.33 ± 0.87 | 12.36 ± 0.57 | |
| NMCC-M4 | 09.33 ± 0.57 | 09.33 ± 1.52 | 10.44 ± 1.15 | 10.16 ± 0.57 | |
| NMCC-M5 | 10.58 ± 1.15 | 08.68 ± 1.15 a | 11.12 ± 0.55 | 11.93 ± 1.15 b | p < 0.05 |
| NMCC-M6 | 10.19 ± 0.57 | 09.33 ± 2.3 | 10.33 ± 1.52 | 09.01 ± 1.73 | |
| NMCC-M7 | 10.61 ± 1.52 a | 06.01 ± 1.0 b,c,e | 09.66 ± 2.08 d | 09.11 ± 1.15 f | p < 0.001; p < 0.01; p < 0.05 |
| Strains ID | Amp a | Bac b | Chl c | Gen d | Kan e | Met f | Pen g | Hemolysis |
|---|---|---|---|---|---|---|---|---|
| NMCC-M2 | S | S | S | S | S | S | S | γ |
| NMCC-M4 | I | I | S | S | I | S c | S | γ |
| NMCC-M5 | I | I | I | S | S | S | I | γ |
| NMCC-M6 | S | I | I | S | S | I | S | γ |
| NMCC-M7 | S | S | S | S | S | S | I | γ |
| Strain ID | Lipolytic Activity | Amylolytic Activity | Proteolytic Activity |
|---|---|---|---|
| NMCC-M2 | ++ | ++ | ++ |
| NMCC-M4 | + | + | + |
| NMCC-M5 | ++ | - | + |
| NMCC-M6 | + | - | - |
| NMCC-M7 | + | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hameed, A.; Condò, C.; Tauseef, I.; Idrees, M.; Ghazanfar, S.; Farid, A.; Muzammal, M.; Al Mohaini, M.; Alsalman, A.J.; Al Hawaj, M.A.; et al. Isolation and Characterization of a Cholesterol-Lowering Bacteria from Bubalus bubalis Raw Milk. Fermentation 2022, 8, 163. https://doi.org/10.3390/fermentation8040163
Hameed A, Condò C, Tauseef I, Idrees M, Ghazanfar S, Farid A, Muzammal M, Al Mohaini M, Alsalman AJ, Al Hawaj MA, et al. Isolation and Characterization of a Cholesterol-Lowering Bacteria from Bubalus bubalis Raw Milk. Fermentation. 2022; 8(4):163. https://doi.org/10.3390/fermentation8040163
Chicago/Turabian StyleHameed, Abdul, Carla Condò, Isfahan Tauseef, Maryam Idrees, Shakira Ghazanfar, Arshad Farid, Muhammad Muzammal, Mohammed Al Mohaini, Abdulkhaliq J. Alsalman, Maitham A. Al Hawaj, and et al. 2022. "Isolation and Characterization of a Cholesterol-Lowering Bacteria from Bubalus bubalis Raw Milk" Fermentation 8, no. 4: 163. https://doi.org/10.3390/fermentation8040163