1. Introduction
Protein sources are the second-most important component in monogastric diets [
1]. Due to the continuous increase in price of soybean meal, nutritionists have been searching for alternative protein sources. Nevertheless, the use of alternative protein sources is limited to low inclusion due to the presence of anti-nutritional factors known to impair animal performance, nutrient digestibility, and feed utilization [
2]. A promising method that can be applied to improve the nutritive quality of these alternative protein sources is the fermentation process, resulting in digestibility improvement [
1,
2]. For optimal fermentation, carbohydrate rich feedstuffs such as cereal grains are also necessary as the glucose generated is a preferred substrate for fermentative microbes [
3]. Therefore, fermentation of alternative protein sources requires co-fermentation with grains, and hence can lead to overcoming negative properties through the action of fermentation itself (for example in rye: complex carbohydrates, phytic acid; sorghum: tannins, [
4]). As a whole, through fermentation, the usability and/or digestibility of proteins from alternative protein sources can be improved and the antinutritional factors of carbohydrate sources can be eliminated [
5,
6]. This should be taken into account not only because of the potential for better nutritional properties, but also because fermented feeds can have beneficial effects on animal health and well-being as well as food safety [
7].
Apart from improved nutritional properties, fermentation is often associated with a high concentration of organic acids and a low pH [
3,
8]. Van Winsen et al. [
9] described the desirable characteristics for fermented liquid feed as having a pH below 4.5, lactic acid concentrations above 150 mmol/L, and acetic acid and ethanol concentrations below 40 and 0.8 mmol/L, respectively. Thus, measuring different parameters of fermented substrates such as pH and concentration of acids are necessary to determine the efficiency of the fermentation process.
Different factors such as moisture, starter, and substrate, play a role in determining the fermentation characteristics as well as the nutrient profile of fermented substrates. The availability of water/moisture determines the fermentation quality, as it has an impact on microorganisms and their metabolic activity [
7]. Studies over the past years have strongly demonstrated that the moisture content of fermentation substrates should be adjusted according to many factors, such as substrate properties (granularity, hydraulics) [
10]. The starting culture, such as lactic acid bacteria (LAB), is considered another important factor to improve the fermentation process and act as the primary fermenting agent or aid in the growth and activity of other microorganisms required for the fermentation to take place [
11,
12]. Additionally, one of the most important factors affecting the quality/efficiency of fermentation is the type of carbohydrates, including starch and sugar, in the substrates. Starch in most cereals is the most abundant component within the grain, but the amount and composition vary depending upon grain type as well as variety [
13]. Starch is a complex branched glucose polymer that is composed of amylose and amylopectin [
14]. During fermentation, the glucose generated is a preferred substrate for fermentative microbes in feed, which may help to explain why the total carbohydrate content of feed decreases after fermentation [
12]. As a result, the protein supplements’ low starch and sugar content could be a concern which could be mitigated by utilizing cereals (co-fermentation), for example. Rye and sorghum, for example, are considered sustainable ingredients which may be good carbohydrate sources for fermentation and/or co-fermentation with alternative protein sources.
Rye is characterized by high tolerance to adverse environmental conditions (especially to low temperatures or drought) and high yield, even on less fertile soils of irregular pH [
15]. However, rye contains arabinoxylans, which are complex cell wall polysaccharides amounting to about 85 g/kg dry matter (DM) [
16]. It is well known that arabinoxylans enhance digesta viscosity [
17] due to a lack of endogenous enzymes that degrade dietary fiber, including soluble non-starch polysaccharides (NSP).
Sorghum is the fifth-most important cereal in the world, with high potential applications in feed production, such as replacement of corn in monogastric diets as well as high resistance to drought and limited water [
18]. The primary component of the sorghum kernel is starch that can amount to up to 80% of DM [
19]. Additionally, Knudsen [
20] found that the NSP content in the sorghum was about 54 g/kg DM.
Therefore, the objective of the present study was to improve some nutritional as well as physicochemical parameters of rye and sorghum (for example, as sustainable grains of different starch structures) by means of the fermentation process to be used as a co-fermentation for different alternative protein sources. Accordingly, in vitro experiments were carried out to elucidate the fermentation of rye or sorghum mixed with different ratios of water in terms of fermentation efficiency, especially the pH, acid concentration and viscosity. This could be an interesting perspective to be used as substrates and/or co-fermentation for some protein supplements.
2. Materials and Methods
2.1. Samples and Grinding
Two different cereal grains were investigated, including one genotype of hybrid rye (Trebiano, KWS LOCHOW GmbH, Bergen, Germany) and sorghum (Sorghum bicolor, Lupus, KWS LOCHOW GmbH, Bergen, Germany). The average DM contents of hybrid rye and sorghum were 880 g/kg and 860 g/kg, respectively. A rotor mill/ultra-centrifugal mill (Retsch ZM 200 mill, Retsch GmbH, Haan, Germany) with 1 or 10 mm sieve size was used for the comminuting the samples (cereal grains) to mimic the particle size distribution that was produced from a hammer mill as well as to have a maximal capacity procedure of grinding. Sieve sizes of 1 and 10 mm diameter produced coarse meal of varying fineness, which was then mixed and sampled with the aid of a sample divider (Tyler sample divider type 1, Haver & Boecker OHG, Oelde, Germany) for analysis.
2.2. Fermentation
The ground grains (rye or sorghum) of both 1 and 10 mm (mixed) were fermented in vitro by using a 300 mL sterile beaker. Briefly, the sample-to-water ratio was adjusted to achieve the planned DM contents (25%, 37.5%, 50%, 62.5%, and 75%) in the mixture before fermentation. To avoid malfermentation, a freeze-dried, granulated starter culture (Schaumalac Feed Protect XP G, H. Wilhelm Schaumann GmbH, Pinneberg, Germany), consisting of 1k2079
Lactobacillus plantarum, 1k2103
Pediococcus pentosaceus, and 1k2082
Lactococcus lactis was added at the beginning of each fermentation process at a dose of 2 × 10
7 CFU/g ingredient. The beakers were anaerobically (CO
2 = 10.0%, O
2 = 0.2%, N = 89.8% and temperature = 37.0 °C) incubated (Binder-Anaerobier Incubator, Fa. BINDER GmBH, Tuttlingen, Germany) for 6 h, 12 h, 24 h, and 48 h (
Figure 1).
A temperature during fermentation of 37 °C was ensured for the entire fermentation period. Samples were taken during and after fermentation at 0, 6, 12, 24, and 48 h for further analysis (
Figure 2).
2.3. Wet Sieve Analysis
Wet sieve analysis was carried out as described by Borgelt [
21] using the same sieves as for the dry sieve analysis. The sieves were dried at 103 °C until constant weight was achieved and then cooled to room temperature in a desiccator. The individual sieves were then weighed, thus completing the preparation of the sieve tower. For sample preparation, about 50 g of the sample to be analyzed was filled into a beaker. Then 800 mL of distilled water was added, and the sample was mixed vigorously for 10 s. After soaking for 1 h, stirring was repeated. The suspension was then added to the top sieve (largest mesh size) of the already prepared sieve tower. A further 10 L of distilled water was used to rinse the sieve tower. The wet sieve tower including the sample material was placed in the drying oven (model 600, Memmert GmbH & Co. KG, Schwabach, Germany) overnight. On the following day, the sieves were placed in the desiccator to cool down and could then be weighed again. The dry fractions on the individual sieves were calculated as a percentage of the total amount of weighed DM. The percentage of particles <200 µm included those particles that had been dissolved or washed out. Accordingly, this fine fraction can be calculated by subtracting the total mass of weighed DM and the sum of the DM mass in the individual sieves. The wet sieve analysis tests were repeated as previously mentioned, but with a soaking phase of 24 h instead of 1 h.
2.4. Dry Matter and Protein Analysis
The DM content was calculated by weighing the samples before and after drying them at 103 °C. The Dumas incineration method was also used to determine the total nitrogen content by using about 0.3 g of the sample in a crucible at 1000 °C in Elementar analyser (Vario Max CNS, Elementar Analysensysteme GmbH, Langenfeld, Germany). The crude protein can then be determined by multiplying the nitrogen content by the factor 6.25.
2.5. Lactic Acid Bacteria Counts
Samples (25 g) were obtained in stomacher bag from each grain with different DM contents containing Schaumalac Feed Protect XP G at three time points: before fermentation starting (zero time), after 24 h and 48 h of fermentation. In each sample, Buffered Peptone Water (BPW, ThermoFisher scientific GmbH, Germany) was added in a stomacher bag in a ratio of 1:10 and mixed using Bagmixer (BagMixer®400, Interscience SARL, Saint-Nom-la-Bretèche, France) for 90 s at speed level 3. This mixture was the first dilution step after homogenization of the suspension. Then, 1 mL of the suspension was transferred to a deep well block (96 well MegaBlock.RTM. 2.2 mL, Sarstedt AG & Co, Nümbrecht) and thereafter, 10-fold serial dilution using Phosphate Buffer Saline (PBS, ThermoFisher scientific GmbH, Germany) was performed. Three concentrations of 100 µL were streaked on Rogosa agar (RA, ThermoFisher scientific GmbH, Germany) in duplicate. The plates were anaerobically incubated (CO2 = 10.0%, O2 = 0.2%, N = 89.8%, Binder-Anaerobier Incubator, Fa. BINDER GmBH, Tuttlingen, Germany) at 37.0 °C for 24 h and 48 h. After incubation, the characteristic creamy white of LAB colonies were counted and expressed in a Log10 colony forming unit (CFU)/g.
2.6. pH Value
The pH values in the collected samples were determined directly. A certain amount of the sample (about 2 g) was diluted in a ratio of 1:5 with distilled water and was left to stand for 30 min at room temperature. The pH value was determined using a calibrated glass electrode (HI 2211 pH/ORP Meter, Hanna Instruments Deutschland GmbH, Vöhringen, Germany).
2.7. Lactic Acid
For determining lactate concentrations, perchloric acid (1 mol/L) was first added to the sample material and centrifuged. Potassium hydroxide solution was then added to 2 mL of the supernatant until a pH range of 8–10 was reached. After re-centrifugation, an enzymatic determination (D-/L-lactic acid UV test, Roche Diagnostics GmbH, Mannheim, Germany) of the L- and D-lactate content was carried out in accordance with Bunte et al. [
22].
2.8. Fatty Acid
The content of short-chain fatty acids (SCFA) in the homogenized sample was measured with the help of a gas chromatograph (610 Series, Unicam Chromatography GmbH & Co. KG, Kassel, Germany) in accordance with Bunte et al. [
22]. The sample was mixed with distilled water at a ratio of 1:4 and centrifuged. After adding an internal standard to the sample (preparation: 10 mL formic acid 89% and 0.1 mL 4-methylvaleric acid), the sample was centrifuged and then gas chromatography was applied. The column has a temperature program of maximum 155 °C (injector: 175 °C, detector: 180 °C). The fatty acids were separated within a running time of 30 min and then determined by means of a flame ionization detector.
2.9. Extract Viscosity
The method used to determine the extract viscosity was based on that described by Dusel et al. [
23] but with some modifications. About 5 g of fermented grain was added to 20 mL tap water and then shaken for 5 s on a vortex mixer (Heidolph Reax 2000, Fa.KaliChemie Pharma GmbH, Hannover, Germany). After a standing time of 30 min at 38 °C (incubator model 500, Memmert, GmbH & Co. KG, Schwabach, Germany), the samples were processed again using a vortex mixer and then centrifuged for 10 min at a force of 10,000 g (Heraeus Biofuge Stratos, Kendro Laboratory Products, Osterode, Germany). After centrifugation, the viscosity was determined using Model DVNext, DVNXLVCJG, Viscometer from Ametek Brookfield, Inc., Instrumentation & Specialty Controls Division, Middleborough, MA, USA. For this purpose, 600 µL were removed from the supernatant fluid in the centrifuge tubes and transferred to the measuring unit of the viscometer set to 26 °C. The measuring unit contained an S40 spindle that rotated at 10 rpm. After 1 min, the specified value was recorded.
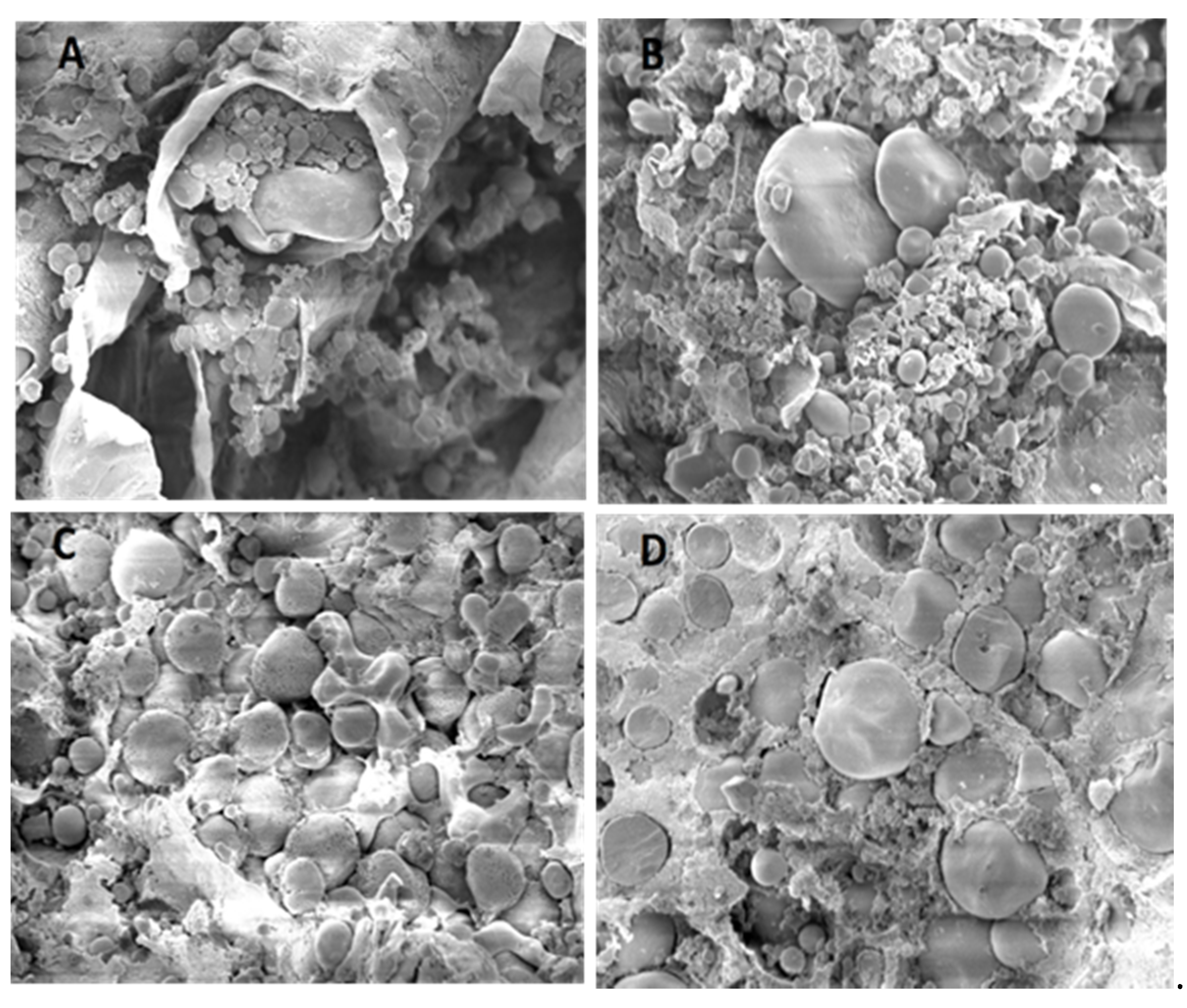
2.10. Scanning Electron Microscopy Analysis
For determining the microstructure of the unfermented and fermented rye and sorghum (of 37.5% DM), a scanned electron microscope was used (EVO15: Carl Zeiss Scanning Electron Microscope GmbH, Jena, Germany) at ×1000 magnification. The examination was done at the Institute for Pathology, University of Veterinary Medicine Hannover, Foundation, Hannover, Germany. Briefly, the samples were fixed in 5% buffered glutaraldehyde (Electron Microscopy Sciences), then rinsed several times in 0.1 M cacodylate buffer. Thereafter, the samples were treated in 1% osmium tetroxide (Electron Microscopy Sciences, Hatfield, PA, USA) for two hours and washed again in 0.1 M cacodylate buffer for one day. The samples were dehydrated in graded alcohol dilutions for two days, infiltrated in isoamyl acetate, and critically point dried using a Polaron E3000 critical point dryer (Quorum Design GmbH, Darmstadt, Germany). Samples were dried doubled at the critical point and coated with gold particles in a sputtering coater (SCD 040: Balzers Union Limited, Balzers, Liechtenstein).
2.11. Statistical Analysis
Statistical analysis was performed using the Statistical Analysis System for Windows, the SAS® Enterprise Guide®, version 9.3 (SAS Institute Inc., Cary, NC, USA). All parameters were analyzed for the individual samples (n = 4). Moreover, the mean values as well as the standard deviation of the mean (SD) were calculated. Assuming normally distributed data, a Ryan–Einot–Gabriel–Welsch test (simple Anova) was performed for checking significant differences of the data. Differences with a significant level of p < 0.05 were considered significant.
3. Results
3.1. Particle Size Distribution
The particle size distribution of the rye and sorghum is summarized in
Table 1. The average percentage of rye particles before fermentation (0 h) for experimental groups was about 29.3% DM and 44.9% DM at <0.20 mm and >1.00 mm, respectively. After 24 h of fermentation, significant differences in the particle size distribution <0.20 mm among the different DM contents of rye were noted. Whereas fermented rye with 37.5% DM had the greatest particle size distribution <0.20 mm (73.9%), fermented rye with 62.5% DM content had the lowest (46.4%). On the contrary, fermented rye with 75% DM content had the greatest particle size distribution >1.00 mm (49.8%), while fermented rye with 25% and 37.5% had the lowest percentages (14.3 and 17.0%, respectively).
The average percentage of sorghum particles before fermentation (0 h) for experimental groups was about 20.5% DM and 27.8% DM at <0.20 mm and >1.00 mm, respectively. After 24 h of fermentation, only significant differences were apparent in the particle size distribution <0.20 mm between fermented sorghum of DM contents of 62.5% and 75% (24.2 and 17.5%, respectively). Fermented sorghum with 50% and 75% DM contents had the greatest particle size distribution >1.00 mm (27.9 and 27.3%, respectively) compared to those with 37.5% and 62.5% DM contents (22.1 and 22.0%, respectively).
3.2. Protein Content
The protein contents of the rye and sorghum are presented in
Table 2. The protein content in the fermented rye after 24 h did not significantly differ between groups (range: 104–106 g/kg DM). Also, after 48 h of rye fermentation, no significant differences in protein content were seen among groups, except between fermented rye with 25% DM and 62.5% DM (106 and 104 g/kg DM, respectively).
After 24 h of sorghum fermentation, significant differences were only seen in protein content between fermented sorghum with DM contents of 25% and 50% (132 and 126 g/kg DM, respectively). No significant differences were observed in the protein content after 48 h of sorghum fermentation between groups (range: 129–132 g/kg DM).
3.3. Counting Lactic Acid Bacteria
The LAB counts of the rye and sorghum is summarized in
Table 3. After fermenting rye for either 24 h or 48, the LAB counts were significantly the lowest for fermented rye with 75% DM (7.15 and 6.32 log
10 CFU/g, respectively) compared to the other groups (
p < 0.0001). However, no significant differences were noted in the LAB counts for fermented rye with 25, 37.5, 50, and 62.5% DM contents after either 24 h or 48 h.
In the same trend, the LAB counts in the fermented sorghum with 75% DM for either 24 h or 48 h (5.93 and 6.38 log10 CFU/g, respectively) had significantly the lowest counts compared to the other groups (p < 0.0001). Moreover, no significant differences were noted in the LAB counts for fermented sorghum with 25, 37.5, 50, and 62.5% DM contents after either 24 h or 48 h.
3.4. pH Value
The pH values of fermented rye containing different DM contents are shown in
Table 4. After fermenting the rye for 6 h, the DM contents of 62.5% and 75% had the highest significant pH values (6.48 and 6.49, respectively), while the DM content of 25% was the lowest in pH value (6.13). Also, after fermenting the rye for 12 h, the DM contents of 62.5% and 75% had the highest significant pH values (6.27 and 6.49, respectively), while the DM content of 37.5% was the lowest in pH value (4.16). Generally, after 24 h or 48 h of rye fermentation, the DM content of 75% had the highest significant pH values (6.42 and 6.40, respectively), while the DM content of 25% was the lowest in pH value (3.57 and 3.55, after 24 and 48 h, respectively).
Statistical differences were noted in the pH values after 6 h of sorghum fermentation among all treatments, whereas the DM content of 75% was the greatest and the DM content of 25% was the lowest (6.54 vs 5.95). The same trend was observed in the pH values of fermented sorghum after 12, 24, and 48 h, whereas the DM content of 75% was the greatest (range: 6.27–6.51) and the DM content of 25% was the lowest (in addition to DM content of 37.5% after 48 h) with a range of 3.77–3.93.
3.5. L-lactic Acid Content
The content of L-lactic acid in fermented rye and sorghum after 24 h or 48 h is reported in
Table 5. The L-lactic acid concentration in the fermented rye after either 24 h or 48 h was significantly the greatest in the DM content of 25% (18.7 and 22.9 g/kg DM, respectively) compared to other treatments. While, the L-lactic acid concentration in the fermented rye after either 24 h or 48 h was significantly the lowest in the DM content of 75% (0.36 and 0.55 g/kg DM, respectively) compared to the other treatments.
In the case of fermented sorghum, the L-lactic acid concentration after either 24 h or 48 h was significantly the greatest in the DM content of 25% (22.2 g/kg and 24.4 g/kg DM, respectively) compared to other treatments. Whereas, the L-lactic acid concentration in the fermented sorghum after either 24 h or 48 h was significantly the lowest in the DM content of 75% (0.13 and 0.19 g/kg DM, respectively) compared to the other treatments.
3.6. Fatty Acid Concentrations
The fatty acid content in the fermented rye after either 24 h or 48 h is presented in
Table 6. Acetic acid concentration in the fermented rye after either 24 h or 48 h was significantly the greatest in the DM content of 25% (3.02 and 2.32 g/kg DM, respectively) compared to other treatments. While, no significant differences were noted in the acetic acid content among the treatments in the different DM contents (37.5–75%) of fermented rye (24 h or 48 h). After fermenting the rye for 24 h, the propionic acid concentration was the highest with a DM content of 37.5% (0.12 g/kg DM) compared to the other treatments. No concentration of propionic acid was found in the case of rye in DM contents of 62.5% and 75% after 48 h fermentation, while it was in a range of 0.05–0.08 g/kg DM for fermented rye after 48 h with 25–50% DM contents. Butyric acid concentration in the fermented rye after 24 h was significantly the greatest in the DM content of 25% (0.72 g/kg DM) compared to other treatments, while no significant differences were noted in the butyric acid concentration in the DM contents of 37.5–75% for fermented rye (24 h) among the treatments. Nonetheless, no significant differences were noted in the butyric acid concentration among the treatments of different DM contents.
The fatty acid content in the fermented sorghum after either 24 h or 48 h are presented in
Table 6. Acetic acid concentration in the fermented sorghum after either 24 h or 48 h was significantly the greatest in the DM content of 25% (1.49 and 1.97 g/kg DM, respectively) compared to other treatments. While, the acetic content was significantly the lowest in the fermented sorghum either after 24 h or 48 h for 75% DM (0.16 and 0.23 g/kg DM, respectively). After fermenting the sorghum for 24 h and 48 h, the propionic acid concentration was the lowest with a DM content of 75% (0.02 g/kg DM) compared to other treatments. Butyric acid concentration in the fermented sorghum after either 24 h or 48 h was significantly the greatest in the DM content of 25% (0.08 and 0.12 g/kg DM) compared to other treatments, while no concentration of butyric acid was detected in any of the treatments with DM contents of 62.5–75% of sorghum fermented for 48 h.
3.7. Viscosity
The data of viscosity measurement in rye during different fermentation periods are displayed in
Table 7. At 0 h of fermentation, the rye with 62.5% and 75% DM contents had the highest viscosity values (4.52 and 4.65 mPa·s, respectively) compared to the other treatments. After fermenting the rye for 6 h, the DM content of 75% showed a significantly higher viscosity value (4.62 mPa·s) compared to those contained in 25–50% DM contents (range: 3.37–3.62 mPa·s). When increasing the fermentation period to 12 h, the 75% DM content had significantly the highest viscosity value (4.65 mPa·s), while the 25% DM content had the lowest value (1.72 mPa·s). Following the same trend, when increasing the fermentation process of rye up to 24 h and 48 h, the viscosity value was the highest in the case of 75% DM content (4.65 and 4.61 mPa·s, respectively), while the lowest value was recorded in the case of 25% and 37.5% DM contents (1.52 and 1.55, respectively for 24 h). Also, the lowest viscosity values were observed after fermenting the rye for 48 h in DM contents of 25–50% (range: 1.38–1.65 mPa·s).
The data of viscosity measurement in sorghum during different fermentation periods are presented in
Table 7. At 0 h of fermentation, the sorghum with 75% DM content had the highest viscosity values (1.72 mPa·s) compared to the other treatments. When increasing the fermentation period from 12 h to 48 h, the 75% DM had significantly the highest viscosity values (range: 1.21–1.31 mPa·s), while no significant differences were noted among the other treatments in 25–62.5% DM contents.
3.8. Specific Microscopic Features
Figure 3 and
Figure 4 show the morphological features of rye and sorghum before and after fermentation by scanning electron microscope at ×1000 magnifications. Scanning electron microscopy after fermentation, initially revealed no damage to the lateral surface of large granules. In rye, characteristic structural changes such as partial or complete lack of cell walls were visible, with partial or complete disappearance of the protein matrix. In sorghum, the peripheral regions of large granules contained numerous, large and deep holes; the lateral surfaces were found to contain different sized hollows, some directed towards the inside of the granule. No uniformity was apparent regarding distribution and size of holes on the surface of large granules.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}