
Engineering Escherichia coli for Efficient Aerobic Conversion of Glucose to Malic Acid through the Modified Oxidative TCA Cycle
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Media
2.2. Culturing of the Engineered Strains for Aerobic Production of Malic Acid
2.3. Analytical Techniques
3. Results and Discussion
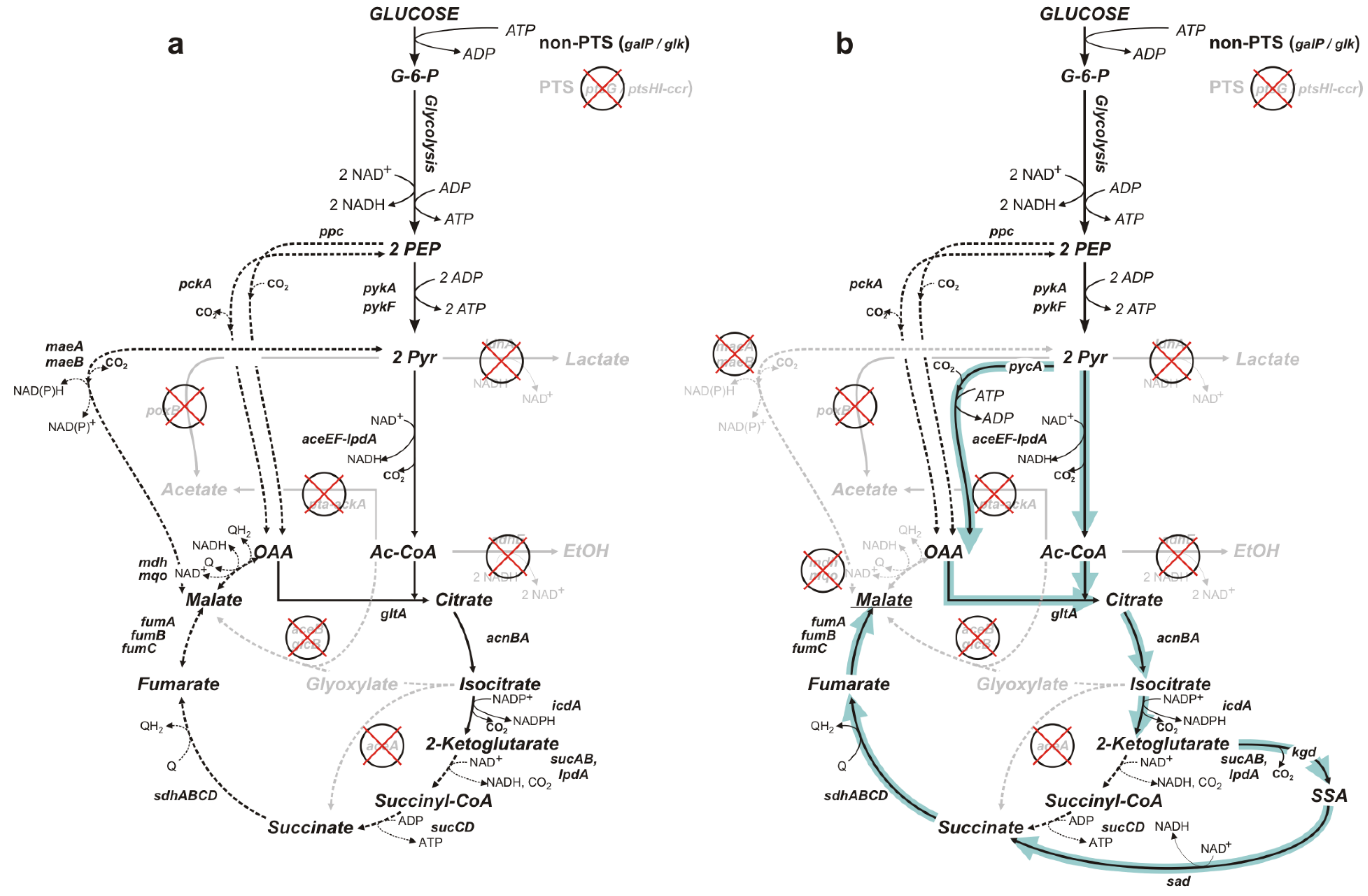
3.1. Construction and Evaluation of an Engineered E. coli Strain Capable of the Aerobic Production of Malic Acid from Glucose
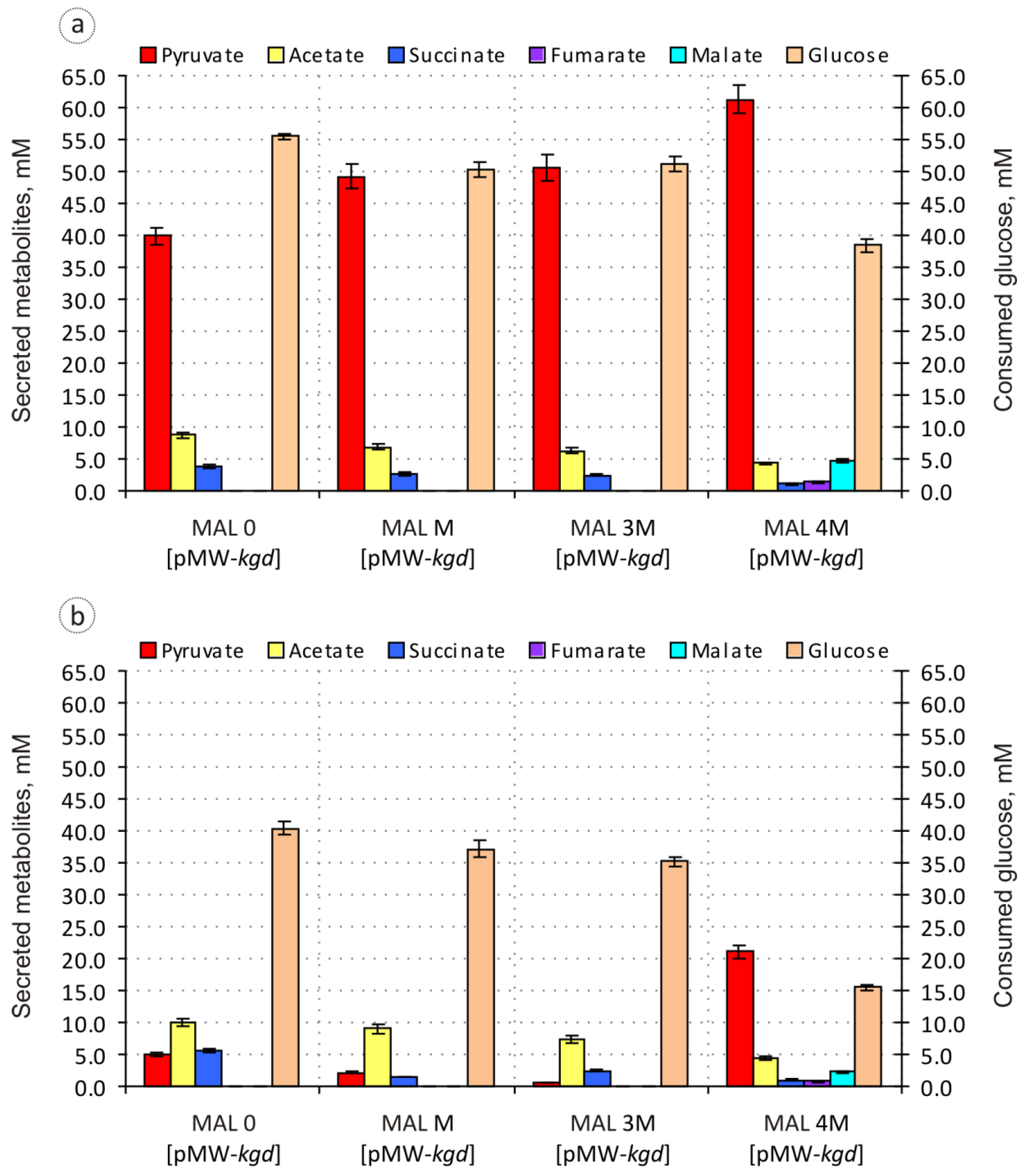
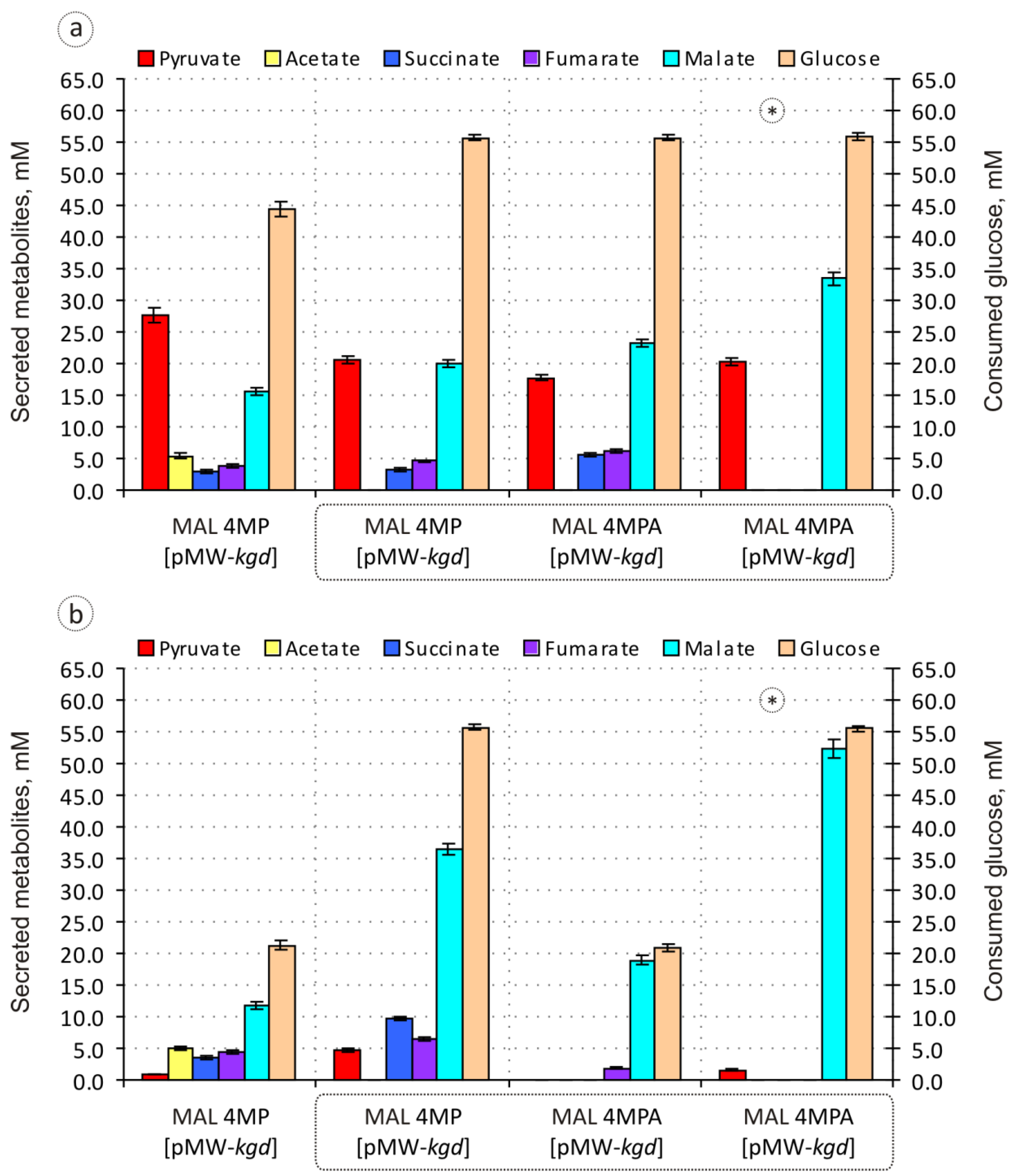
3.2. Improvement of the Engineered Malic Acid-Producing Strain and Evaluation of Its Biosynthetic Potential
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chi, Z.; Wang, Z.P.; Wang, G.Y.; Khan, I.; Chi, Z.M. Microbial biosynthesis and secretion of malic acid and its applications. Crit. Rev. Biotechnol. 2014, 36, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Thakker, C.; Martínez, I.; Li, W.; San, K.Y.; Bennett, G.N. Metabolic engineering of carbon and redox flow in the production of small organic acids. J. Ind. Microbiol. Biotechnol. 2015, 42, 403–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Xu, Y.; Xu, Q.; Cao, W.; Huang, H.; Liu, H. Microbial biosynthesis of L-malic acid and related metabolic engineering strategies: Advances and prospects. Front. Bioeng. Biotechnol. 2021, 9, 765685. [Google Scholar] [CrossRef] [PubMed]
- Kövilein, A.; Kubisch, C.; Cai, L.; Ochsenreither, K. Malic acid production from renewables: A review. J. Chem. Technol. Biotechnol. 2019, 95, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Zhang, J.; Chen, Y.; Xu, Q.; Song, P.; Li, Y.; Li, K.; Liu, H. Recent advances in microbial production of L-malic acid. Appl. Microbiol. Biotechnol. 2022, 106, 7973–7992. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass, Volume I: Results of Screening for Potential Candidates from Sugars and Synthesis Gas; US Department of Energy: Washington, DC, USA, 2004. [Google Scholar]
- Kim, J.S.; Baek, J.H.; Ryu, Y.B.; Hong, S.S.; Lee, M.S. Liquid hydrogenation of maleic anhydride with Pd/C catalyst at low pressure and temperature in batch reactor. J. Nanosci. Nanotechnol. 2015, 15, 290–294. [Google Scholar] [CrossRef]
- Peleg, Y.; Stieglitz, B.; Goldberg, I. Malic-acid accumulation by Aspergillus flavus. 1. Biochemical aspects of acid biosynthesis. Appl. Microbiol. Biotechnol. 1988, 28, 69–75. [Google Scholar] [CrossRef]
- Battat, E.; Peleg, Y.; Bercovitz, A.; Rokem, J.S.; Goldberg, I. Optimization of L-malic acid production by Aspergillus flavus in a stirred fermenter. Biotechnol. Bioeng. 1991, 37, 1108–1116. [Google Scholar] [CrossRef]
- Knuf, C.; Nookaew, I.; Remmers, I.; Khoomrung, S.; Brown, S.; Berry, A.; Nielsen, J. Physiological characterization of the high malic acid-producing Aspergillus oryzae strain 2103a–68. Appl. Microbiol. Biotechnol. 2014, 98, 3517–3527. [Google Scholar] [CrossRef]
- Ochsenreither, K.; Fischer, C.; Neumann, A.; Syldatk, C. Process characterization and influence of alternative carbon sources and carbon-to-nitrogen ratio on organic acid production by Aspergillus oryzae DSM1863. Appl. Microbiol. Biotechnol. 2014, 98, 5449–5460. [Google Scholar] [CrossRef]
- Schmitt, V.; Derenbach, L.; Ochsenreither, K. Enhanced L-malic acid production by Aspergillus oryzae DSM 1863 using repeated-batch cultivation. Front. Bioeng. Biotechnol. 2022, 9, 760500. [Google Scholar] [CrossRef] [PubMed]
- West, T.P. Malic acid production from thin stillage by Aspergillus species. Biotechnol. Lett. 2011, 33, 2463–2467. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.H.; Bashkirova, L.; Berka, R.; Chandler, T.; Doty, T.; McCall, K.; McCulloch, M.; McFarland, S.; Thompson, S.; Yaver, D.; et al. Metabolic engineering of Aspergillus oryzae NRRL 3488 for increased production of L-malic acid. Appl. Microbiol. Biotechnol. 2013, 97, 8903–8912. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shan, L.; Zhou, Y.; Xie, Z.; Ball, A.S.; Cao, W.; Liu, H. Development of a Cre-loxP-based genetic system in Aspergillus niger ATCC1015 and its application to construction of efficient organic acid-producing cell factories. Appl. Microbiol. Biotechnol. 2019, 103, 8105–8114. [Google Scholar] [CrossRef] [PubMed]
- Zelle, R.M.; de Hulster, E.; van Winden, W.A.; de Waard, P.; Dijkema, C.; Winkler, A.A.; Geertman, J.-M.A.; van Dijken, J.P.; Pronk, J.T.; van Maris, A.J. Malic acid production by Saccharomyces cerevisiae: Engineering of pyruvate carboxylation, oxaloacetate reduction, and malate export. Appl. Environ. Microbiol. 2008, 74, 2766–2777. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, Y.; Dong, X.; Hu, G.; Liu, L. Engineering rTCA pathway and C4-dicarboxylate transporter for L-malic acid production. Appl. Microbiol. Biotechnol. 2017, 101, 4041–4052. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Shanmugam, K.T.; Ingram, L.O. L-malate production by metabolically engineered Escherichia coli. Appl. Environ. Microb. 2011, 77, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Chen, X.; Qian, Y.; Wang, Y.; Wang, L.; Qiao, W.; Liu, L. Metabolic engineering of Escherichia coli W3110 to produce L-malate. Biotechnol. Bioeng. 2017, 114, 656–664. [Google Scholar] [CrossRef]
- Jiang, Y.; Zheng, T.; Ye, X.; Xin, F.; Zhang, W.; Dong, W.; Ma, J.; Jiang, M. Metabolic engineering of Escherichia coli for l-malate production anaerobically. Microb. Cell Fact. 2020, 19, 165. [Google Scholar] [CrossRef]
- Park, S.; Chang, K.S.; Jin, E.; Pack, S.P.; Lee, J. Oxaloacetate and malate production in engineered Escherichia coli by expression of codon-optimized phosphoenolpyruvate carboxylase2 gene from Dunaliella salina. Bioprocess. Biosyst. Eng. 2013, 36, 127–131. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, F.; Zhang, C.; Hu, G.; Gao, C.; Chen, X.; Liu, L. Enhancement of malate production through engineering of the periplasmic rTCA pathway in Escherichia coli. Biotechnol. Bioeng. 2018, 115, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Li, Z.; Ma, D.; Ye, C.; Zhang, L.; Gao, C.; Liu, L.; Chen, X. Light-driven CO2 sequestration in Escherichia coli to achieve theoretical yield of chemicals. Nat. Catal. 2021, 4, 395–406. [Google Scholar] [CrossRef]
- Khandelwal, R.; Srivastava, P.; Bisaria, V.S. Expression of Escherichia coli malic enzyme gene in Zymomonas mobilis for production of malic acid. J. Biotechnol. 2022, 351, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Trichez, D.; Auriol, C.; Baylac, A.; Irague, R.; Dressaire, C.; Carnicer-Heras, M.; Heux, S.; François, J.M. Engineering of Escherichia coli for Krebs cycle-dependent production of malic acid. Microb. Cell Fact. 2018, 17, 113. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Chao, G.; Gunsalus, R.P. Aerobic regulation of the sucABCD genes of Escherichia coli, which encode alpha-ketoglutarate dehydrogenase and succinyl coenzyme A synthetase: Roles of ArcA, Fnr, and the upstream sdhCDAB promoter. J. Bacteriol. 1997, 179, 4138–4142. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Wang, S.; Hu, G.; Guo, L.; Chen, X.; Xu, P.; Liu, L. Engineering Escherichia coli for malate production by integrating modular pathway characterization with CRISPRi-guided multiplexed metabolic tuning. Biotechnol Bioeng. 2018, 115, 661–672. [Google Scholar] [CrossRef]
- Skorokhodova, A.Y.; Stasenko, A.A.; Gulevich, A.Y.; Debabov, V.G. Construction of a synthetic bypass for improvement of aerobic synthesis of succinic acid through the oxidative branch of the tricarboxylic acid cycle by recombinant Escherichia coli strains. Appl. Biochem. Microbiol. 2018, 54, 245–251. [Google Scholar] [CrossRef]
- Skorokhodova, A.Y.; Gulevich, A.Y.; Debabov, V.G. Engineering Escherichia coli for efficient aerobic conversion of glucose to fumaric acid. Biotechnol. Rep. 2022, 33, e00703. [Google Scholar] [CrossRef]
- Tian, J.; Bryk, R.; Itoh, M.; Suematsu, M.; Nathan, C. Variant tricarboxylic acid cycle in Mycobacterium tuberculosis: Identification of alpha-ketoglutarate decarboxylase. Proc. Natl. Acad. Sci. USA 2005, 102, 10670–10675. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Bryant, D.A. The tricarboxylic acid cycle in cyanobacteria. Science 2011, 334, 1551–1553. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katashkina, J.I.; Skorokhodova, A.Y.; Zimenkov, D.V.; Gulevich, A.Y.; Minaeva, N.I.; Doroshenko, V.G.; Biryukova, I.Y.; Mashko, S.V. Tuning the expression level of a gene located on a bacterial chromosome. Mol. Biol. 2005, 39, 719–726. [Google Scholar] [CrossRef]
- Gulevich, A.Y.; Skorokhodova, A.Y.; Ermishev, V.Y.; Krylov, A.A.; Minaeva, N.I.; Polonskaya, Z.M.; Zimenkov, D.V.; Biryukova, I.V.; Mashko, S.V. A new method for the construction of translationally coupled operons in a bacterial chromosome. Mol. Biol. 2009, 43, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Gulevich, A.Y.; Skonechny, M.S.; Sukhozhenko, A.V.; Skorokhodova, A.Y.; Debabov, V.G. Study on aerobic biosynthesis of 4-hydroxybutyric acid by Escherichia coli cells upon heterologous expression of the 2-ketoglutarate decarboxylase gene. Appl. Biochem. Microbiol. 2015, 51, 804–811. [Google Scholar] [CrossRef]
- Fuhrer, T.; Chen, L.; Sauer, U.; Vitkup, D. Computational prediction and experimental verification of the gene encoding the NAD+/NADP+-dependent succinate semialdehyde dehydrogenase in Escherichia coli. J. Bacteriol. 2007, 189, 8073–8078. [Google Scholar] [CrossRef] [Green Version]
- De Mey, M.; Lequeux, G.J.; Beauprez, J.J.; Maertens, J.; Van Horen, E.; Soetaert, W.K.; Vanrolleghem, P.A.; Vandamme, E.J. Comparison of different strategies to reduce acetate formation in Escherichia coli. Biotechnol. Prog. 2007, 23, 1053–1063. [Google Scholar] [CrossRef]
- Veit, A.; Polen, T.; Wendisch, V.F. Global gene expression analysis of glucose overflow metabolism in Escherichia coli and reduction of aerobic acetate formation. Appl. Microbiol. Biotechnol. 2007, 74, 406–421. [Google Scholar] [CrossRef]
- Sauer, U.; Eikmanns, B.J. The PEP-pyruvate-oxaloacetate node as the switch point for carbon flux distribution in bacteria. FEMS Microbiol. Rev. 2005, 29, 765–794. [Google Scholar] [CrossRef] [Green Version]
- van der Rest, M.E.; Frank, C.; Molenaar, D. Functions of the membrane-associated and cytoplasmic malate dehydrogenases in the citric acid cycle of Escherichia coli. J. Bacteriol. 2000, 182, 6892–6899. [Google Scholar] [CrossRef] [Green Version]
- Khankal, R.; Chin, J.W.; Ghosh, D.; Cirino, P.C. Transcriptional effects of CRP* expression in Escherichia coli. J. Biol. Eng. 2009, 3, 13. [Google Scholar] [CrossRef]
- Park, S.J.; Cotter, P.A.; Gunsalus, R.P. Regulation of malate dehydrogenase (mdh) gene expression in Escherichia coli in response to oxygen, carbon, and heme availability. J. Bacteriol. 1995, 177, 6652–6656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bologna, F.P.; Andreo, C.S.; Drincovich, M.F. Escherichia coli malic enzymes: Two isoforms with substantial differences in kinetic properties, metabolic regulation, and structure. J. Bacteriol. 2007, 189, 5937–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, D.E.; Shin, S.; Rhee, J.S.; Pan, J.G. Acetate metabolism in a pta mutant of Escherichia coli W3110: Importance of maintaining acetyl coenzyme A flux for growth and survival. J. Bacteriol. 1999, 181, 6656–6663. [Google Scholar] [CrossRef] [Green Version]
- Baev, M.V.; Baev, D.; Radek, A.J.; Campbell, J.W. Growth of Escherichia coli MG1655 on LB medium: Monitoring utilization of amino acids, peptides, and nucleotides with transcriptional microarrays. Appl. Microbiol. Biotechnol. 2006, 71, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Theodosiou, E.; Frick, O.; Bühler, B.; Schmid, A. Metabolic network capacity of Escherichia coli for Krebs cycle-dependent proline hydroxylation. Microb. Cell Fact. 2015, 14, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodosiou, E.; Breisch, M.; Julsing, M.K.; Falcioni, F.; Bühler, B.; Schmid, A. An artificial TCA cycle selects for efficient α-ketoglutarate dependent hydroxylase catalysis in engineered Escherichia coli. Biotechnol. Bioeng. 2017, 114, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Burgard, A.; Burk, M.J.; Osterhout, R.; Van Dien, S.; Yim, H. Development of a commercial scale process for production of 1,4-butanediol from sugar. Curr. Opin. Biotechnol. 2016, 42, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J. The acetate switch. Microbiol. Mol. Biol. Rev. 2005, 69, 12–50. [Google Scholar] [CrossRef] [Green Version]
- Burstein, C.; Tiankova, L.; Kepes, A. Respiratory control in Escherichia coli K 12. Eur. J. Biochem. 1979, 94, 387–392. [Google Scholar] [CrossRef]
- Causey, T.B.; Shanmugam, K.T.; Yomano, L.P.; Ingram, L.O. Engineering Escherichia coli for efficient conversion of glucose to pyruvate. Proc. Natl. Acad. Sci. USA 2004, 101, 2235–2240. [Google Scholar] [CrossRef]
- Zhu, Y.; Eiteman, M.A.; Altman, R.; Altman, E. High glycolytic flux improves pyruvate production by a metabolically engineered Escherichia coli strain. Appl. Environ. Microbiol. 2008, 74, 6649–6655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Causey, T.B.; Zhou, S.; Shanmugam, K.T.; Ingram, L.O. Engineering the metabolism of Escherichia coli W3110 for the conversion of sugar to redox-neutral and oxidized products: Homoacetate production. Proc. Natl. Acad. Sci. USA 2003, 100, 825–832. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strain/Plasmid | Genotype | Reference |
|---|---|---|
| E. coli strains | ||
| MG1655 | Wild-type E. coli K-12 strain VKPM B-6195 | VKPM |
| MAL 0 | MG1655 ∆ackA-pta, ∆poxB, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB | [28] |
| MAL M | MG1655 ∆ackA-pta, ∆poxB, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB, ∆mdh | This study |
| MAL 3M | MG1655 ∆ackA-pta, ∆poxB, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB, ∆mdh, ∆maeA, ∆maeB | This study |
| MAL 4M | MG1655 ∆ackA-pta, ∆poxB, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB, ∆mdh, ∆maeA, ∆maeB, ∆mqo | This study |
| MAL 4MP | MG1655 ∆ackA-pta, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB, ∆mdh, ∆maeA, ∆maeB, ∆mqo; poxB::PL-pycABs | This study |
| MAL 4MPA | MG1655 ∆ackA-pta, ∆ldhA, ∆adhE, ∆ptsG, PL-glk, Ptac-galP, ∆aceBAK, ∆glcB, ∆mdh, ∆maeA, ∆maeB, ∆mqo; poxB::PL-pycABs; ∆atpFH | This study |
| Plasmids | ||
| pKD46 | pINT-ts, bla, ParaB-λgam-bet-exo | [32] |
| pMW118-(λattL-Cm-λattR) | pSC101, bla, cat, λattL-cat-λattR cassette | [33] |
| pMWts-Int/Xis | pSC101-ts, bla, PR-λxis-int, cIts857 | [34] |
| pMW-kgd | pMW119 with cloned M. tuberculosis 2-ketoglutarate decarboxylase gene (kgd) | [35] |
| Strain | IPTG | Pyruvate | Acetate | Succinate | Fumarate | Malate | % C-Recovery a | DCW (g/L) |
|---|---|---|---|---|---|---|---|---|
| MAL 0 [pMW-kgd] | − | 71.9 ± 1.6 | 15.7 ± 0.4 | 7.0 ± 0.2 | n.d. | n.d. | 46 | 4.18 ± 0.12 |
| + | 12.5 ± 0.3 | 24.8 ± 1.1 | 13.6 ± 0.6 | n.d. | n.d. | 24 | 4.90 ± 0.15 | |
| MAL M [pMW-kgd] | − | 98.0 ± 1.7 | 13.7 ± 0.5 | 5.2 ± 0.4 | n.d. | n.d. | 59 | 4.16 ± 0.10 |
| + | 5.8 ± 0.3 | 24.2 ± 0.9 | 4.0 ± 0.3 | n.d. | n.d. | 14 | 4.82 ± 0.16 | |
| MAL 3M [pMW-kgd] | − | 98.7 ± 1.5 | 12.0 ± 0.4 | 4.9 ± 0.4 | n.d. | n.d. | 58 | 4.20 ± 0.12 |
| + | 1.5 ± 0.1 | 20.7 ± 1.0 | 7.1 ± 0.6 | n.d. | n.d. | 13 | 4.96 ± 0.15 | |
| MAL 4M [pMW-kgd] | − | 159.6 ± 2.1 | 11.2 ± 0.5 | 2.9 ± 0.2 | 3.6 ± 0.2 | 12.5 ± 0.4 | 96 | 2.75 ± 0.07 |
| + | 135.9 ± 1.8 | 29.0 ± 1.2 | 6.6 ± 0.5 | 5.2 ± 0.2 | 14.9 ± 0.5 | 95 | 2.27 ± 0.06 | |
| MAL 4MP [pMW-kgd] | − | 62.3 ± 1.0 | 12.1 ± 0.6 | 6.5 ± 0.4 | 8.8 ± 0.3 | 35.1 ± 0.6 | 69 | 2.72 ± 0.08 |
| + | 3.8 ± 0.2 | 23.5 ± 1.2 | 16.9 ± 0.7 | 20.7 ± 0.8 | 55.9 ± 0.9 | 72 | 1.74 ± 0.04 |
| Strain | IPTG | Pyruvate | Acetate | Succinate | Fumarate | Malate | % C-Recovery a | DCW (g/L) |
|---|---|---|---|---|---|---|---|---|
| MAL 4MP [pMW-kgd] | − | 36.9 ± 0.7 | n.d. | 5.9 ± 0.4 | 8.3 ± 0.3 | 35.7 ± 0.7 | 52 | 2.74 ± 0.08 |
| + | 8.6 ± 0.4 | n.d. | 17.2 ± 0.7 | 11.5 ± 0.5 | 65.5 ± 1.2 | 67 | 2.76 ± 0.09 | |
| MAL 4MPA [pMW-kgd] | − | 31.8 ± 0.6 | n.d. | 10.1 ± 0.5 | 11.1 ± 0.6 | 41.8 ± 0.7 | 58 | 1.32 ± 0.03 |
| + | n.d. | n.d. | n.d. | 9.1 ± 0.2 | 90.4 ± 1.5 | 67 | 0.58 ± 0.02 |
| Strain | IPTG | Pyruvate | Acetate | Succinate | Fumarate | Malate | % C-Recovery a | DCW (g/L) |
|---|---|---|---|---|---|---|---|---|
| MAL 4MPA [pMW-kgd] | − | 36.2 ± 0.8 | n.d. | n.d. | n.d. | 59.9 ± 1.3 | 58 | 2.56 ± 0.07 |
| + | 2.8 ± 0.1 | n.d. | n.d. | n.d. | 93.7 ± 1.8 | 64 | 2.58 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skorokhodova, A.Y.; Stasenko, A.A.; Krasilnikova, N.V.; Gulevich, A.Y.; Debabov, V.G. Engineering Escherichia coli for Efficient Aerobic Conversion of Glucose to Malic Acid through the Modified Oxidative TCA Cycle. Fermentation 2022, 8, 738. https://doi.org/10.3390/fermentation8120738
Skorokhodova AY, Stasenko AA, Krasilnikova NV, Gulevich AY, Debabov VG. Engineering Escherichia coli for Efficient Aerobic Conversion of Glucose to Malic Acid through the Modified Oxidative TCA Cycle. Fermentation. 2022; 8(12):738. https://doi.org/10.3390/fermentation8120738
Chicago/Turabian StyleSkorokhodova, Alexandra Yu., Anastasiya A. Stasenko, Natalya V. Krasilnikova, Andrey Yu. Gulevich, and Vladimir G. Debabov. 2022. "Engineering Escherichia coli for Efficient Aerobic Conversion of Glucose to Malic Acid through the Modified Oxidative TCA Cycle" Fermentation 8, no. 12: 738. https://doi.org/10.3390/fermentation8120738