
Effects of Starter Cultures and Type of Casings on the Microbial Features and Volatile Profile of Fermented Sausages
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sausage Manufacture
2.2. Starter Cultures
2.3. Physico-Chemical Parameters
2.4. Microbial Counts
2.5. Aroma Profile Analysis
2.6. Sensory Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical Parameters
3.2. Microbial Counts
3.3. Flavour Formation
3.4. Sensory Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suurs, P.; Barbut, S. Collagen use for co-extruded sausage casings—A review. Trends Food Sci. Technol. 2020, 102, 91–101. [Google Scholar] [CrossRef]
- Feiner, G. Meat Products Handbook. Practical Science and Technology, 1st ed.; Woodhead Publishing: Cambridge, UK, 2006. [Google Scholar]
- Niinivaara, F.P. Starter cultures in the processing of meat by fermentation and dehydration. In Proceedings of the 44th Annual Reciprocal Meat Conference, Kansas State University, Manhattan, KS, USA, 9–12 June 1991. [Google Scholar]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of starter cultures on the safety of fermented meat products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.; Ferreira, V.; Magalhães, R.; Teixeira, P. Biocontrol strategies for Mediterranean-style fermented sausages. Food Res. Int. 2018, 103, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Cocconcelli, P.S.; Fontana, C. Bacteria. In Handbook of Fermented Meat and Poultry; Toldrá, F., Hui, Y.H., Astiasarán, I., Sebranek, J.G., Talon, R., Eds.; Wiley Blackwell: Ames, IA, USA, 2014; pp. 117–128. [Google Scholar]
- Berni, E. Mold. In Handbook of Fermented Meat and Poultry; Toldrá, F., Hui, Y.H., Astiasarán, I., Sebranek, J.G., Talon, R., Eds.; Wiley Blackwell: Ames, IA, USA, 2014; pp. 147–153. [Google Scholar]
- Carballo, J. The role of fermentation reactions in the generation of flavor and aroma of foods. In Fermentation, Effects on Food Properties; Mehta, B.M., Kamal-Eldin, A., Iwanski, R.Z., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 51–83. [Google Scholar]
- Harper, B.A.; Barbut, S.; Lim, L.T.; Marcone, M.F. Microstructural and textural investigation of various manufactured collagen sausage casings. Food Res. Int. 2012, 49, 494–500. [Google Scholar] [CrossRef]
- Savic, Z.; Savic, I. Sausage Casings, 2nd ed.; Victus International GmbH: Wien, Austria, 2016. [Google Scholar]
- Ratanavaraporn, J.; Kanokpanont, S.; Tabata, Y.; Damrongsakkul, S. Effects of acid type on physical and biological properties of collagen scaffolds. J. Biomater. Sci. 2008, 19, 945–952. [Google Scholar] [CrossRef]
- Bedia, M.; Méndez, L.; Bañón, S. Evaluation of different starter cultures (Staphylococci plus Lactic Acid Bacteria) in semi-ripened Salami stuffed in swine gut. Meat Sci. 2011, 87, 381–386. [Google Scholar] [CrossRef]
- Franciosa, L.; Alessandria, V.; Dolci, P.; Rantsiou, K.; Cocolin, L. Sausage fermentation and starter cultures in the era of molecular biology methods. Int. J. Food Microbiol. 2018, 279, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Montanari, C.; Gatto, V.; Torriani, S.; Barbieri, F.; Bargossi, E.; Lanciotti, R.; Grazia, L.; Magnani, R.; Tabanelli, G.; Gardini, F. Effects of the diameter on physicochemical, microbiological and volatile profile in dry fermented sausages produced with two different starter cultures. Food Biosci. 2018, 22, 9–18. [Google Scholar] [CrossRef]
- Pasini, F.; Soglia, F.; Petracci, M.; Caboni, M.F.; Marziali, S.; Montanari, C.; Gardini, F.; Grazia, L.; Tabanelli, G. Effect of fermentation with different lactic acid bacteria starter cultures on biogenic amine content and ripening patterns in dry fermented sausages. Nutrients 2018, 10, 1497. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Marino, R.A.; Della Malva, A.; Sevi, A.; Del Nobile, M.A. Influence of different casings on salami produced with meat from buffalo and Podolian cattle. J. Food Qual. 2012, 35, 127–136. [Google Scholar] [CrossRef]
- Zając, M.; Pająk, P.; Skowyra, G. Characterization of edible collagen casings in comparison with the ovine casing and their effect on sausage quality. J. Sci. Food Agric. 2021, 101, 6001–6009. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, L.; Zhang, Y.; Han, W.; Duan, Y. Effect of collagen casing on the quality characteristics of fermented sausage. PLoS ONE 2022, 17, e0263389. [Google Scholar] [CrossRef]
- Montanari, C.; Bargossi, E.; Gardini, A.; Lanciotti, R.; Magnani, R.; Gardini, F.; Tabanelli, G. Correlation between volatile profiles of Italian fermented sausages and their size and starter culture. Food Chem. 2016, 192, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Coloretti, F.; Tabanelli, G.; Chiavari, C.; Lanciotti, R.; Grazia, L.; Gardini, F.; Montaneri, C. Effect of wine addition on microbiological characteristics, volatile molecule profiles and biogenic amine contents in fermented sausages. Meat Sci. 2014, 96, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Wien, Austria, 2020. [Google Scholar]
- Cevoli, C.; Fabbri, A.; Tabanelli, G.; Montanari, C.; Gardini, F.; Lanciotti, R.; Guarnieri, A. Finite element model of salami ripening process and successive storage in package. J. Food Eng. 2014, 132, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Tabanelli, G.; Coloretti, F.; Chiavari, C.; Grazia, L.; Lanciotti, R.; Gardini, F. Effects of starter cultures and fermentation climate on the properties of two types of typical Italian dry fermented sausages produced under industrial conditions. Food Control 2012, 26, 416–426. [Google Scholar] [CrossRef]
- Barbieri, F.; Laghi, L.; Montanari, C.; Lan, Q.; Levante, A.; Gardini, F.; Tabanelli, G. Insights into the metabolomic diversity of Latilactobacillus sakei. Foods 2022, 11, 477. [Google Scholar] [CrossRef]
- Rimaux, T.; Rivière, A.; Illeghems, K.; Weckx, S.; Vuyst, L.D.; Leroy, F. Expression of the arginine deiminase pathway genes in Lactobacillus sakei is strain dependent and is affected by the environmental pH. Appl. Environ. Microbiol. 2012, 78, 4874–4883. [Google Scholar] [CrossRef] [Green Version]
- Champomier-Vergés, M.C.; Zuñiga, M.; Morel-Deville, F.; Perez-Martinez, G.; Zagorec, M. Relationship between arginine degradation, pH and survival in Lactobacillus sakei. FEMS Microbiol. Lett. 1999, 180, 297–304. [Google Scholar] [CrossRef]
- Smid, E.J.; Kleerebezem, M. Production of aroma compounds in lactic fermentations. Annu. Rev. Food Sci. Technol. 2014, 5, 313–326. [Google Scholar] [CrossRef]
- Flores, M.; Corral, S.; Cano-García, L.; Salvador, A.; Belloch, C. Yeast strains as potential aroma enhancers in dry fermented sausages. Int. J. Food Microbiol. 2015, 212, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Gardini, F.; Suzzi, G.; Lombardi, A.; Galgano, F.; Crudele, M.A.; Andrighetto, C.; Schirone, M.; Tofalo, R. A survey of yeasts in traditional sausages of southern Italy. FEMS Yeast Res. 2001, 1, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, R.C.S.; Gouvêa, D.M.; Hungaro, H.M.; Sodré, A.F.F.; Querol-Simon, A. Dynamics of the yeast flora in artisanal country style and industrial dry cured sausage (yeast in fermented sausage). Food Control 2013, 29, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Tabanelli, G.; Bargossi, E.; Gardini, A.; Lanciotti, R.; Magnani, R.; Gardini, F.; Montanari, C. Physico-chemical and microbiological characterisation of Italian fermented sausages in relation to their size. J. Sci. Food Agric. 2015, 96, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, J.A.; Hierro, E.M.; Bruna, J.M.; de la Hoz, L. Changes in the components of dry-fermented sausages during ripening. Crit. Rev. Food Sci. Nutr. 1999, 39, 329–367. [Google Scholar] [CrossRef]
- Flores, M. Understanding the implications of current health trends on the aroma of wet and dry cured meat products. Meat Sci. 2018, 144, 53–61. [Google Scholar] [CrossRef]
- Liu, S.Q. Practical implications of lactate and pyruvate metabolism by lactic acid bacteria in food and beverage fermentations. Int. J. Food Microbiol. 2003, 83, 115–131. [Google Scholar] [CrossRef]
- Montanari, C.; Barbieri, F.; Magnani, M.; Grazia, L.; Gardini, F.; Tabanelli, G. Phenotypic diversity of Lactobacillus sakei strains. Front. Microbiol. 2018, 9, 2003. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Sánchez-Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats—A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marco, A.; Navarro, J.L.; Flores, M. Volatile compounds of dry-fermented sausages as affected by solid-phase microextraction (SPME). Food Chem. 2004, 84, 633–641. [Google Scholar] [CrossRef]
- Flores, M.; Olivares, A. Flavor. In Handbook of Fermented Meat and Poultry; Toldrá, F., Hui, Y.H., Astiasarán, I., Sebranek, J.G., Talon, R., Eds.; Wiley Blackwell: Ames, IA, USA, 2014; pp. 217–225. [Google Scholar]
- Montanari, C.; Barbieri, F.; Gardini, F.; Tabanelli, G. Competition between starter cultures and wild microbial population in sausage fermentation: A case study regarding a typical Italian salami (Ventricina). Foods 2021, 10, 2138. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 Days | 4 Days | 30 Days | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Starter | p-Value | Starter | p-Value | Starter | p-Value | ||||||||||||||
| Casing | Microbial Group | SM-181 | TMX-M | SBM-11 | C | S | CxS | SM-181 | TMX-M | SBM-11 | C | S | C × S | SM-181 | TMX-M | SBM-11 | C | S | C × S |
| Natural | Lactic acid bacteria | 6.88 ± 0.03 | 7.01 ± 0.03 | 7.00 ± 0.03 | NS † | NS | NS | 8.21 a ± 0.02 | 8.84 b ± 0.02 | 8.62 c ± 0.02 | ** | ** | NS | 8.44 a ± 0.02 | 8.43 a ± 0.03 | 8.71 b ± 0.02 | ** | NS | * |
| Collagen | 6.97 ± 0.08 | 7.08 ± 0.05 | 6.94 ± 0.06 | 8.02 d ± 0.03 | 8.45 e ± 0.02 | 8.28 a ± 0.03 | 7.52 c ± 0.02 | 7.64 c ± 0.02 | 7.50 c ± 0.03 | ||||||||||
| Natural | Coagulase negative cocci | 6.63 a ± 0.03 | 6.08 b ± 0.02 | 6.77 a ± 0.02 | NS | ** | NS | 6.30 a ± 0.02 | 5.87 b ± 0.03 | 6.32 ac ± 0.02 | ** | ** | ** | 7.40 a ± 0.01 | 7.39 a± 0.02 | 7.51 a± 0.02 | NS | ** | ** |
| Collagen | 6.68 a ± 0.03 | 6.10 b ± 0.05 | 6.70 a ± 0.02 | 6.95 d ±0.01 | 7.48 e ± 0.01 | 6.48 c ± 0.02 | 7.81 b ±0.01 | 7.42 a ±0.03 | 7.24 c ±0.03 | ||||||||||
| Natural | Yeasts | 1.54 ± 0.04 | 1.51 ± 0.01 | 1.68 ± 0.05 | NS | NS | NS | 2.70 a ± 0.02 | 2.00 b ± 0.01 | 1.23 c ± 0.03 | ** | * | * | 1.96 a ± 0.04 | 2.00 a ± 0.01 | 1.35 b ± 0.04 | ** | ** | ** |
| Collagen | 1.52 ± 0.03 | 1.51 ± 0.04 | 1.67 ± 0.04 | 2.78 ad ± 0.03 | 2.96 d ± 0.04 | 2.00 b ± 0.01 | 6.15 c ± 0.04 | 6.79 d ± 0.01 | 6.50 e ± 0.01 | ||||||||||
| Natural | Enterococci | 2.32 ± 0.03 | 2.20 ± 0.02 | 2.49 ± 0.03 | NS | NS | NS | 2.05 a ± 0.05 | 2.05 a ± 0.06 | 2.01 a ± 0.09 | ** | NS | NS | 2.07 ad ± 0.03 | 2.09 ad ± 0.03 | 2.38 b± 0.05 | ** | ** | NS |
| Collagen | 2.27 ± 0.08 | 2.26 ± 0.03 | 2.45 ± 0.05 | 2.37 b± 0.05 | 2.65 b± 0.06 | 1.49 c± 0.05 | 1.89 ac ± 0.04 | 1.81 c ± 0.04 | 2.26 bd ± 0.05 | ||||||||||
| Natural | Enterobacteria | 1.95 ± 0.06 | 2.00 ± 0.04 | 1.81 ± 0.02 | NS | NS | NS | 1.00 a ± 0.07 | 1.29 ab ± 0.04 | 1.02 a ± 0.05 | ** | ** | * | 2.16 a ± 0.05 | 2.15 a ± 0.03 | 1.79 b ± 0.07 | ** | * | ** |
| Collagen | 1.95 ± 0.06 | 2.00 ± 0.03 | 1.81 ± 0.03 | 1.60 ab ± 0.06 | 1.77 b ± 0.06 | 1.00 a ± 0.05 | 1.30 c ± 0.07 | 1.86 b ± 0.09 | 1.48 c ± 0.05 | ||||||||||
| Natural Casing | Collagen Casing | |||||
|---|---|---|---|---|---|---|
| Compounds | SM-181 | TMX-M | SBM-11 | SM-181 | TMX-M | SBM-11 |
| Butanal | 0.01 ± 0.00 | 0.03 ± 0.01 | 0.03 ± 0.00 | 0.05 ± 0.00 | 0.04 ± 0.00 | 0.03 ± 0.00 |
| Hexanal | 0.25 ± 0.09 | 0.41 ± 0.06 | 0.50 ± 0.02 | 0.76 ± 0.05 | 0.49 ± 0.14 | 0.80 ± 0.07 |
| Nonanal | 2.09 a ± 0.18 | 2.49 a ± 0.05 | 6.16 b ± 0.76 | 2.49 a ± 0.23 | 1.79 a ± 0.17 | 1.76 a ± 0.02 |
| Decanal | 0.61 a ± 0.26 | 0.78 a ± 0.16 | 0.84 a ± 0.10 | 1.19 b ± 0.21 | 1.30 ab ± 0.05 | 1.26 ab ± 0.05 |
| Benzaldehyde | 0.26 ± 0.10 | 0.31 ± 0.02 | 0.39 ± 0.04 | 0.45 ± 0.08 | 0.48 ± 0.02 | 0.36 ± 0.02 |
| Benzeneacetaldehyde | 0.60 ± 0.14 | 0.43 ± 0.01 | 0.58 ± 0.07 | 0.44 ± 0.06 | 0.37 ± 0.03 | 0.44 ± 0.01 |
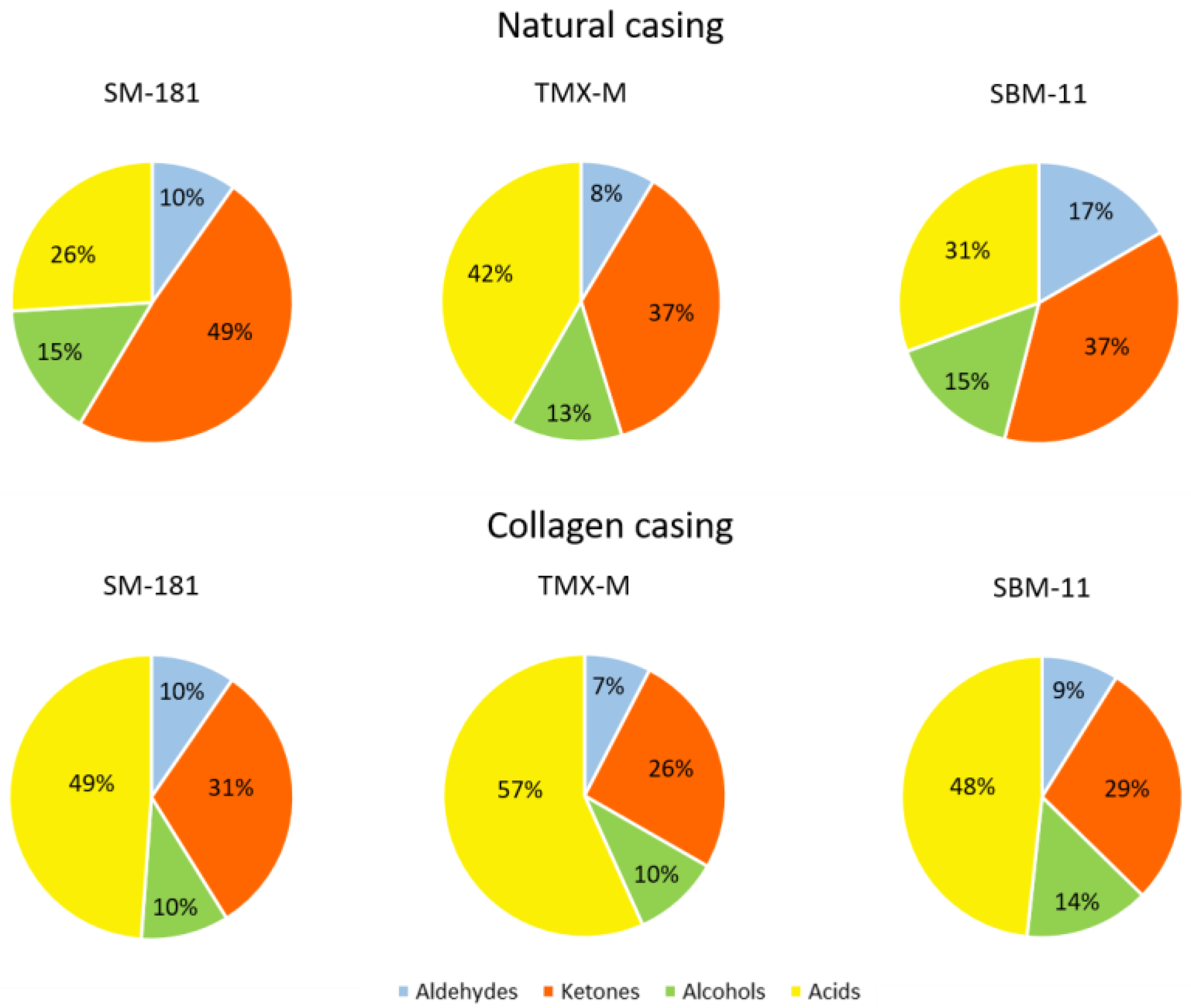
| ALDEHYDES | 3.82 a ± 0.10 | 4.45 b ± 0.16 | 8.50 b ± 0.93 | 5.38 ac ± 0.37 | 4.48 a ± 0.38 | 4.65 a ± 0.04 |
| Acetone | 4.04 a ± 0.11 | 3.39 b ± 0.12 | 3.29 b ± 0.17 | 6.21 c ± 0.43 | 4.02 a ± 0.06 | 4.70 d ± 0.31 |
| 2-butanone | 2.64 a ± 0.16 | 1.41 b ± 0.07 | 1.73 b ± 0.07 | 0.68 c ± 0.04 | 0.46 c ± 0.01 | 0.78 c ± 0.12 |
| 2-pentanone | 0.37 a ± 0.02 | 0.27 a ± 0.01 | 0.32 a ± 0.04 | 0.53 ab ± 0.05 | 0.45 a ± 0.03 | 0.60 c ± 0.06 |
| 2,3-butanedione | 0.89 a ± 0.13 | 1.08 a ± 0.17 | 0.87 a ± 0.16 | 0.47 b ± 0.03 | 0.29 b ± 0.08 | 0.15 b ± 0.02 |
| Methyl isobutil ketone | 1.30 a ± 0.02 | 1.08 b ± 0.03 | 1.14 b ± 0.00 | 1.31 a ± 0.06 | 1.42 a ± 0.11 | 1.37 a ± 0.03 |
| 5-methyl-3-hexanone | 0.30 ± 0.02 | 0.20 ± 0.03 | 0.11 ± 0.02 | 0.38 ± 0.01 | 0.12 ± 0.04 | 0.25 ± 0.03 |
| 4-methyl-3-penten-2-one | 4.07 a ± 0.98 | 5.43 b ± 0.11 | 5.68 b ± 0.11 | 5.53 b ± 0.12 | 6.07 bc ± 0.05 | 5.70 c ± 0.11 |
| 2-heptanone | 1.30 a ± 0.13 | 1.27 a ± 0.05 | 1.32 a ± 0.08 | 0 † bc | 0.15 c ± 0.09 | 0.44 c ± 0.03 |
| 2-octanone | 0.75 a ± 0.11 | 0.90 a ± 0.21 | 0.87 a ± 0.03 | 0.59 ab ± 0.02 | 0.34 b ± 0.10 | 0.18 b ± 0.10 |
| 3-hydroxy-2-butanone | 2.84 a ± 0.36 | 3.40 a ± 0.59 | 2.86 a ± 0.12 | 1.55 b ± 0.03 | 1.54 b ± 0.50 | 0.47 b ± 0.03 |
| 2-nonanone | 0.43 a ± 0.07 | 0.33 a ± 0.01 | 0.67 a ± 0.07 | 0.17 b ± 0.02 | 0.14 b ± 0.01 | 0.14 b ± 0.01 |
| Acetophenone | 0.17 ± 0.03 | 0.13 ± 0.01 | 0.16 ± 0.01 | 0.25 ± 0.02 | 0.14 ± 0.02 | 0.14 ± 0.01 |
| KETONES | 19.10 a ± 1.13 | 18.88 a ± 1.05 | 19.02 a ± 0.33 | 17.66 ab ± 0.63 | 15.14 b ± 0.37 | 14.92 b ± 0.38 |
| Isopropyl alcohol | 0.45 a ± 0.02 | 0.41 a ± 0.02 | 0.39 a ± 0.02 | 0.31 b ± 0.03 | 0.29 b ± 0.00 | 0.30 b ± 0.02 |
| Ethyl alcohol | 2.50 a ± 0.15 | 3.02 a ± 0.12 | 4.08 b ± 0.20 | 2.47 a ± 0.25 | 3.14 c ± 0.08 | 4.63 b ± 0.62 |
| 2-butanol | 0.41 a ± 0.02 | 0.47 a ± 0.01 | 0.36 a ± 0.03 | 0 b | 0 b | 0 b |
| 1-pentanol | 0.16 ± 0.01 | 0.22 ± 0.02 | 0.25 ± 0.03 | 0.20 ± 0.02 | 0.18 ± 0.01 | 0.21 ± 0.01 |
| 3-methyl-3-buten-1-ol | 0.17 ± 0.01 | 0.12 ± 0.01 | 0.08 ± 0.01 | 0.35 ± 0.02 | 0.09 ± 0.03 | 0.16 ± 0.01 |
| 3-methyl-2-buten-1-ol | 0.16 a ± 0.02 | 0.35 b ± 0.02 | 0.30 b ± 0.06 | 0.35 b ± 0.02 | 0.52 bc ± 0.03 | 0.57 c ± 0.05 |
| 1-hexanol | 0 a | 0.50 b ± 0.03 | 0.52 b ± 0.04 | 0.48 b ± 0.07 | 0.27 b ± 0.08 | 0.37 b ± 0.04 |
| 1-octen-3-ol | 1.54 a ± 0.20 | 0.88 b ± 0.05 | 0.99 ab ± 0.03 | 0.60 b ± 0.04 | 0.41 c ± 0.01 | 0.42 c ± 0.02 |
| 1-octanol | 0.28 a ± 0.01 | 0.36 a ± 0.03 | 0.71 a ± 0.07 | 0.26 b ± 0.01 | 0.24 a ± 0.02 | 0.20 a ± 0.01 |
| Benzyl alcohol | 0.14 a ± 0.00 | 0.27 b ± 0.02 | 0.20 ab ± 0.03 | 0.33 b ± 0.03 | 0.42 c ± 0.02 | 0.36 b ± 0.00 |
| Phenylethyl alcohol | 0.24 a ± 0.02 | 0.10 a ± 0.05 | 0 b | 0.25 a ± 0.02 | 0.34 a ± 0.03 | 0.33 a ± 0.04 |
| ALCOHOLS | 6.05 ± 0.09 | 6.68 ± 0.23 | 7.88 ± 0.46 | 5.59 ± 0.48 | 5.89 ± 0.21 | 7.54 ± 0.78 |
| Acetic acid | 6.43 a ± 0.20 | 14.31 b ± 1.23 | 8.14 a ± 1.05 | 15.37 a ± 1.57 | 19.42 bc ± 2.25 | 12.61 b ± 0.65 |
| Butanoic acid | 1.07 a ± 0.02 | 2.29 b ± 0.31 | 1.13 a ± 0.16 | 3.88 c ± 0.35 | 4.54 c ± 0.32 | 3.93 c ± 0.28 |
| 3-methyl-Butanoic acid | 0 a | 0 a | 0 a | 0.33 b ± 0.03 | 1.13 c ± 0.07 | 1.35 c ± 0.07 |
| Pentanoic acid | 0 a | 0 a | 0 a | 0 a | 0.20 b ± 0.06 | 0.26 b ± 0.01 |
| Hexanoic acid | 0.69 a ± 0.05 | 0.91 a ± 0.05 | 0.76 a ± 0.08 | 1.30 b ± 0.14 | 1.32 b ± 0.11 | 1.13 ab ± 0.09 |
| Heptanoic acid | 0.25 ± 0.01 | 0.26 ± 0.02 | 0.23 ± 0.02 | 0.34 ± 0.05 | 0.31 ± 0.02 | 0.30 ± 0.03 |
| Octanoic acid | 0.68 a ± 0.02 | 0.91 a ± 0.06 | 0.74 a ± 0.08 | 1.58 b ± 0.18 | 1.62 b ± 0.12 | 1.34 b ± 0.02 |
| Nonanoic acid | 0.57 a ± 0.02 | 0.85 a ± 0.05 | 0.82 a ± 0.10 | 1.12 b ± 0.12 | 0.88 a ± 0.05 | 1.21 b ± 0.12 |
| n-decanoic acid | 0.48 a ± 0.02 | 0.76 a ± 0.11 | 0.77 a ± 0.09 | 0.97 ab ± 0.11 | 1.27 b ± 0.06 | 1.22 b ± 0.08 |
| n-hexadecanoic acid | 0 a | 0 a | 0.90 b ± 0.52 | 0 a | 1.03 b ± 0.30 | 0.61 b ± 0.35 |
| Dodecanoic acid | 0 a | 0.62 b ± 0.18 | 1.00 b ± 0.41 | 1.36 bc ± 0.24 | 0.49 b ± 0.29 | 0.41 b ± 0.12 |
| Tetradecanoic acid | 0 a | 0.59 b ± 0.21 | 1.11 c ± 0.22 | 1.11 c ± 0.02 | 1.22 c ± 0.10 | 0.93 ac ± 0.11 |
| ACIDS | 10.16 a ± 0.21 | 21.50 b ± 2.01 | 15.61 ab ± 1.90 | 27.35 b ± 2.62 | 33.43 bc ± 2.85 | 25.31 b ± 1.32 |
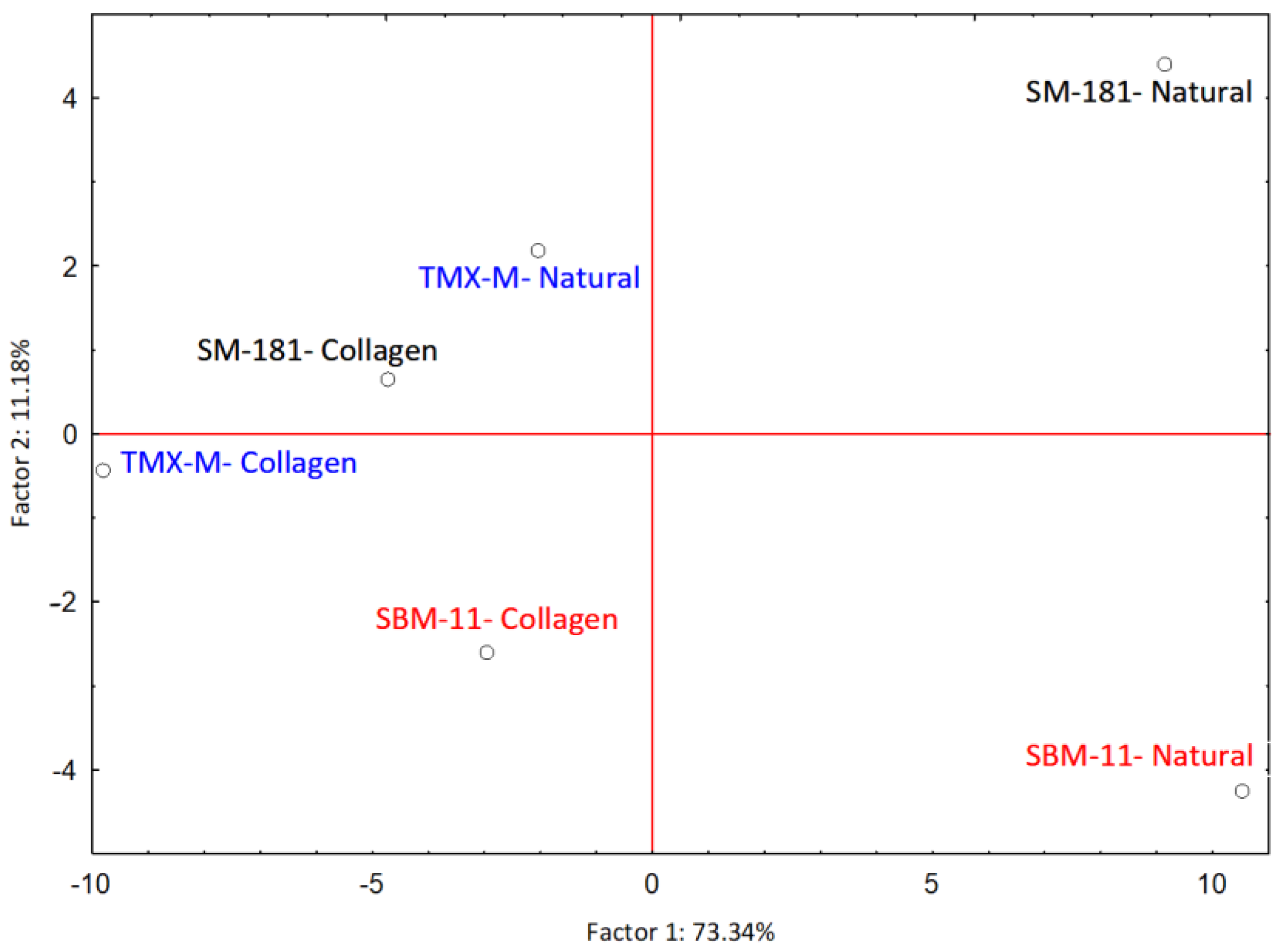
| Volatile Compound | Variable Contribution, Based on Covariances, to PC1 and PC2 | Factor Coordinates of the Variables PC1 and PC2, Based on Correlations | ||
|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | |
| Butanal | 0.0000 | 0.0000 | −0.0133 | −0.0041 |
| Hexanal | 0.0002 | 0.0035 | −0.1384 | −0.1888 |
| Nonanal | 0.0994 | 0.2143 | 2.5428 | −1.4578 |
| Decanal | 0.0011 | 0.0032 | −0.2767 | −0.1782 |
| Benzaldehyde | 0.0000 | 0.0001 | −0.0320 | −0.0362 |
| Benzeneacetaldehyde | 0.0011 | 0.0014 | 0.2727 | 0.1212 |
| Acetone | 0.0000 | 0.0812 | 0.0271 | 0.8975 |
| 2-butanone | 0.0524 | 0.1380 | 1.8463 | 1.1698 |
| 2-pentanone | 0.0000 | 0.0000 | −0.0374 | 0.0023 |
| 2,3-butanedione | 0.0062 | 0.0161 | 0.6368 | 0.4006 |
| Methyl isobutyl ketone | 0.0004 | 0.0044 | 0.1651 | 0.2108 |
| 5-methyl-3-hexanone | 0.0000 | 0.0022 | 0.0469 | 0.1503 |
| 4-methyl-3-penten-2-one | 0.0002 | 0.0159 | 0.1339 | −0.3979 |
| 2-heptanone | 0.0214 | 0.0182 | 1.1797 | 0.4253 |
| 2-octanone | 0.0037 | 0.0082 | 0.4960 | 0.2864 |
| 3-hydroxy-2-butanone | 0.0492 | 0.1567 | 1.7891 | 1.2468 |
| 2-nonanone | 0.0030 | 0.0000 | 0.4482 | −0.0144 |
| Acetophenone | 0.0000 | 0.0002 | 0.0328 | 0.0458 |
| Isopropyl alcohol | 0.0005 | 0.0020 | 0.1894 | 0.1419 |
| Ethyl alcohol | 0.0094 | 0.0859 | 0.7830 | −0.9231 |
| 2-butanol | 0.0021 | 0.0056 | 0.3741 | 0.2356 |
| 1-pentanol | 0.0000 | 0.0000 | 0.0527 | −0.0162 |
| 3-methyl-3-buten-1-ol | 0.0000 | 0.0006 | −0.0152 | 0.0821 |
| 3-methyl-2-buten-1-ol | 0.0004 | 0.0012 | −0.1621 | −0.1109 |
| 1-hexanol | 0.0000 | 0.0053 | −0.0533 | −0.2297 |
| 1-octen-3-ol | 0.0137 | 0.0486 | 0.9453 | 0.6946 |
| 1-octanol | 0.0014 | 0.0016 | 0.3028 | −0.1268 |
| Benzyl alcohol | 0.0002 | 0.0001 | −0.1349 | −0.0370 |
| Phenylethyl alcohol | 0.0001 | 0.0008 | −0.1066 | 0.0928 |
| Acetic acid | 0.6365 | 0.0274 | −6.4331 | 0.5217 |
| Butanoic acid | 0.0786 | 0.0136 | −2.2613 | −0.3680 |
| 3-methyl-Butanoic acid | 0.0081 | 0.0206 | −0.7272 | −0.4525 |
| Pentanoic acid | 0.0002 | 0.0008 | −0.1270 | −0.0939 |
| Hexanoic acid | 0.0010 | 0.0000 | −0.2654 | 0.0298 |
| Heptanoic acid | 0.0000 | 0.0001 | −0.0009 | 0.0403 |
| Octanoic acid | 0.0039 | 0.0001 | −0.5039 | −0.0380 |
| Nonanoic acid | 0.0001 | 0.0012 | −0.1102 | −0.1134 |
| n-decanoic acid | 0.0014 | 0.0050 | −0.3052 | −0.2226 |
| n-hexadecanoic acid | 0.0000 | 0.0501 | −0.0703 | −0.7055 |
| Dodecanoic acid | 0.0001 | 0.0148 | −0.1090 | −0.3840 |
| Tetradecanoic acid | 0.0022 | 0.0489 | −0.3815 | −0.6966 |
| Starter | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Casing | Attribute | SM-181 | TMX-M | SBM-11 | C | S | C × S |
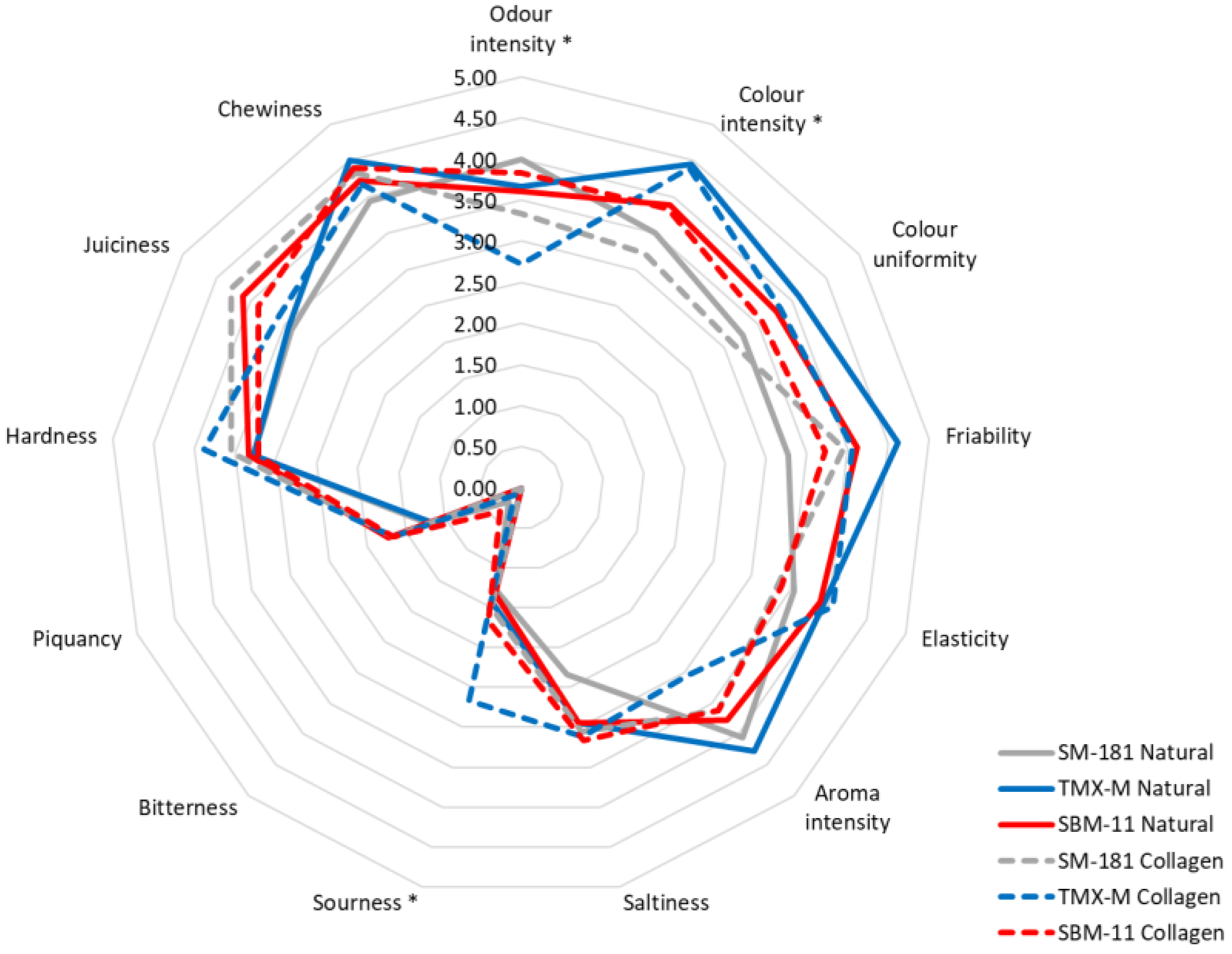
| Natural | Odour intensity | 4.00 a ± 0.08 | 3.67 ab ± 0.10 | 3.61 ab ± 0.11 | * | NS † | NS |
| Collagen | 3.22 bc ± 0.10 | 2.72 c ± 0.07 | 3.67 ab ± 0.08 | ||||
| Natural | Colour intensity | 3.50 a ± 0.10 | 4.44 b ± 0.10 | 3.89 ab ± 0.10 | NS | ** | NS |
| Collagen | 3.22 a ± 0.10 | 4.39 b ± 0.11 | 3.83 ab ± 0.08 | ||||
| Natural | Sourness | 1.28 a ± 0.13 | 1.44 a ± 0.06 | 1.33 a ± 0.11 | * | NS | NS |
| Collagen | 1.50 a ± 0.11 | 2.67 b ± 0.11 | 1.67 a ± 0.08 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montanari, C.; Barbieri, F.; Gardini, G.; Magnani, R.; Gottardi, D.; Gardini, F.; Tabanelli, G. Effects of Starter Cultures and Type of Casings on the Microbial Features and Volatile Profile of Fermented Sausages. Fermentation 2022, 8, 683. https://doi.org/10.3390/fermentation8120683
Montanari C, Barbieri F, Gardini G, Magnani R, Gottardi D, Gardini F, Tabanelli G. Effects of Starter Cultures and Type of Casings on the Microbial Features and Volatile Profile of Fermented Sausages. Fermentation. 2022; 8(12):683. https://doi.org/10.3390/fermentation8120683
Chicago/Turabian StyleMontanari, Chiara, Federica Barbieri, Gabriele Gardini, Rudy Magnani, Davide Gottardi, Fausto Gardini, and Giulia Tabanelli. 2022. "Effects of Starter Cultures and Type of Casings on the Microbial Features and Volatile Profile of Fermented Sausages" Fermentation 8, no. 12: 683. https://doi.org/10.3390/fermentation8120683