
Oenological Characterization of Native Hanseniaspora uvarum Strains

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of the Yeasts
2.2. Fermentative Characteristics
2.3. Agar Plate Screening for Aroma Production
2.4. Microvinification
2.5. HPLC Analysis
2.6. Statistical Analysis
3. Results
3.1. Isolation and Identification of the Yeasts
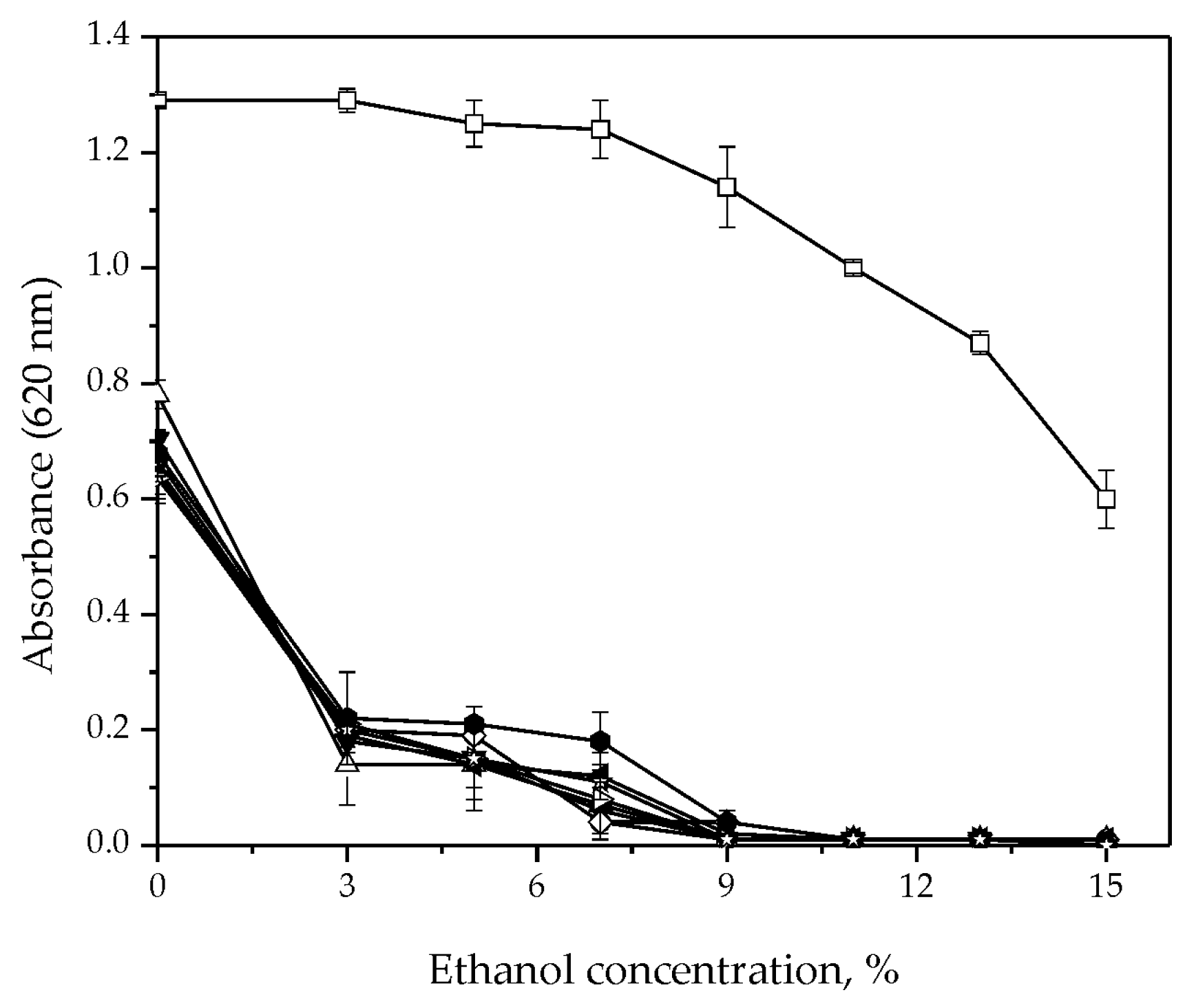
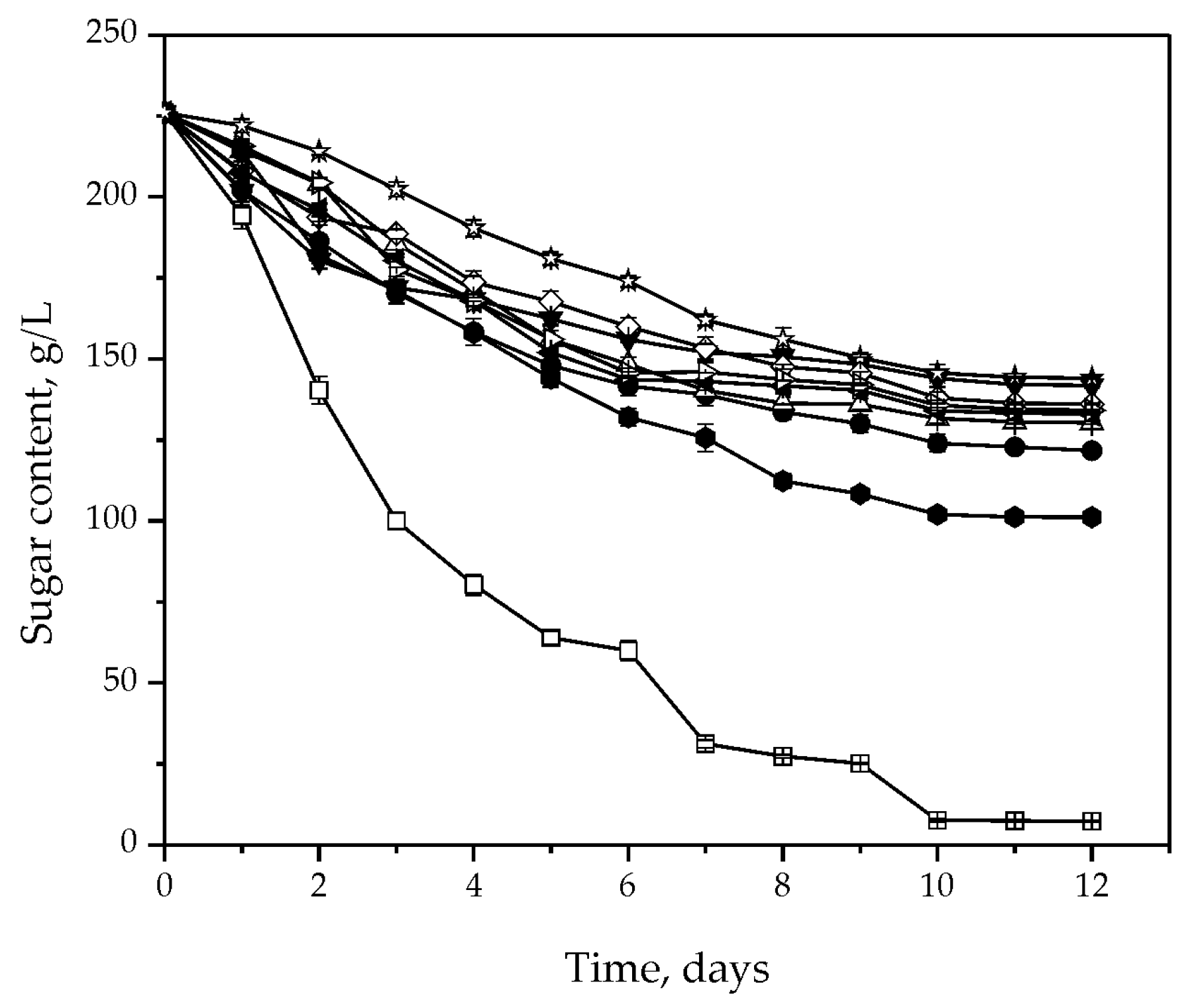
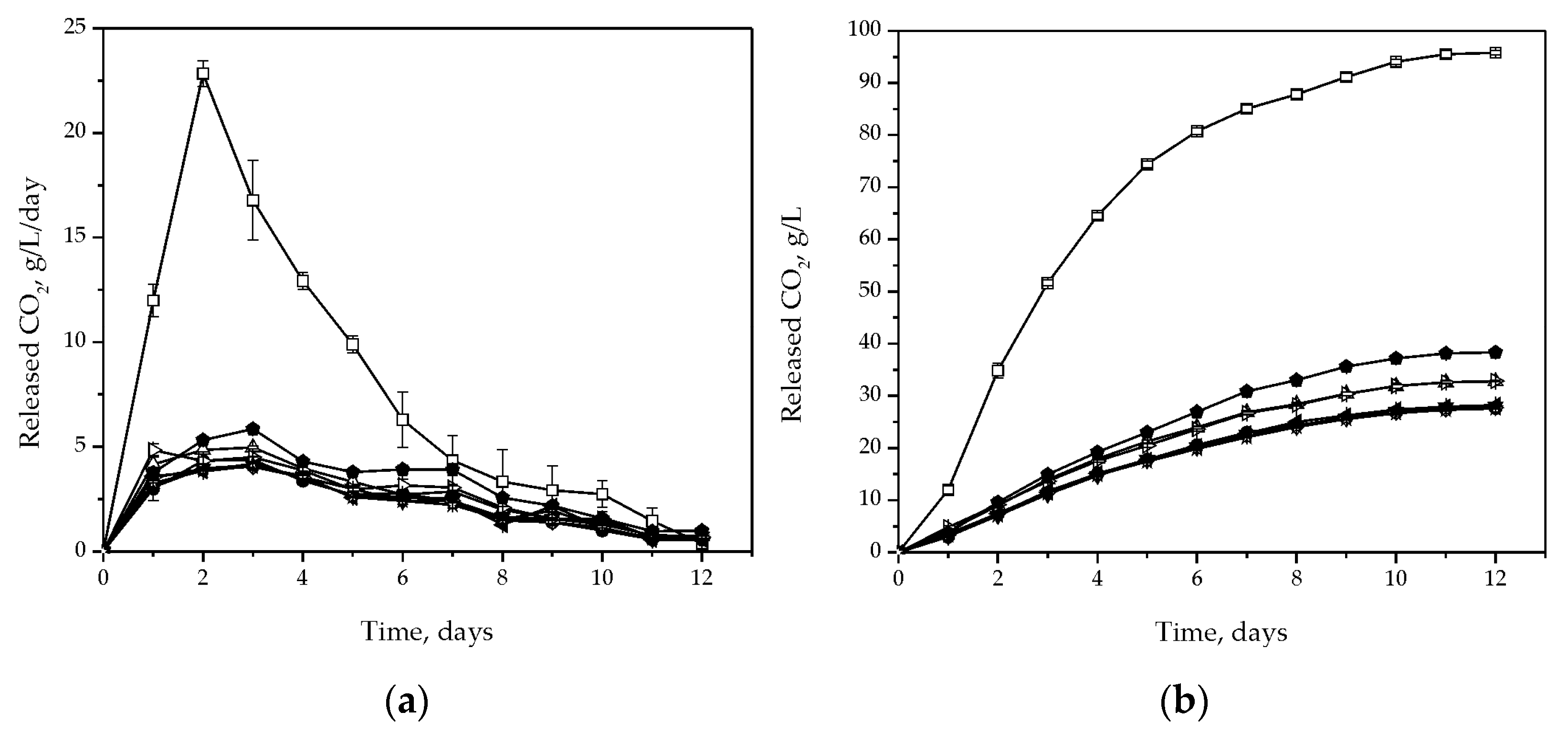
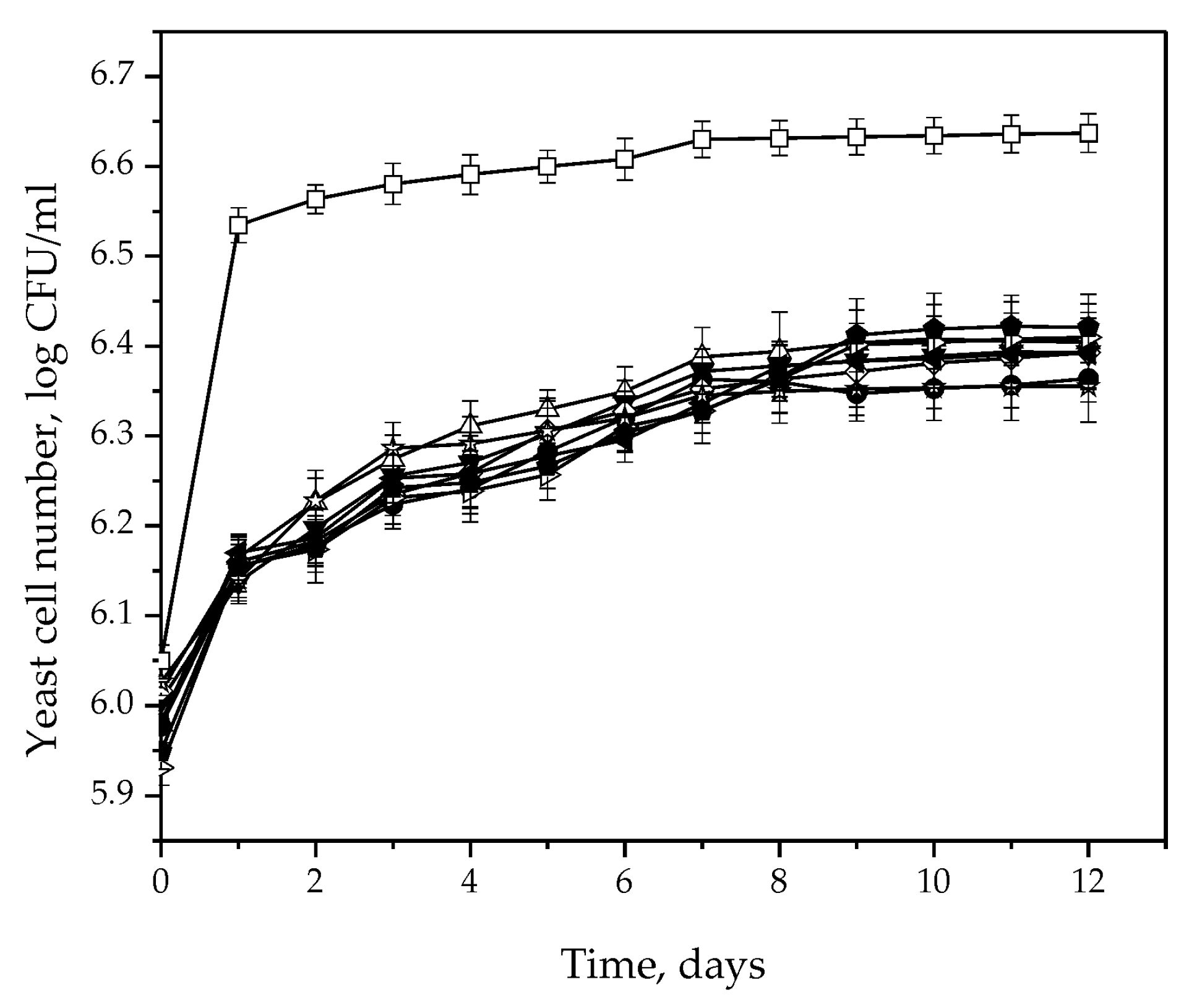
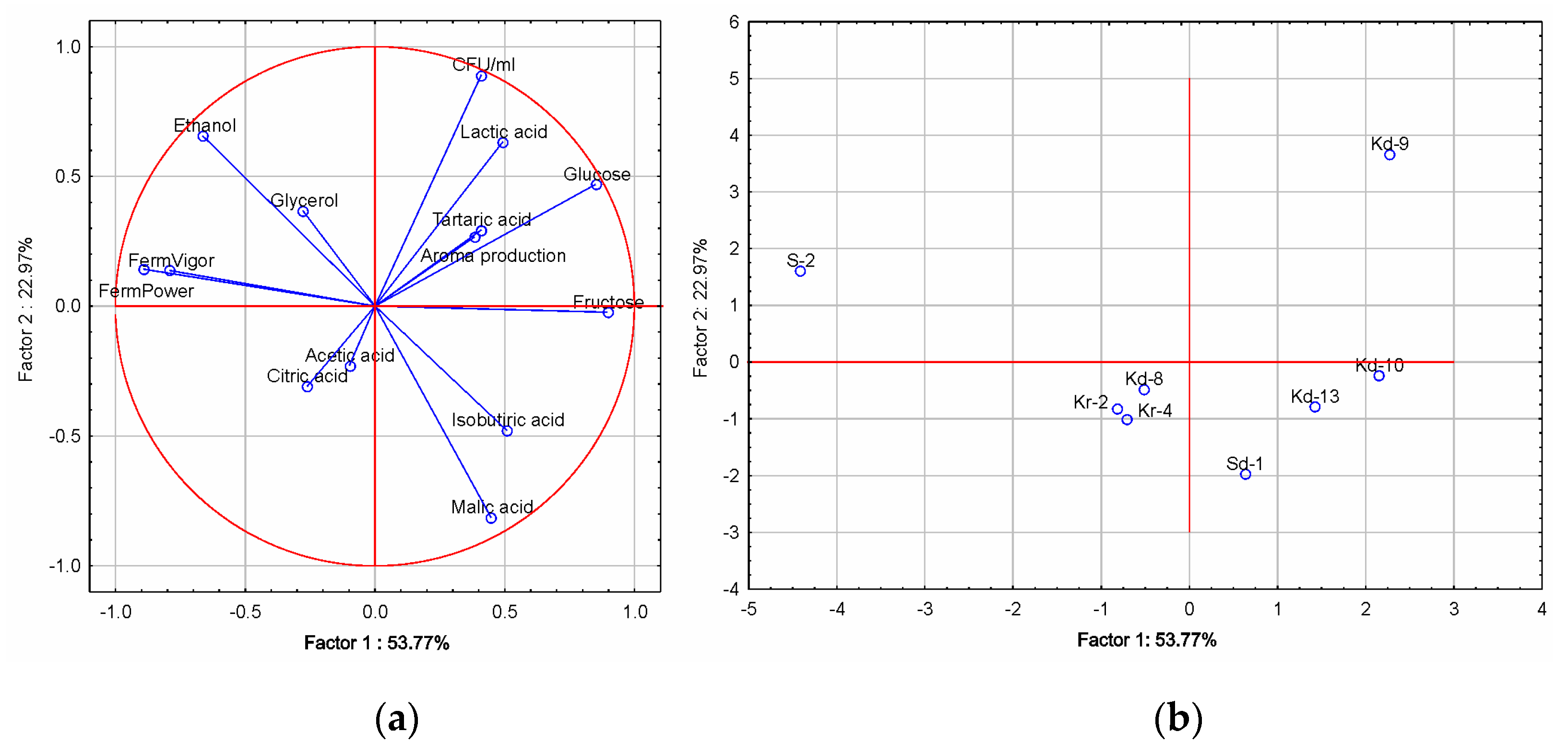
3.2. Fermentative Characteristics and Aroma Production
3.3. Chemical Composition of the Wine
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karabegović, I.; Malićanin, M.; Danilović, B.; Stanojević, J.; Stojanović, S.S.; Nikolić, N.; Lazić, M. Potential of non-Saccharomyces yeast for improving the aroma and sensory profile of Prokupac red wine. Oeno One 2021, 55, 181–195. [Google Scholar] [CrossRef]
- Mančić, S.; Danilović, B.; Malićanin, M.; Stojanović, S.S.; Nikolić, N.; Lazić, M.; Karabegović, I. Fermentative potential of native yeast Candida famata for Prokupac grape must fermentation. Agriculture 2021, 11, 358. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Morata, A. New trends in yeast selection for winemaking. Trends Food Sci. Technol. 2012, 23, 39–50. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Arroyo, T. Non-Saccharomyces yeasts: Biotechnological role for wine production. In Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2016. [Google Scholar]
- Larroque, M.N.; Carrau, F.; Fariña, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef] [PubMed]
- Rollero, S.; Bloem, A.; Brand, J.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Nitrogen metabolism in three non-conventional wine yeast species: A tool to modulate wine aroma profiles. Food Microbiol. 2021, 94, 103650. [Google Scholar] [CrossRef]
- Seguinot, P.; Bloem, A.; Brial, P.; Meudec, E.; Ortiz-Julien, A.; Camarasa, C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-Saccharomyces strains. Int. J. Food Microbiol. 2020, 316, 108441. [Google Scholar] [CrossRef]
- Zhang, B.; Ivanova-Petropulos, V.; Duan, C.; Yan, G. Distinctive chemical and aromatic composition of red wines produced by Saccharomyces cerevisiae co-fermentation with indigenous and commercial non-Saccharomyces strains. Food Biosci. 2021, 41, 100925. [Google Scholar] [CrossRef]
- Testa, B.; Coppola, F.; Lombardi, S.J.; Iorizzo, M.; Letizia, F.; Di Renzo, M.; Succi, M.; Tremonte, P. Influence of Hanseniaspora uvarum as27 on chemical and sensorial characteristics of aglianico wine. Processes 2021, 9, 326. [Google Scholar] [CrossRef]
- Jolly, P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Picciotti, G. The growth kinetics and fermentation behaviour of some non-Saccharomyces yeasts associated with wine-making. Biotechnol. Lett. 1995, 17, 1247–1250. [Google Scholar] [CrossRef]
- Cadez, N.; Raspor, P.; De Cock, A.W.A.M.; Boekhout, T.; Smith, M.T. Molecular identification and genetic diversity within species of the genera Hanseniaspora and Kloeckera. FEMS Yeast Res. 2002, 1, 279–289. [Google Scholar] [CrossRef]
- Martin, V.; Jose Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Brilli, L.; Buscioni, G.; Moriondo, M.; Bindi, M.; Vincenzini, M. Influence of interannual meteorological variability on yeast content and composition in sangiovese grapes. Am. J. Enol. Vitic. 2014, 65, 375–380. [Google Scholar] [CrossRef]
- De Benedictis, M.; Bleve, G.; Tristezza, M.; Tufariello, M.; Grieco, F. An optimized procedure for the enological selection of non-Saccharomyces starter cultures. Antonie Leeuwenhoek 2011, 99, 189–200. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Hall, H.; Zhou, Q.; Qian, M.C.; Osborne, J.P. Impact of yeasts present during prefermentation cold maceration of pinot noir grapes on wine volatile aromas. Am. J. Enol. Vitic. 2017, 68, 81–90. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Comi, G.; Romano, P.; Cocolin, L.; Fiore, C. Characterization of Kloeckera apiculata strains from the Friuli region in Northern Italy. World J. Microbiol. Biotechnol. 2001, 17, 391–394. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G.; Comi, G.; Zironi, R.; Maifreni, M. Glycerol and other fermentation products of apiculate wine yeasts. J. Appl. Microbiol. 1997, 82, 615–618. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The effect of non-Saccharomyces yeasts on fermentation and wine quality. S. Afr. J. Enol. Vitic. 2003, 24, 8–10. [Google Scholar] [CrossRef]
- Nieuwoudt, H.H.; Prior, B.A.; Pretorius, I.S. Glycerol in South African table wines: An assessment of its relationship to wine quality. S. Afr. J. Enol. Vitic. 2002, 23, 22–30. [Google Scholar] [CrossRef]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Silva, R.D.; Chaves, S.R.; Sousa, M.J.; Côrte-Real, M. The impact of acetate metabolism on yeast fermentative performance and wine quality: Reduction of volatile acidity of grape musts and wines. Appl. Microbiol. Biotechnol. 2011, 89, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappa, I.K.; Kachrimanidou, V.; Pateraki, C.; Koulougliotis, D.; Eriotou, E.; Kopsahelis, N. Indigenous yeasts: Emerging trends and challenges in winemaking. Curr. Opin. Food Sci. 2020, 32, 133–143. [Google Scholar] [CrossRef]
- Di Gennaro, S.F.; Matese, A.; Mancin, M.; Primicerio, J.; Palliotti, A. An open-source and low-cost monitoring system for precision enology. Sensors 2014, 14, 23388–23397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantelić, M.; Dabić Zagorac, D.; Gašić, U.; Jović, S.; Bešlić, Z.; Todić, S.; Natić, M. Phenolic profiles of Serbian autochthonous variety ‘Prokupac’ and monovarietal international wines from the Central Serbia wine region. Nat. Prod. Res. 2018, 32, 2356–2359. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Nikolić-Milojević, N.; Mošić, I.; Karabegović, I.; Lazić, M.; Nikolić, N.; Perić, S.; Golubović, S.; Davidović, D.; Veličković, D. Influence of the time of maceration on phenolic composition of wines produced from the indigenous variety Prokupac. Biologica Nyssana 2020, 11, 121–128. [Google Scholar] [CrossRef]
- Castellari, M.; Versari, A.; Spinabelli, U.; Galassi, S.; Amati, A. An improved HPLC method for the analysis of organic acids, carbohydrates and alchocols in grape musts and wines. J. Liq. Chromatogr. Relat. Technol. 2007, 23, 2047–2056. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Mendes-Faia, A. The role of yeasts and lactic acid bacteria on the metabolism of organic acids during winemaking. Foods 2020, 9, 1231. [Google Scholar] [CrossRef]
- Jackson, R.S. Fermentation. In Wine Science; Jackson, R.S., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 427–534. ISBN 978-0-12-381468-5. [Google Scholar]
- Vontrobová, E.; Kubizniakova, P.; Fiala, J.; Sochor, J.; Matoulková, D. Autochthonous yeasts as one of the tools to produce wines by original technologies. Kvas. Prum. 2019, 65, 38–45. [Google Scholar] [CrossRef]
- Bink, F.J. Molekulargenetische und Physiologische Untersuchungen an der Weinhefe Kloeckera apiculata (Hanseniaspora uvarum). Ph.D. Thesis, Osnabrück University, Osnabrück, Germany, 2010. [Google Scholar]
- Hierro, N.; González, Á.; Mas, A.; Guillamón, J.M. Diversity and evolution of non-Saccharomyces yeast populations during wine fermentation: Effect of grape ripeness and cold maceration. FEMS Yeast Res. 2006, 6, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ough, C.S.; Crowell, E.A. Use of sulfur dioxide in winemaking. J. Food Sci. 1987, 52, 386–388. [Google Scholar] [CrossRef]
- Máleková, E.; Lakatošová, J.; Jankura, E. Identification and technological characterization of naturally occurring yeasts on grapevines. In Proceedings of the 39th World Congress of Vine and Wine, Bento Gonçalves, Brazil, 23–28 October 2016. [Google Scholar]
- Jankura, E.; Piknová, L.; Lopašovská, J. Identification and technological characteristics of yeast strains from vineyards in slovakia. J. Food Nutr. Res. 2020, 59, 241–249. [Google Scholar]
- Snoek, T.; Verstrepen, K.J.; Voordeckers, K. How do yeast cells become tolerant to high ethanol concentrations? Curr. Genet. 2016, 62, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.P.; Du Le, H.; Le, V.V.M. Effect of ethanol stress on fermentation performance of Saccharomyces cerevisiae cells immobilized on Nypa fruticans leaf sheath pieces. Food Technol. Biotechnol. 2015, 53, 96. [Google Scholar] [CrossRef]
- Satora, P.; Tuszyński, T. Influence of indigenous yeasts on the fermentation and volatile profile of plum brandies. Food Microbiol. 2010, 27, 418–424. [Google Scholar] [CrossRef]
- Zohre, D.E.; Erten, H. The influence of Kloeckera apiculata and Candida pulcherrima yeasts on wine fermentation. Process Biochem. 2002, 38, 319–324. [Google Scholar] [CrossRef]
- González-Robles, I.W.; Estarrón-Espinosa, M.; Díaz-Montaño, D.M. Fermentative capabilities and volatile compounds produced by Kloeckera/Hanseniaspora and Saccharomyces yeast strains in pure and mixed cultures during Agave tequilana juice fermentation. Antonie Leeuwenhoek 2015, 108, 525–536. [Google Scholar] [CrossRef]
- Mendoza, L.M.; De Nadra, M.C.M.; Farías, M.E. Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef]
- Mendoza, L.M.; De Nadra, M.C.M.; Bru, E.; Farías, M.E. Influence of wine-related physicochemical factors on the growth and metabolism of non-Saccharomyces and Saccharomyces yeasts in mixed culture. J. Ind. Microbiol. Biotechnol. 2009, 36, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, A.; Irastorza, A.; Dueñas, M.; Fernandez, K. The effect of temperature on the growth of strains of Kloeckera apiculata and Saccharomyces cerevisiae in apple juice fermentation. Lett. Appl. Microbiol. 1997, 24, 37–39. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Fleet, G.H. Degradation of malic and tartaric acids by high density cell suspensions of wine yeasts. Food Microbiol. 1995, 12, 65–71. [Google Scholar] [CrossRef]
- Bauer, F.; Chidi, B.S.; Rossouw, D.; Buica, A.S.; Bauer, F.F. Determining the Impact of Industrial wine yeast trains on organic acid production under white and red wine-like fermentation conditions. S. Afr. J. Enol. Vitic. 2015, 36, 316–327. [Google Scholar]
- Fonseca, A. Utilization of tartaric acid and related compounds by yeasts: Taxonomic implications. Can. J. Microbiol. 2011, 38, 1242–1251. [Google Scholar] [CrossRef]
- Ivanova-Petropulos, V.; Petruševa, D.; Mitrev, S. Rapid and simple method for determination of target organic acids in wine using HPLC-DAD analysis. Food Anal. Methods 2020, 13, 1078–1087. [Google Scholar] [CrossRef]
- Geffroy, O.; Lopez, R.; Feilhes, C.; Violleau, F.; Kleiber, D.; Favarel, J.L.; Ferreira, V. Modulating analytical characteristics of thermovinified Carignan musts and the volatile composition of the resulting wines through the heating temperature. Food Chem. 2018, 257, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Chidi, B.S.; Bauer, F.F.; Rossouw, D. Organic acid metabolism and the impact of fermentation practices on wine acidity—A review. S. Afr. J. Enol. Vitic. 2018, 39, 315–329. [Google Scholar] [CrossRef] [Green Version]
- Comuzzo, P.; Battistutta, F. Acidification and pH control in red wines. In Red Wine Technology; Morata, A., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 17–34. ISBN 9780128144008. [Google Scholar]
- Yılmaz, C.; Gökmen, V. Formation of amino acid derivatives in white and red wines during fermentation: Effects of non-Saccharomyces yeasts and Oenococcus oeni. Food Chem. 2021, 343, 128415. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. S. Afr. J. Enol. Vitic. 2019, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Estela-Escalante, W.D.; Rychtera, M.; Melzoch, K.; Guerrero-Ochoa, M.R. Influence of aeration in the fermentative activity of Kloeckera apiculate during fermentation of apple juice. Acta Biol. Colomb. 2012, 17, 309–322. [Google Scholar]
- Jeromel, A.; Korenika, A.M.J.; Tomaz, I. An influence of different yeast species on wine aroma composition. In Fermented Beverages; Grumezascu, A.M., Holban, A.M., Eds.; Woodhead publishing: Duxford, UK, 2019; pp. 171–285. ISBN 9780128152713. [Google Scholar]
- Vázquez-Pateiro, I.; Arias-González, U.; Mirás-Avalos, J.M.; Falqué, E. Evolution of the aroma of Treixadura wines during bottle aging. Foods 2020, 9, 1419. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liu, Y.; Yue, T.X.; Zhang, Z.W. Comparison between aroma compounds in wines from four Vitis vinifera grape varieties grown in different shoot positions. Food Sci. Technol. 2015, 35, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Mateus, D.; Sousa, S.; Coimba, C.; Rogerson, F.; Simões, J. Identification and characterization of Non-Saccharomyces species isolated from port wine spontaneous fermentations. Foods 2020, 9, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivit, N.N.; Longo, R.; Kemp, B. The effect of non-Saccharomyces and Saccharomyces non-Cerevisiae yeasts on ethanol and glycerol levels in wine. Fermentation 2020, 6, 77. [Google Scholar] [CrossRef]
- Iris, L.; Antonio, M.; Antonia, B.M.; Antonio, S.L.J. Isolation, Selection, and Identification Techniques for Non-Saccharomyces Yeasts of Oenological Interest. In Biotechnological Progress and Beverage Consumption; Grumezascu, A.M., Holban, A.M., Eds.; Woodhead publishing: Duxford, UK, 2019; pp. 467–508. ISBN 9780128166789. [Google Scholar]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima selected strain for ethanol reduction in wine: Influence of cell immobilization and aeration condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pérez, M.P.; Bautista-Ortín, A.B.; Pérez-Porras, P.; Jurado, R.; Gómez-Plaza, E. A new approach to the reduction of alcohol content in red wines: The use of high-power ultrasounds. Foods 2020, 9, 726. [Google Scholar] [CrossRef]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17, 10. [Google Scholar] [CrossRef] [Green Version]
- Mestre, M.V.; Maturano, Y.P.; Gallardo, C.; Combina, M.; Mercado, L.; Toro, M.E.; Carrau, F.; Vazquez, F.; Dellacassa, E. Impact on sensory and aromatic profile of low ethanol Malbec wines fermented by sequential culture of Hanseniaspora uvarum and Saccharomyces cerevisiae native yeasts. Fermentation 2019, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Fernández-González, M.; Di Stefano, R.; Briones, A. Hydrolysis and transformation of terpene glycosides from muscat must by different yeast species. Food Microbiol. 2003, 20, 35–41. [Google Scholar] [CrossRef]
- Zhu, X.; Torija, M.J.; Mas, A.; Beltran, G.; Navarro, Y. Effect of a multistarter yeast inoculum on ethanol reduction and population dynamics in wine fermentation. Foods 2021, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Eglinton, J.M.; Heinrich, A.J.; Pollnitz, A.P.; Langridge, P.; Henschke, P.A.; De Barros Lopes, E.M. Decreasing acetic acid accumulation by a glycerol overproducing strain of Saccharomyces cerevisiae by deleting the ALD6 aldehyde dehydrogenase gene. Yeast 2002, 19, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The Oenological Potential of Hanseniaspora uvarum in Simultaneous and Sequential Co-fermentation with Saccharomyces cerevisiae for Industrial Wine Production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic acid bacteria in wine: Technological advances and evaluation of their functional role. Front. Microbiol. 2021, 11, 3192. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Jagatić Korenika, A.M.; Tomaz, I.; Preiner, D.; Plichta, V.; Jeromel, A. Impact of Commercial Yeasts on Phenolic Profile of Plavac Mali Wines from Croatia. Fermentation 2021, 7, 92. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Colomo, B.; Suárez, J.A. Pyruvic Acid and Acetaldehyde Production by Different Strains of Saccharomyces cerevisiae: Relationship with Vitisin A and B Formation in Red Wines. J. Agric. Food Chem. 2003, 51, 7402–7409. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Effects of Non-Saccharomyces Yeasts on Color, Anthocyanin, and Anthocyanin-Derived Pigments of Tannat Grapes during Fermentation. Am. J. Enol. Vitic. 2018, 69, 148–156. [Google Scholar] [CrossRef]
- Tofalo, R.; Suzzi, G.; Perpetuini, G. Discovering the Influence of Microorganisms on Wine Color. Front. Microbiol. 2021, 12, 790935. [Google Scholar] [CrossRef] [PubMed]
- Malićanin, M.; Danilović, B.; Cvetković, D.; Stamenković-Stojanović, S.; Nikolić, N.; Lazić, M.; Karabegović, I. Modulation of Aroma and Sensory Properties of Prokupac Wines by a Bacillus-based Preparation Applied to Grapes Prior to Harvest. S. Afr. J. Enol. Vitic. 2020, 41, 158–167. [Google Scholar] [CrossRef]
- Lakićević, S.; Popović, T.; Matijašević, S.; Ćirković, B.; Lazić, M.; Popović-Đorđević, J. Chemical evaluation of autochthonous variety “Prokupac” red wine with the addition of selected aromatic herbs. Ann. Univ. Craiova Agric. Mont. Cadastre Ser. 2019, 49, 87–97. [Google Scholar]
- Merkytė, V.; Longo, E.; Windisch, G.; Boselli, E. Phenolic Compounds as Markers of Wine Quality and Authenticity. Foods 2020, 9, 1785. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | H. uvarum Isolate | S. cerevisiae | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Kd-8 | Kd-9 | Kd-10 | Kd-13 | Kr-2 | Kr-4 | S-2 | Sd-1 | |||
| Cell number in SDB at 25 °C, log CFU/mL | 7.30 ± 0.04 a | 7.51 ± 0.05 b | 7.37 ± 0.12 a | 7.28 ± 0.01 a | 7.26 ± 0.06 a | 7.24 ± 0.09 a | 7.32 ± 0.12 a | 7.25 ± 0.07 a | 7.90 ± 0.1 c | |
| CO2 production | + | + | + | + | + | + | + | + | + | |
| Growth at different temperatures | 4 °C | + | + | + | + | + | + | + | + | + |
| 10 °C | + | + | + | + | + | + | + | + | + | |
| 15 °C | + | + | + | + | + | + | + | + | + | |
| 20 °C | + | + | + | + | + | + | + | + | + | |
| Tolerance to SO2 | 50 ppm | + | + | + | + | + | + | + | + | + |
| 100 ppm | + | + | + | + | + | + | + | + | + | |
| 200 ppm | + | + | + | + | + | + | + | + | + | |
| 300 ppm | + | + | + | + | + | + | + | + | + | |
| Fermentative vigor 1 | 1.44 ± 0.06 a | 1.22 ± 0.03 b | 1.00 ± 0.03 c | 1.21 ± 0.03 b | 1.20 ± 0.03 b | 1.41 ± 0.04 a | 1.51 ± 0.03 d | 1.20 ± 0.03 b | 3.40 ± 0.09 e | |
| Fermentative power 2 | 3.30 ± 0.04 a | 2.78 ± 0.03 b | 2.42 ± 0.02 c | 2.83 ± 0.03 b | 2.84 ± 0.02 b | 3.30 ± 0.05 a | 3.76 ± 0.04 d | 2.81 ± 0.03 b | 6.20 ± 0.04 e | |
| Aroma production 3 | P++ | P+++ | P++ | P++ | U+++ | P++ | P+ | P++ | P+++ | |
| Compound | Grape Must | H. uvarum Isolate | S. cerevisiae | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Kd-8 | Kd-9 | Kd-10 | Kd-13 | Kr-2 | Kr-4 | S-2 | Sd-1 | |||
| Tartaric acid | 5.85 ± 0.29 a | 5.07 ± 0.16 b,c,d | 5.19 ± 0.04 b,c | 5.03 ± 0.08 b,d | 5.28 ± 0.10 c | 4.87 ± 0.10 d | 4.71 ± 0.07 e | 5.03 ± 0.15 b,c,d | 5.12 ± 0.09 b,c | 4.56 ± 0.09 f |
| Malic acid | 3.23 ± 0.01 a | 3.00 ± 0.11 b | 2.54 ± 0.12 c | 2.85 ± 0.07 d | 2.96 ± 0.10 b,d,e | 2.86 ± 0.05 b,d | 3.11 ± 0.09 b,e | 2.22 ± 0.04 f | 3.22 ± 0.06 a | 1.26 ± 0.05 g |
| Lactic acid | / | / | 0.20 ± 0.06 a | 0.09 ± 0.06 a | / | / | 0.11 ± 0.04 a | / | / | 1.17 ± 0.13 b |
| Acetic acid | / | 0.63 ± 0.07 a | 0.58 ± 0.02 a | 0.56 ± 0.01 a | 0.54 ± 0.08 a | 0.60 ± 0.12 a | 0.56 ± 0.06 a | 0.57 ± 0.05 a | 0.62 ± 0.07 a | 0.49 ± 0.02 b |
| Citric acid | / | / | / | 0.10 ± 0.03 a | 0.10 ± 0.06 a | / | 0.12 ± 0.05 a | 0.13 ± 0.04 a | 0.11 ± 0.05 a | 0.01 ± 0.01 b |
| Isobutyric acid | / | / | / | 0.38 ± 0.05 a | 0.24 ± 0.06 b | / | 0.20 ± 0.05 b | / | 0.21 ± 0.06 b | / |
| Glucose | 120.85 ± 3.24 a | 57.57 ± 0.10 b | 76.31 ± 0.10 c | 62.09 ± 1.82 d | 64.16 ± 0.20 e | 56.41 ± 0.66 b | 57.17 ± 0.10 b | 47.62 ± 0.06 f | 57.25 ± 0.08 b | 0.94 ± 0.07 g |
| Fructose | 115.43 ± 2.78 a | 53.60 ± 0.07 b | 60.72 ± 0.12 c | 51.21 ± 0.22 d | 56.13 ± 0.14 e | 49.12 ± 0.10 f | 50.95 ± 0.08 d | 29.97 ± 0.04 g | 51.36 ± 0.39 d | 2.52 ± 0.14 h |
| Ethanol * | / | 3.5 ± 0.02 a | 4.16 ± 0.01 b | 3.13 ± 0.02 c | 3.2 ± 0.02 d | 3.55 ± 0.01 e | 3.88 ± 0.01 f | 4.68 ± 0.01 g | 3.47 ± 0.01 h | 12.14 ± 0.01 i |
| Glycerol | / | 3.47 ± 0.06 a | 3.74 ± 0.07 b | 4.33 ± 0.08 c | 3.74 ± 0.08 b | 4.07 ± 0.10 d | 3.37 ± 0.07 e | 4.40 ± 0.06 c | 3.35 ± 0.08 e | 2.27 ± 0.15 f |
| Parameter | Fermentation Protocol | ||
|---|---|---|---|
| HU 1 | HUSC 2 | SC 3 | |
| Total phenols, g/L GAE | 1.54 ± 0.11 a | 1.61 ± 0.11 a | 1.77 ± 0.14 a |
| Total flavonoids, mg/L CE | 784.8 ± 12.1 a | 844.5 ± 18.7 b | 848.4 ± 8.4 b |
| Total anthocyanins, mg/L | 204.5 ± 3.66 a | 212.4 ± 4.16 b | 214.8 ± 3.88 b |
| Color intensity | 1.01 ± 0.04 a | 1.05 ± 0.01 a | 0.99 ± 0.02 a |
| Color hue | 1.21 ± 0.09 a | 1.07 ± 0.03 b | 0.92 ± 0.03 c |
| HPLC analysis | |||
| Tartaric acid, g/L | 4.70 ± 0.01 a | 4.81 ± 0.03 b | 5.82 ± 0.06 c |
| Malic acid, g/L | 1.15 ± 0.07 a | 1.15 ± 0.08 a | 3.10 ± 0.01 b |
| Lactic acid, g/L | 0.75 ± 0.03 a | 0.42 ± 0.01 b | 0.28 ± 0.02 c |
| Succinic acid, g/L | 0.38 ± 0.01 a | 0.38 ± 0.01 a | 0.48 ± 0.01 b |
| Acetic acid, g/L | 0.52 ± 0.01 a | 0.36 ± 0.01 b | 0.24 ± 0.00 c |
| Glucose, g/L | 0.81 ± 0.00 a | 0.92 ± 0.01 b | 0.92 ± 0.02 b |
| Fructose, g/L | 1.26 ± 0.00 a | 1.26 ± 0.00 a | 1.06 ± 0.00 b |
| Ethanol, % v/v | 11.41 ± 0.02 a | 11.90 ± 0.15 b | 12.46 ± 0.09 c |
| Glycerol, g/L | 7.41 ± 0.02 a | 7.36 ± 0.13 a | 4.55 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mančić, S.; Stamenković Stojanović, S.; Danilović, B.; Djordjević, N.; Malićanin, M.; Lazić, M.; Karabegović, I. Oenological Characterization of Native Hanseniaspora uvarum Strains. Fermentation 2022, 8, 92. https://doi.org/10.3390/fermentation8030092
Mančić S, Stamenković Stojanović S, Danilović B, Djordjević N, Malićanin M, Lazić M, Karabegović I. Oenological Characterization of Native Hanseniaspora uvarum Strains. Fermentation. 2022; 8(3):92. https://doi.org/10.3390/fermentation8030092
Chicago/Turabian StyleMančić, Stojan, Sandra Stamenković Stojanović, Bojana Danilović, Natalija Djordjević, Marko Malićanin, Miodrag Lazić, and Ivana Karabegović. 2022. "Oenological Characterization of Native Hanseniaspora uvarum Strains" Fermentation 8, no. 3: 92. https://doi.org/10.3390/fermentation8030092