1. Introduction
The brewing of Baijiu has existed for thousands of years in China, representing an intangible cultural heritage of the nation [
1]. Baijiu is a typical alcoholic beverage which is characterized by semi-solid/solid-state fermentation with Daqu as a kind of saccharifying starter [
2,
3]. According to various characteristics of flavor, Baijiu can be divided into three main types: Jiang-flavor (Maotai), strong-flavor (Luzhou), and light-flavor (Fen) [
4]. Among these three types, the Fen-flavor Baijiu is favored by consumers for its unique fermentation technology and taste [
5,
6]. The main steps in the production of Fen-flavor Baijiu include solid-state fermentation, repeated batch fermentation, fractional distillation, and extraction [
6]. In the brewing process of Fen-flavor Baijiu, the grains of sorghum are fermented in a tank, and the liquor of Baijiu can be obtained after distillation. The production process of Fen-flavor Baijiu consists of Dacha (DX) and Ercha (EX). The raw materials of grain (sorghum) and Daqu are mixed well and then fermented in the tank; the solid substrate obtained after distillation is Dacha (DX). Following this, Daqu is added to the solid substrate again, and the solid substrate obtained after the fermentation and distillation is Ercha (EX) [
5].
The components of flavors in Baijiu mainly include organic acids, esters, aldehydes, lactones, phenols, heterocycles, terpenes, and aromatic compounds [
6,
7]. Among these compounds, organic acids and esters are the predominant flavors, especially contributing to the characteristic aroma of Fen-flavor Baijiu. The main organic acids are lactic acid and acetic acid, while the main esters are the related ethyl lactate and ethyl acetate in Fen-flavor Baijiu [
8,
9]. As a higher concentration of lactic acid and its derived ethyl lactate will result in an unpleasant astringent taste [
10], the content of lactic acid and ethyl lactate should be strictly controlled to maintain the desired flavor and fine quality of Baijiu [
11]. Therefore, it is necessary to identify the critical influence factors for the formation of lactic acid, acetic acid, ethyl lactate, and ethyl acetate in the fermented grains during the brewing process of Fen-flavor Baijiu.
The composition of microorganisms in fermented grains of Fen-flavor Baijiu is very complex, including bacteria, mold, and yeast, which produce enzymes and form various organic acids and their derived esters [
12]. Thus, these microorganisms are closely associated with the content of lactic acid and ethyl lactate in the fermented grains and play a decisive role in the quality of Fen-flavor Baijiu. Due to the complex environment for the brewing of Fen-flavor Baijiu, bacteria from different sources can produce various enzymes, forming acids and their derivatives in different seasons [
11,
12]. Especially during fermentation in summer, the population of lactic acid bacteria in fermented grains significantly increases, leading to the excessive accumulation of lactic acid and ethyl lactate [
13]. It results in a serious imbalance in the ratio between ethyl acetate and ethyl lactate (<1.0), resulting in a decrease in liquor yield and an adverse impact on Baijiu. Thus, it is essential to compare and analyze the change in microbial community and the correlation with the formation of lactic acid and ethyl lactate in cold and hot seasons using high-throughput omics technology.
Currently, most research mainly focuses on the correlation analysis of microflora and flavor metabolites for the other types of Baijiu [
14,
15,
16]. However, research on Fen-flavor Baijiu is limited and there is a lack of comparative analysis toward the changes in community during different seasons. In this study, the concentration of lactic acid and ethyl lactate and the composition of the microbial community in fermented grains during hot and cold seasons were detected for Fen-flavor Baijiu [
17]. The relationship between bacteria and lactic acid in fermented grains was systematically analyzed to identify the key microorganisms affecting the content of lactic acid in fermented grains during cold and hot seasons. In addition, the dominant lactic-acid-producing strains in fermented grains were isolated and the capacity of acid production and the environmental tolerance for each strain were determined. These results can provide theoretical guidance to control the content of lactic acid and further improve the quality of Fen-flavor Baijiu.
2. Materials and Methods
2.1. Sample Collection
The samples of fermented grains were collected at Shanxi Xinghuacun Fenjiu Distillery co., Ltd. (Fenyang, China). The fermented grains in the cold season were collected from December 2020 to February 2021 (the average temperature in the cold season was 5 °C), while the fermented grains in the hot season were collected from June 2021 to August 2021 (the average temperature in the hot season was 30 °C). These samples were separately obtained from four jars (the volume of each jar is 300 L) for each fermentation time. The samples in jar 1 were taken at the starting time of fermentation and on day 3 of fermentation; the samples in jar 2 were taken on day 7 of fermentation; the samples in jar 4 were taken on day 15 of fermentation; and the samples in jar 4 were taken on day 21 of fermentation and at the end of fermentation. All the samples were frozen quickly by liquid nitrogen and then stored at −80 °C for later use.
2.2. HPLC Detection
Three parallel samples of fermented grains were pretreated as follows: 1.0 g fermented grains were accurately weighed in a 10 mL centrifuge tube, and 6 mL of 50% ethanol solution was added to stir them well. The samples were soaked at room temperature for 30 min then stirred every 10 min during this period and centrifuged at 6000 r for 5 min. The supernatant was collected and 3 mL of 50% ethanol solution was added to wash particles from the precipitate. Centrifugation was repeated to collect the supernatant. All the supernatants were concentrated in a 10 mL volumetric flask for later use. The concentrations of lactic acid, acetic acid, ethyl lactate, and ethyl acetate were measured by HPLC using the previously mentioned methods [
18]. The detections were performed using a T3 column (Waters Inc., Milford, MA, USA) with a mobile phase consisting of acetonitrile-0.05 mol/L KH
2PO
4 (8:92,
v/
v) at a flow rate of 1.0 mL/min. The column temperature was set at 30 °C and the injection volume was 20 μL.
2.3. DNA Eextraction
The specific steps of DNA extraction were executed as follows: 10.0 g of fermented grains was placed in a 50 mL centrifuge tube with 3 glass beads and 20 mL of sterilized PBS buffer solution was added. The samples were oscillated for 5 min and then centrifuged at 150× g for 5 min. The procedure was repeated twice and 5 mL of PBS buffer solution was added and then the mixture was centrifuged at 150× g for 5 min. The supernatant was slowly poured and balanced accurately with PBS buffer solution and then centrifuged at 10,000 r for 10 min. PBS buffer solution was added to the sample for multiple blowing and suction filtrations. The precipitation was collected in a new 5 mL tube and then centrifugation was carried out at 10,000 r for 10 min. A measure of 500 μL of PBS buffer solution was added to the precipitates and then a freeze–thaw process was repeated 3 times (frozen for 30 s by liquid nitrogen followed by a 70 °C water bath for 2 min), An amount of 5 μL of lysozyme was added to the centrifuge tube, which was placed in a water bath at 30 °C for 1 h. Then, 2 μL of lysozyme was added, with a water bath at 37 °C for 30 min and 5 μL of RNA enzyme was added, followed by a water bath at 37 °C for 1 h. Finally, 2.5 μL of protease K was added, with a further water bath at 65 °C for 30 min. The DNA was extracted using QIAamp PowerFecal Pro DNA Kit (QIAGEN Inc., Frankfurt, Germany). The quality of DNA was examined by agarose electrophoresis and the library of DNA was constructed and sequenced as outlined in the following subsections.
2.4. PCR Amplification and 16S rDNA Sequencing
Qubit (Thermo Inc., Waltham, MA, USA) was used for the detection of DNA concentration. Undegraded samples were selected for subsequent agarose integrity detection. To detect the DNA integrity at a voltage of 150 V, 1.5% agarose gel electrophoresis was used. The time was 25 min and broom-like qualified samples were selected for PCR amplification. Universal primers 515F (5′GTGCCAGCMGCCGCGGTAA3′) and 806R (5′GGACTACHVGGGTWTCT3′) for the V4 region of the bacterial 16S rRNA gene were used. The reaction conditions were pre-denaturation for 5 min at 94 °C, followed by 30 cycles. Each cycle was as follows: 94 °C denaturation 30 s; 54 °C annealing 30 s; 72 °C extension 30 s; and after the end of the cycle, 72 °C for an additional 10 min to end the reaction. The product was cut from 1.5% agarose gel and purified by AxyPrep DNA Gel extraction kit (AXYGEN Inc., Union City, CA, USA). The purified product was sequenced by Illumina Hiseq2500 (BGI, Shenzhen, China).
2.5. Data Filtering and Quality Control
The READFQ software was used for data filtering and quality control [
19] and 30 bp was set as the window length. If the average quality value of the window was lower than 20, the end read sequence was cut from the window and the reads whose final read length were lower than 75% of the original read length were removed. Joint contamination reads (the default adapter sequence and read sequence overlap is 15 bp, which is set to 15 bp, and the allowed mismatch number is 3) were removed; reads containing N were removed; and low-complexity reads (10 bp is set to reads where the length of a consecutive base is ≥10) were removed. If the sample was built through a barcode combination, the clean data were obtained and the barcode sequence was used to split the sample through an internally written program.
2.6. Screening and Identification of Key Microorganisms Producing Lactic Acid
Dilute hydrochloric acid was used to adjust the pH of MRS medium to 6.0, and then 8 μg/mL of amphotericin was added to the medium to inhibit the growth of fungi. A certain amount of bromocresol green was added to the sterilized medium as an indicator of acid-producing bacteria. A measure of 10.0 g of fermented grains was added to 100 mL PBS buffer and oscillated at room temperature for 30 min to fully mix the sample and water. Then, 1 mL of supernatant was taken and placed in a test tube with 9 mL of sterile water to obtain 10
−1 diluent and the 10
−2, 10
−3, 10
−4, 10
−5, 10
−6, and 10
−7 dilute sample solutions were similarly prepared. Finally, 0.1 mL of gradient diluent was evenly spread on the medium plate and incubated at 37 °C for 120 h. At the beginning of culturing, the color of the colony was observed every 4 h to mark the green colony in time. The strains with different morphologies in the chromogenic colony were selected and purified on the MRS plate 3 times. The single colony was obtained and then inoculated in a liquid medium. After being cultured at 37 °C for 24 h, it was placed in a glycerol tube and stored at −80°C for the following identification using the previously described methods [
20,
21].
2.7. The Physiological Properties of Key Microorganisms Producing Lactic Acid
The selected strains were activated and cultured in MRS medium at 37 °C and were obtained every 12 h in the first two days and every 24 h in the later period to detect the decline rate of pH values. At each sampling point, the content of lactic acid in the fermentation broth was detected by HPLC. To examine the acidic resistance for each strain, the MRS media were adjusted with lactic acid at pH values of 2.5, 3.0, 3.5, 4.0, 4.5, and 5.0, respectively. To examine the alcoholic resistance for each strain, MRS media with 2%, 4%, 6%, 8%, and 10% ethanol gradient were used. The activated strains were inoculated into the media with different acid and alcohol gradients and were cultured at 37 °C for 24 h. Three replicates were performed for each acid and alcohol gradient and the values of OD
600 were measured by a microplate analyzer (Thermo Inc., Waltham, MA, USA) to evaluate the acidic and alcoholic resistance of strains [
22].
2.8. Statistical Analysis of Data
Alpha diversity analysis was used to compare the differences in the microbial community between cold and hot seasons by the Wilcox test using software R (V3.2.1, Pittsburgh, PA, USA). The bar chart visually displays the species composition and proportion for each sample using software R (V3.4.1, Pittsburgh, PA, USA), reflecting the changes in species among samples. Redundancy analysis (RDA) was used to assess the relationship between abundant microorganisms and environmental factors and analyze the correlation between microbiota and metabolism using Origin 2022 (OriginLab, Northampton, MA, USA). [
23]. In |ρ| > 0.7, Pearson’s correlation coefficient was used to evaluate the correlation between bacterial and metabolites (
p ≤ 0.05). All line charts were drawn using Microsoft Excel (Microsoft, Redmond, WA, USA).
3. Results and Discussion
3.1. The Discrepant Analysis of Lactic Acid during Fermentation in Cold and Hot Seasons
Lactic acid is the dominant organic acid in the fermented grains of Fen-flavor Baijiu [
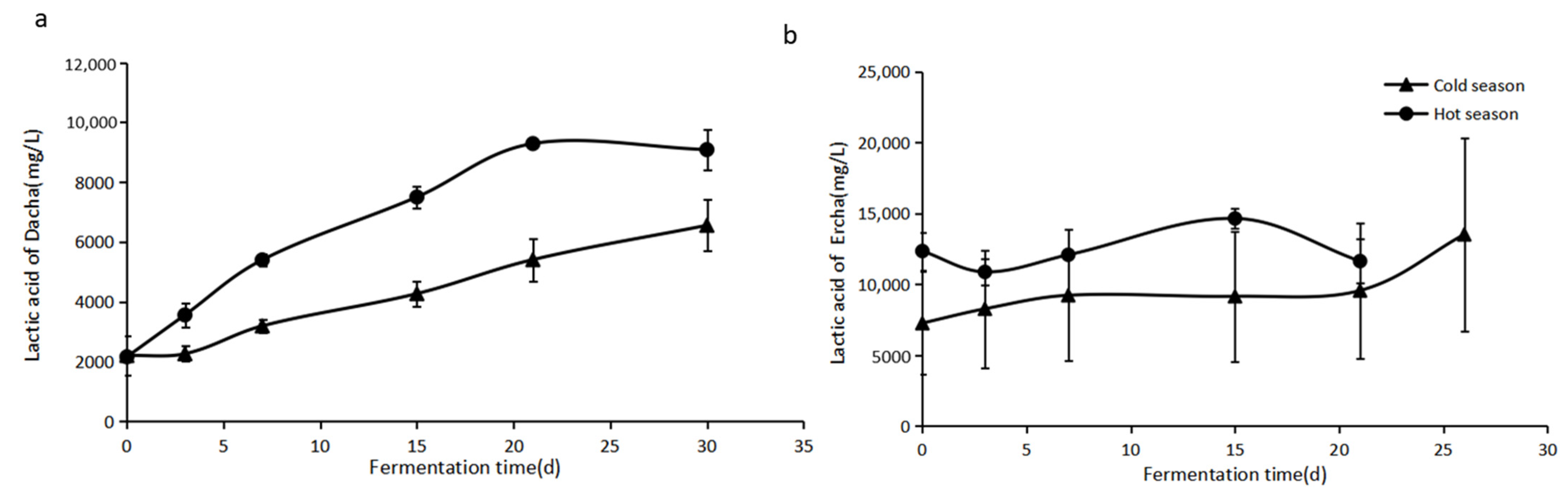
24], and its content varied widely according to the temperature in different seasons. It can be seen from
Figure 1a that the highest concentration of lactic acid in fermented grains reached 9.29 g/L on the 21st day of the Dacha stage, and its concentration was 27.89% higher in the hot season than that in the cold season at the end of Dacha fermentation. As for the Ercha stage, the concentration of lactic acid in fermented grains slightly fluctuated, and the highest concentration reached 14.63 g/L on the 15th day in the hot season and then decreased to 11.62 g/L at the end of Ercha fermentation. The concentration of lactic acid was still lower in the cold season at the early stage of Ercha fermentation, but its content significantly increased from the 21st day, finally reaching 13.50 g/L at the end of Ercha fermentation (
Figure 1b).
In general, the change in lactic acid content in Ercha fermentation was significantly smaller than that in Dacha fermentation, which means the change in lactic acid content in the fermentation of Fen-flavor Baijiu mainly occurred in the early fermentation stage. In the fermentation of strong-flavor Baijiu, the change in physical and chemical properties, including the content of acids, also mainly occurred in the early stage of fermentation (0~25 d) [
14]. Similarly, at the latter stage of fermentation in Jiang-flavor Baijiu (from the third to the eighth rounds), the values of pH and the content of lactic acid in fermented grains gradually tended to be stable [
25], which is consistent with the change in lactic acid content in Fen-flavor Baijiu.
3.2. The Discrepant Analysis of Microbial Community during Fermentation in Cold and Hot Seasons
Besides the concentration of lactic acid, the composition of the microbial community should be given extra attention because the excess lactic acid was produced by lactic acid bacteria from different sources. The Hiseq platform was selected to sequence the V3-V4 region of bacterial 16S rDNA in samples from cold and hot seasons. Data filtering, TAGS splicing, and OUT clustering were performed based on the original sequencing results. Sequences that matched with more than 97% similarity were classified as an OUT. Finally, the aggregation of fermented samples in the cold season was 4597 OTUs, which were divided into 4 phyla, 22 classes, 5 orders, 8 families, and 10 genera. The aggregation of fermented samples in the hot season was 6589 OTUs, which were divided into 5 phyla, 22 classes, 7 orders, 15 families, and 15 genera.
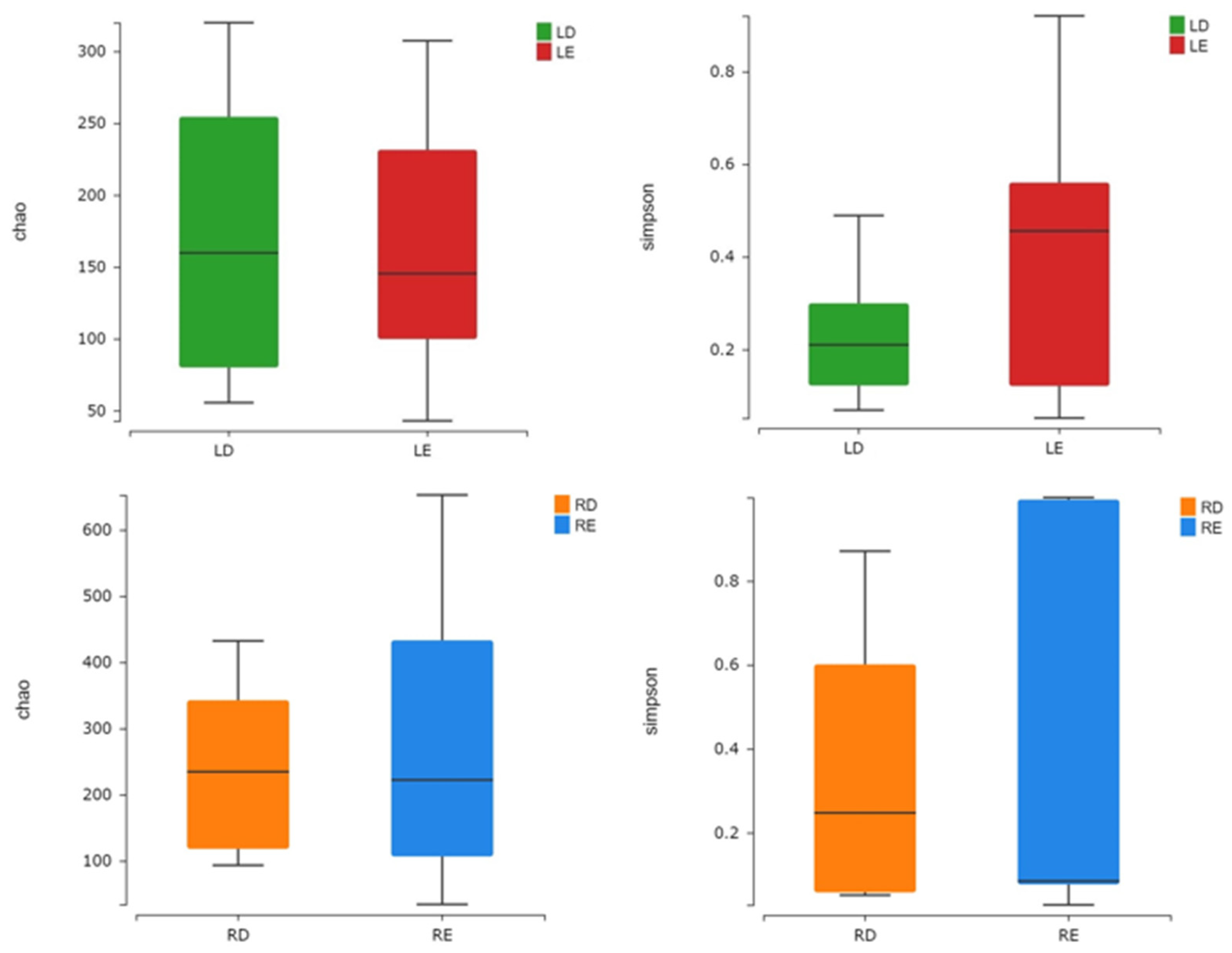
Based on these sequencing results, the index of the alpha diversity was analyzed to present the number of species during the fermentation and the richness and diversity of the microbial community in different seasons [
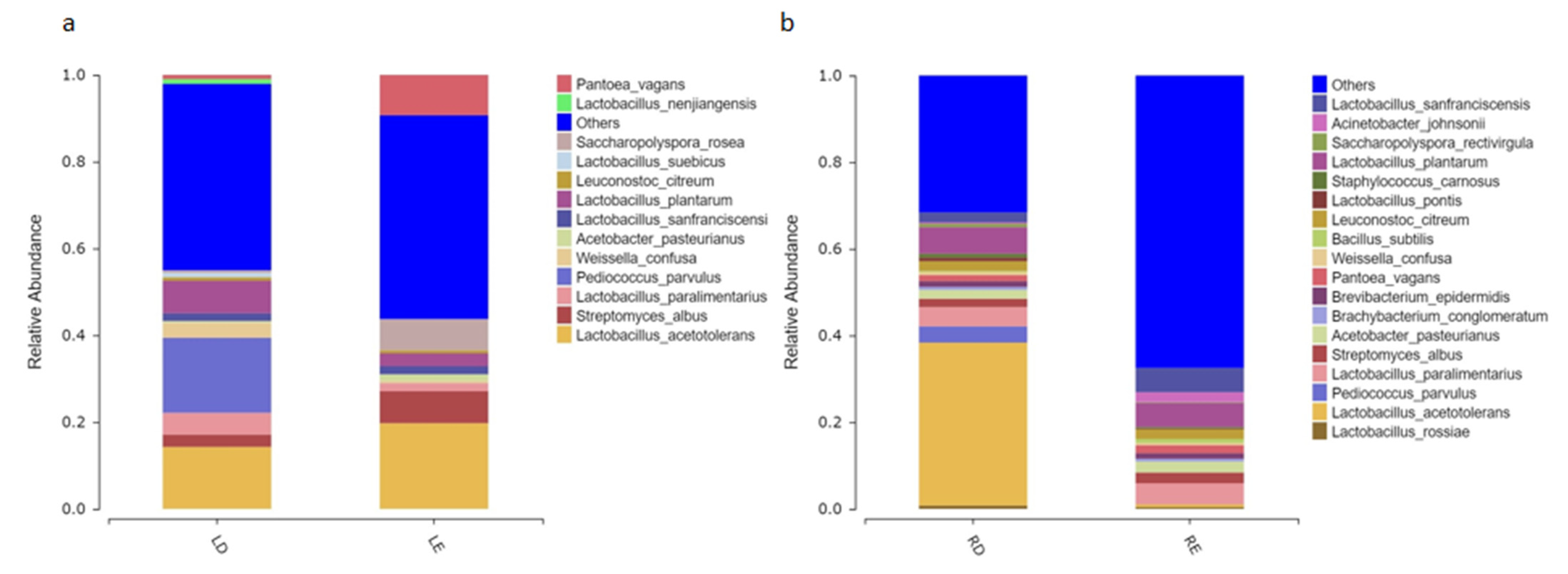
26]. The Chao index showed that LD (Dacha in the cold season) had a higher richness of microbial community than LE (Ercha in the cold season), while RD (Dacha in the hot season) had a lower richness of microbial community than RE (Ercha in the cold season). The Simpson index showed that the bacterial diversity of LE and RE was higher than that of LD and RD (
Figure 2). The results showed that the bacterial species in fermented grains became more diverse in both cold and hot seasons along with the progress of fermentation. In the latter stage of fermentation, the number of bacteria in fermented grains of the hot season gradually increased while the number of bacteria in fermented grains of the cold season significantly decreased.
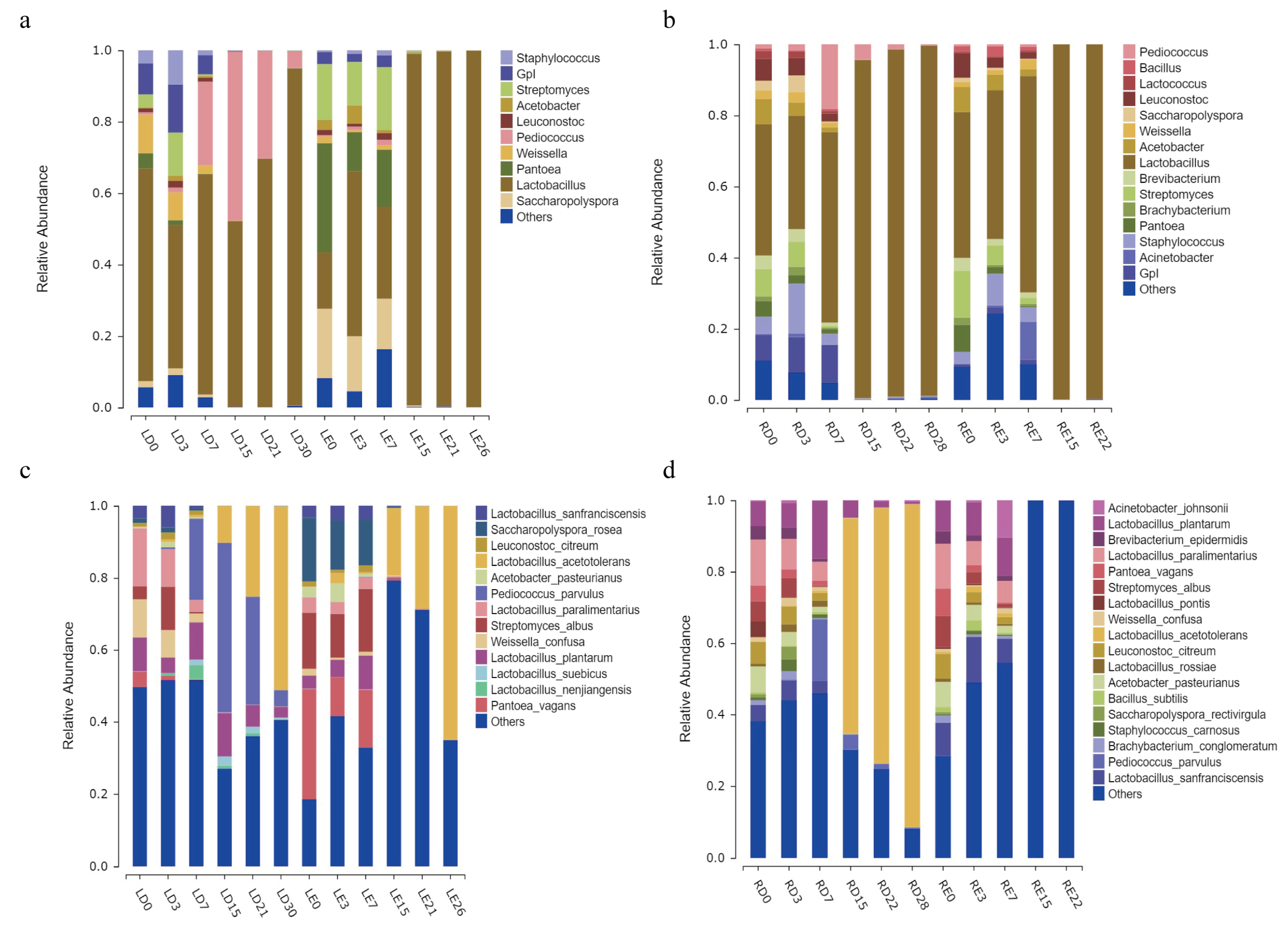
At the genus level (
Figure 3a,b),
Bacillus,
Lactococcus,
Brevibacterium,
Brachybacterium, and
Acinetobacter only existed in fermented grains in the hot season.
Lactobacillus is the dominant bacterium during the process of fermentation in both the cold and hot seasons, which is consistent with the previous results on the main genera in the fermentation of Fen-flavor Baijiu [
27]. Especially in the latter stage of fermentation, the relative abundance of
Lactobacillus in the hot season is significantly higher than that in the cold season, which almost reaches 100%. The previous studies also found that
Lactobacillus accounted for the largest proportion in the middle and late stages of fermentation [
28]. At the species level (
Figure 3c,d),
Lactobacillus acetotolerans was dominant in the latter stages of Dacha and Ercha in the cold season, and the trend became increasingly evident. However,
L. acetotolerans only existed in the latter stage of Dacha in the hot season, and its relative abundance reached 90.28% at the end of Dacha fermentation.
Pediococcus parvulus mainly existed from 7 days to the end of Dacha fermentation in both the cold and hot seasons, especially at the 15th day of Dacha in the cold season, accounting for 47.13%. However,
P. parvulus nearly disappeared during Ercha fermentation.
3.3. The Correlation Analysis between Lactic Acid and Bacteria Diversity in Cold and Hot Seasons
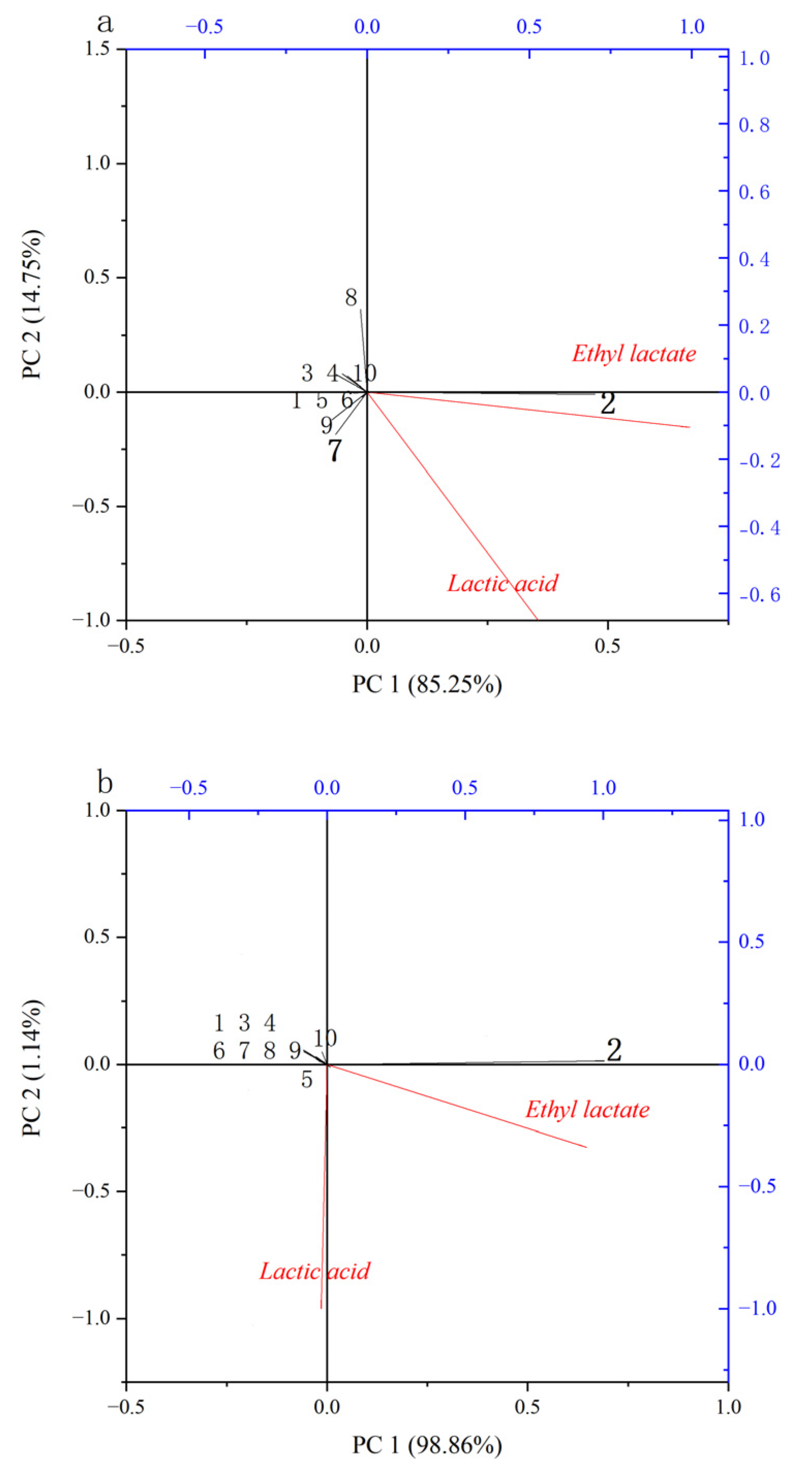
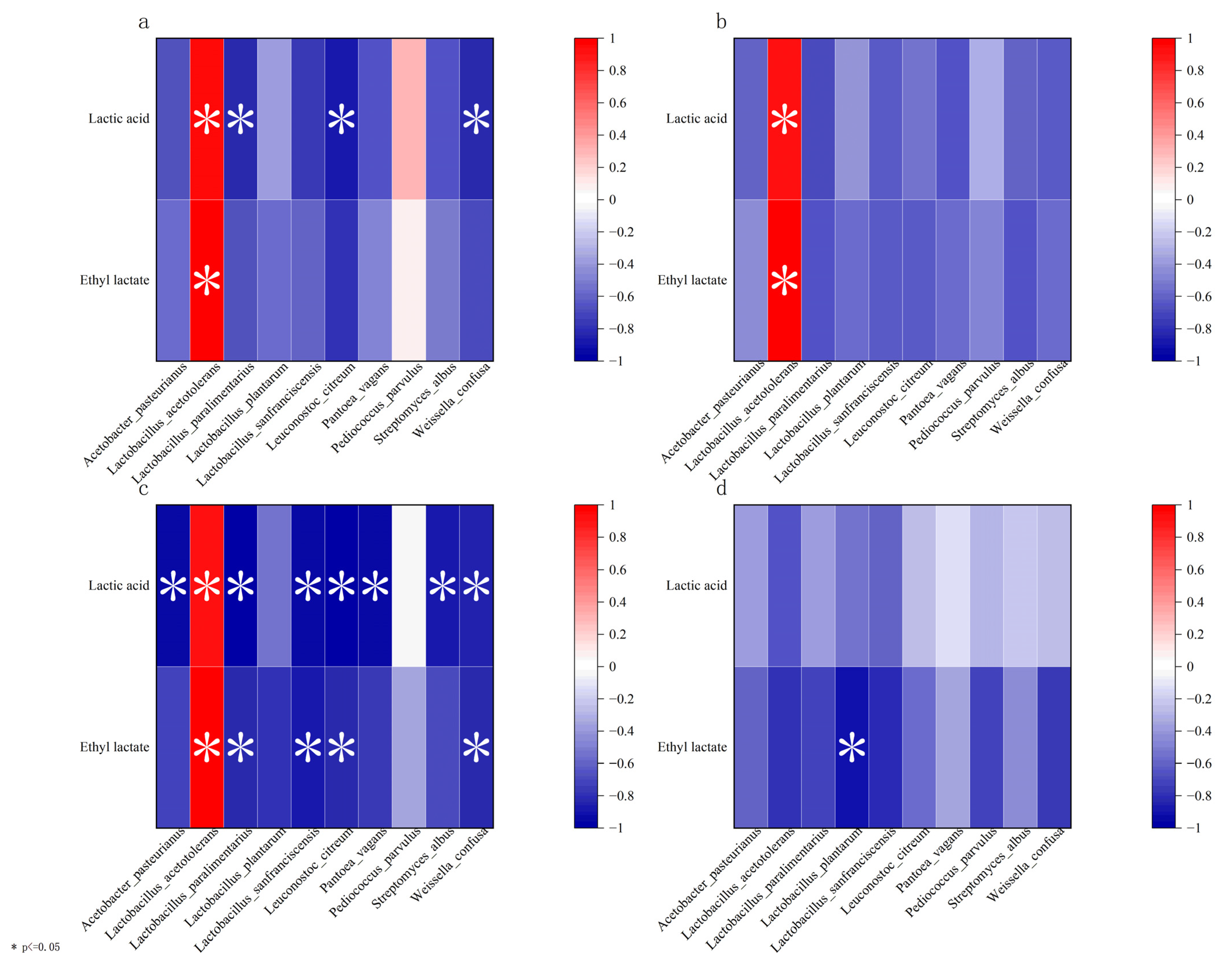
In order to further explore the key microorganisms affecting the concentration of lactic acid, RDA (redundancy analysis) and Pearson’s coefficient [
29] were used to analyze the relationship between the content of lactic acid/ethyl lactate and 10 bacteria with a high abundance in cold and hot seasons. As shown in
Figure 4a,
L. acetotolerans (2) has a positive correlation with lactic acid and ethyl lactate (the acute angle), while
Pantoea vagans (7) is positively correlated with lactic acid but negatively correlated with ethyl lactate (the obtuse angle) during fermentation in the cold season. The results in
Figure 4b showed that strains with high abundance were negatively correlated with lactic acid, whereas
L. acetotolerans (2) has a positive correlation with ethyl lactate. The previous research also showed that
L. acetotolerans was positively correlated with ethyl lactate, and the lack of
L. acetotolerans might cause a lag in fermentation and lower the content of ester [
9,
30]. These results suggest that
L. acetotolerans is the key microorganism that causes the different concentrations of lactic acid between cold and hot seasons.
During the Dacha fermentation in the cold season (
Figure 5a),
P. parvulus and
L. acetotolerans showed a positive correlation with lactic acid and ethyl lactate (
p ≤ 0.05).
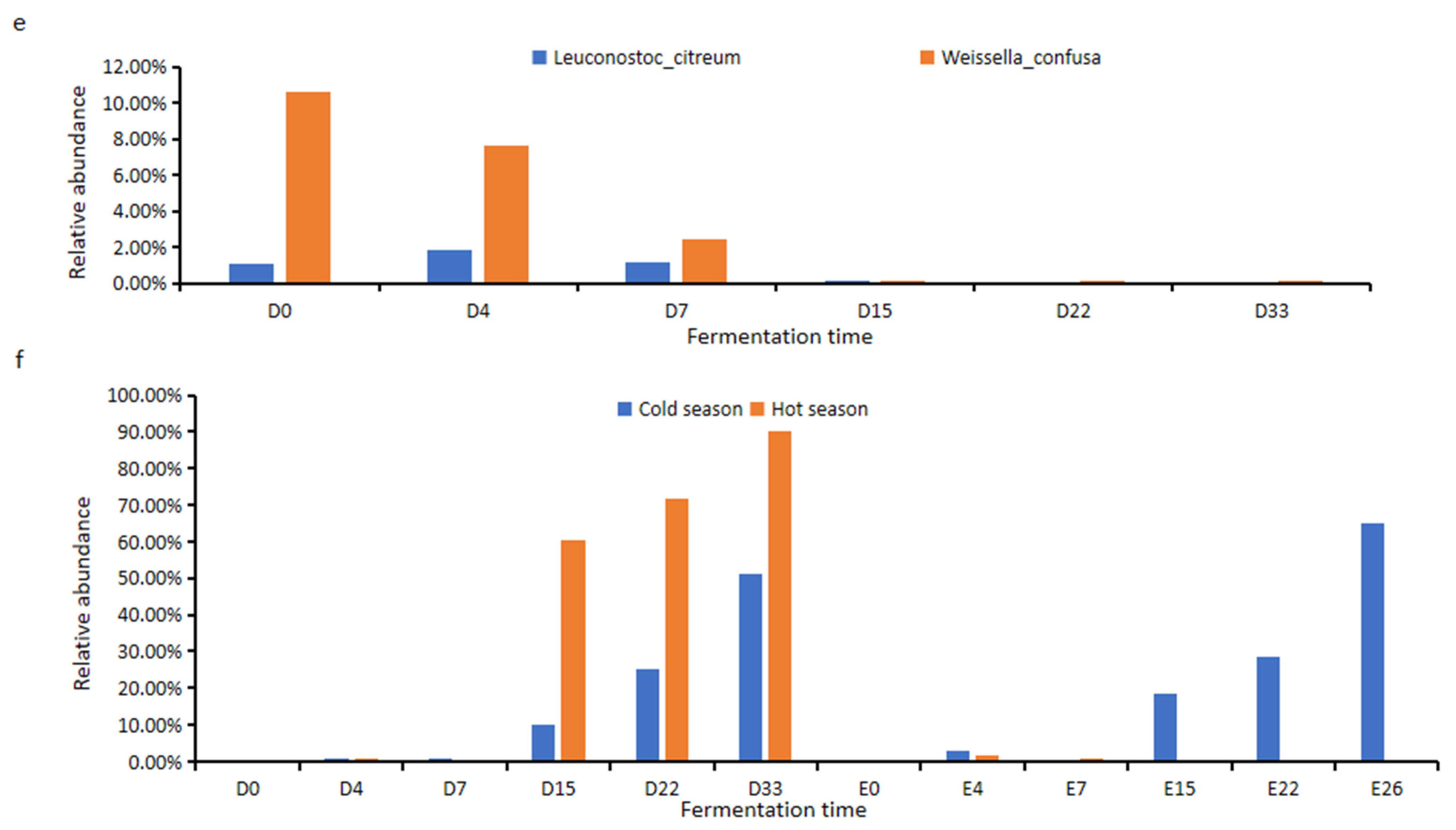
Leuconostoc citreum and
Weissella confusa were negatively correlated with their metabolites (ρ < −0.7) and the relative abundance of the two strains showed a decreasing trend during fermentation (
Figure 5e), indicating that the inhibition factors of acid production in fermented grains gradually decreased with the progress of fermentation. During the Ercha fermentation in the cold season (
Figure 5b), most of the strains showed a negative correlation with lactic acid and ethyl lactate (ρ < 0), while
L. acetotolerans showed a positive correlation with lactic acid and ethyl lactate (
p ≤ 0.05).
During the Dacha fermentation in the hot season (
Figure 5c),
L. acetotolerans was also positively correlated with lactic acid and ethyl lactate (
p ≤ 0.05), which was consistent with the results in the cold season but negatively correlated with lactic acid and ethyl lactate in other species. During the Ercha fermentation in the hot season (
Figure 5d), all species with high abundance showed a negative correlation with lactic acid and ethyl lactate (ρ < 0), especially for
Lactobacillus plantarum and ethyl lactate (ρ < −0.7).
L. plantarum showed an increasing trend during the Dacha fermentation in cold and hot seasons and during the Ercha fermentation in the cold season. However, the relative abundance of
L. plantarum significantly decreased during the Ercha fermentation in the hot season (
Figure 5f); it almost disappeared in the latter fermentation of Ercha in the hot season. At the same time,
L. plantarum was negatively correlated with the metabolites in this period (ρ < −0.5), which indicates that
L. plantarum is always a favorable factor for acid production in the fermentation of Fen-flavor Baijiu. Relevant studies also showed that
L. plantarum was the main strain producing ethyl lactate during the fermentation of other types of Baijiu [
31].
3.4. Isolation and Identification of Dominant Lactic-Acid-Producing Bacteria in Fermented Grains
In order to further identify the dominant lactic-acid-producing bacteria in the fermentation process of cold and hot seasons, MRS medium was used to isolate lactic-acid-producing microorganisms in the samples of fermented grains. As a result, 12 lactic-acid-producing microorganisms were isolated and identified:
Lacticaseibacillus paracasei LPC,
Lentilactobacillus hilgardii LH,
Lacticaseibacillus rhamnosus LR,
Lentilactobacillus parabuchneri LPB,
Lentilactobacillus buchneri LBC,
Weissella cibaria WCB,
Pediococcus pentosaceus PPP,
Latilactobacillus curvatus LCV,
Limosilactobacillus fermentum LF,
Weissella paramesenteroides WP,
W. confusa WCF, and
L. plantarum LP. In a previous study,
Lactobacillus brevis,
L. rhamnosus,
Lactobacillus casei, and
Enterococcus casseliflavus were isolated from fermented pit mud, and various lactic acid bacteria had different characteristics and effects on the fermentation of Baijiu [
32].
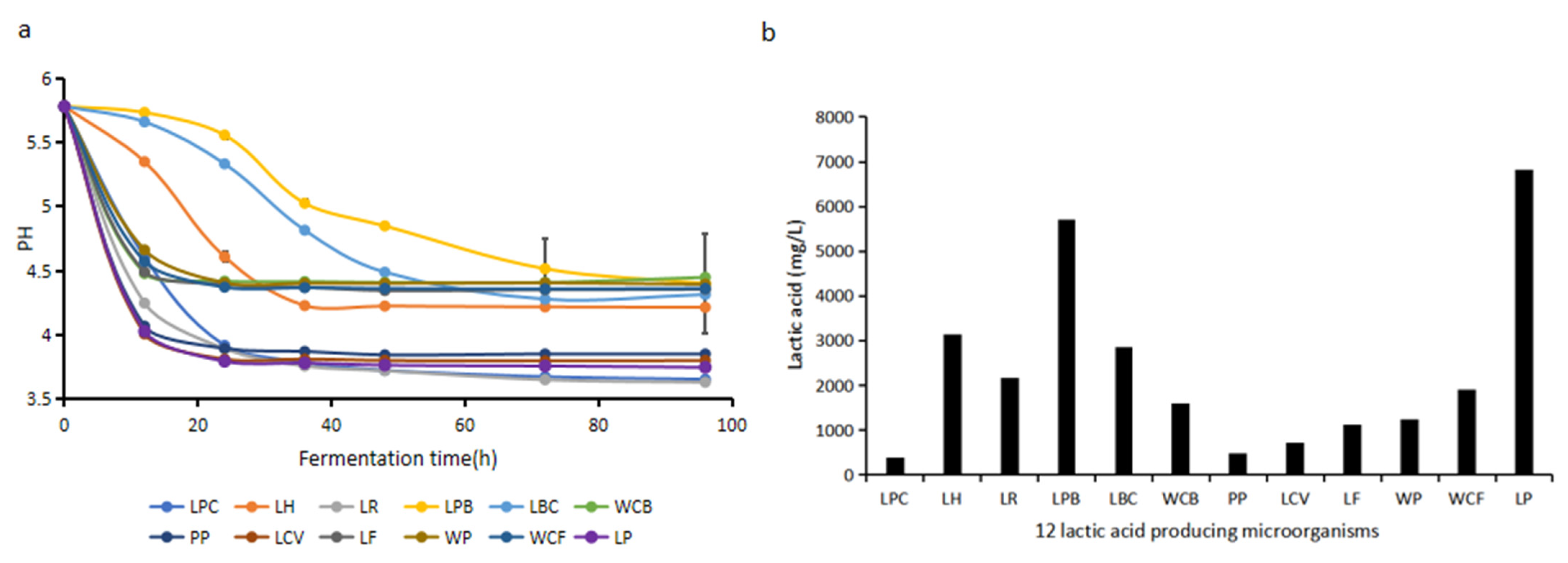
Among the 12 lactic-acid-producing microorganisms,
W. confusa WCF accounted for 3.5% during Dacha fermentation in the cold season, 0.57% during Ercha fermentation in the cold season, 0.73% during Dacha fermentation in the hot season, and 0.55% during Ercha fermentation in the hot season.
L. plantarum LP accounted for 7.52% during Dacha fermentation in the cold season, 2.97% during Ercha fermentation in the cold season, 6.12% during Dacha fermentation in the hot season, and 5.56% during Ercha fermentation in the hot season (
Figure 6). The other ten strains could not be found in
Figure 6 because the abundance of species is too low and some microbial information was missing during the extraction of the microbial genome. The high-abundance strains (
L. acetotolerans, et al.) in
Figure 6 could not be isolated mainly due to the requirement of a strict anaerobic environment for the culture of these strains.
3.5. The Capacity of Acid Production and the Resistance of Acid and Alcohol for the Dominant Lactic-Acid-Producing Microorganisms
The acid-producing capacities of 12 lactic-acid-producing microorganisms screened from fermented grains in cold and hot seasons were verified and analyzed. It can be seen from
Figure 7a that all the pH values in the fermentation broth dropped after the inoculation of 12 lactic-acid-producing microorganisms. The fastest decreasing rates of pH values were obtained from the LP and LCV strains. The pH values of fermentation broth for the two strains first decreased to 4.03 at 12 h and reached 3.75 at 24 h. The concentration of lactic acid produced by the LP strain was the highest at the end of fermentation, reaching 6.82 g/L (
Figure 7b). The pH values in the fermentation broth of LR and LPC strains dropped to the lowest level at the end of fermentation. The pH value of LR was as low as 3.63 when the fermentation reached the stable stage, and the concentration of lactic acid in the fermentation broth of LR was 2.17 g/L (
Figure 7b). Therefore, the LP strain has the strongest capacity for lactic acid production and the LR strain has the strongest capacity for acid production.
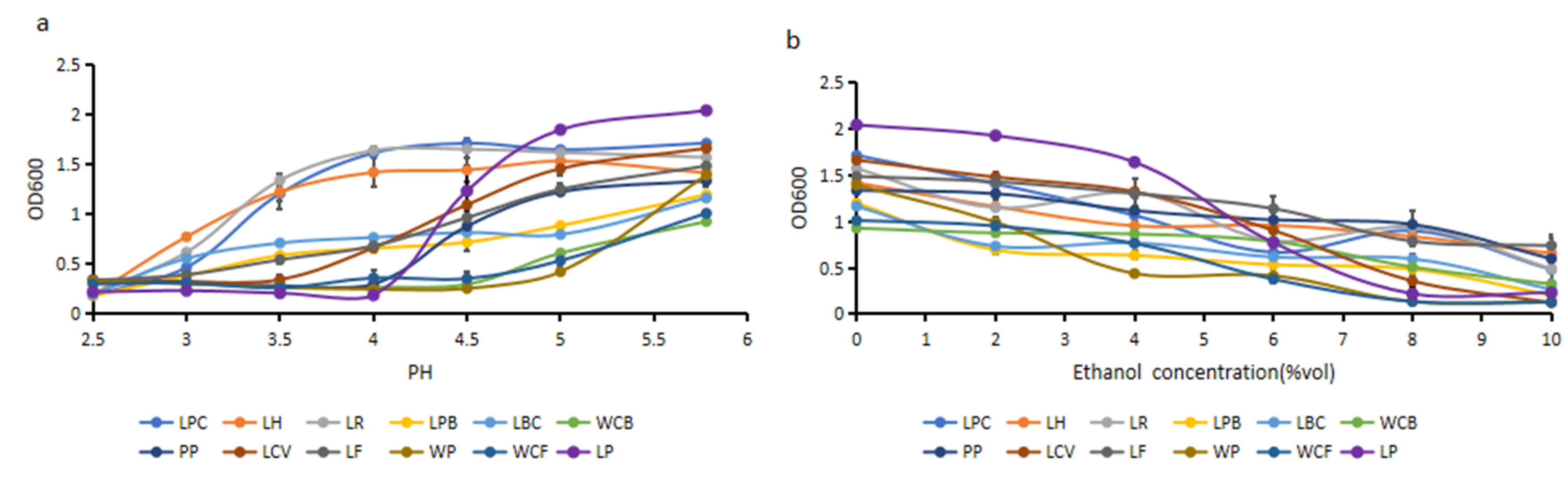
The acid and alcohol resistance of lactic-acid-producing microorganisms were verified and analyzed. As can be seen from
Figure 8a, the OD
600 values of LH, LR, and LPC remained stable at pH 4.0 and significantly decreased at pH 3.5. The OD
600 value of strain LH was as high as 0.77 at pH 3.0. As can be seen from
Figure 8b, the OD
600 value of WCB remained stable at a concentration of 6% ethanol. The OD
600 value of LH slightly decreased with the increasing concentration of ethanol, and the OD
600 value decreased from 1.42 to 0.66. Therefore, the LH strain has the strongest capacity of acid and alcohol resistance among 12 lactic-acid-producing microorganisms.
In this study, the changes in lactic acid and its derived esters, the composition of microbial community, and their correlation in cold and hot seasons were analyzed for Fen-flavor Baijiu. In addition, the key microorganisms (Lactobacillus_acetotolerans and Lactobacillus_plantarum) influencing the content of lactic acid in cold and hot seasons were identified. The physiological and biochemical properties of these microorganisms were also determined. These results pave the way for effectively controlling the content of lactic acid in summer, and the isolated strains have potential value for improving the flavor and taste of Fen-flavor Baijiu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








