
Potential Prebiotic Effect of Cava Lees: Changes in Gut Microbiota

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Design
2.3. Intestinal Microbiota Extraction and Analysis
2.4. Statistic Analysis
3. Results and Discussion
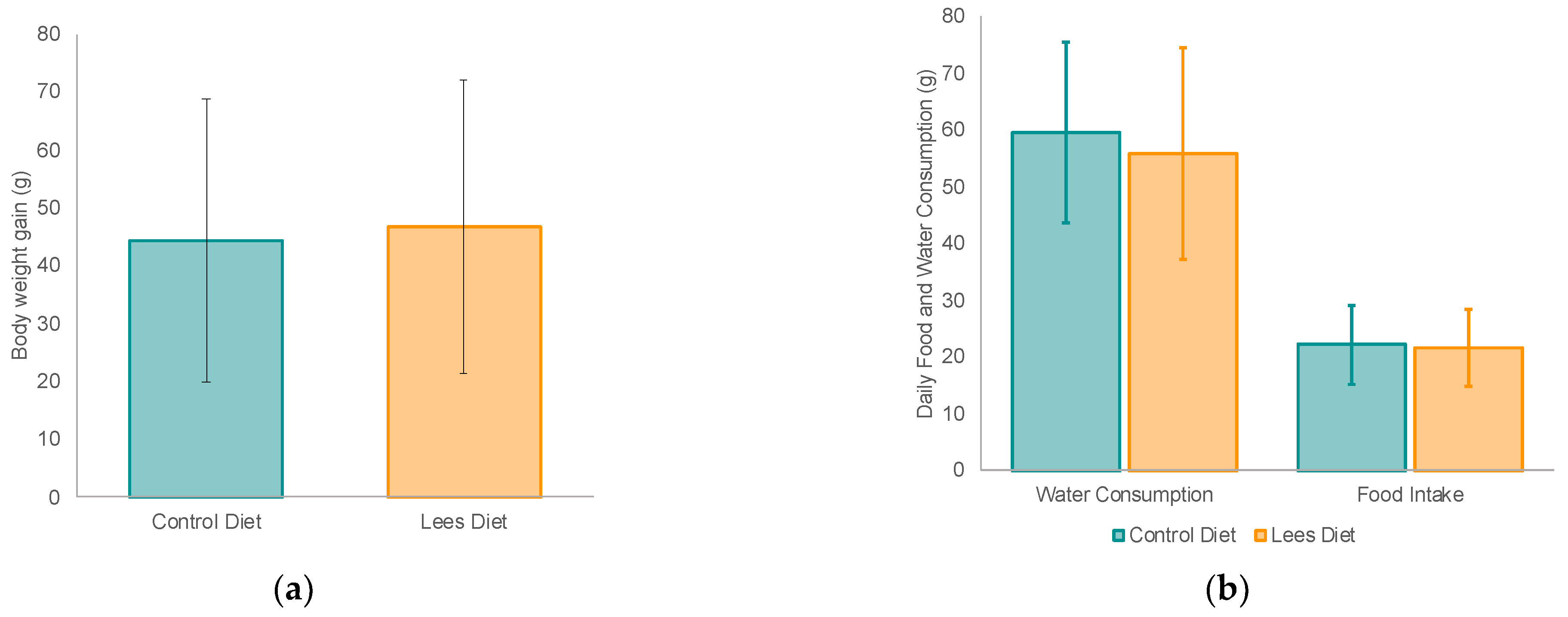
3.1. Effect of Cava Lees on Body Weght, Food Intake and Organs of Rats
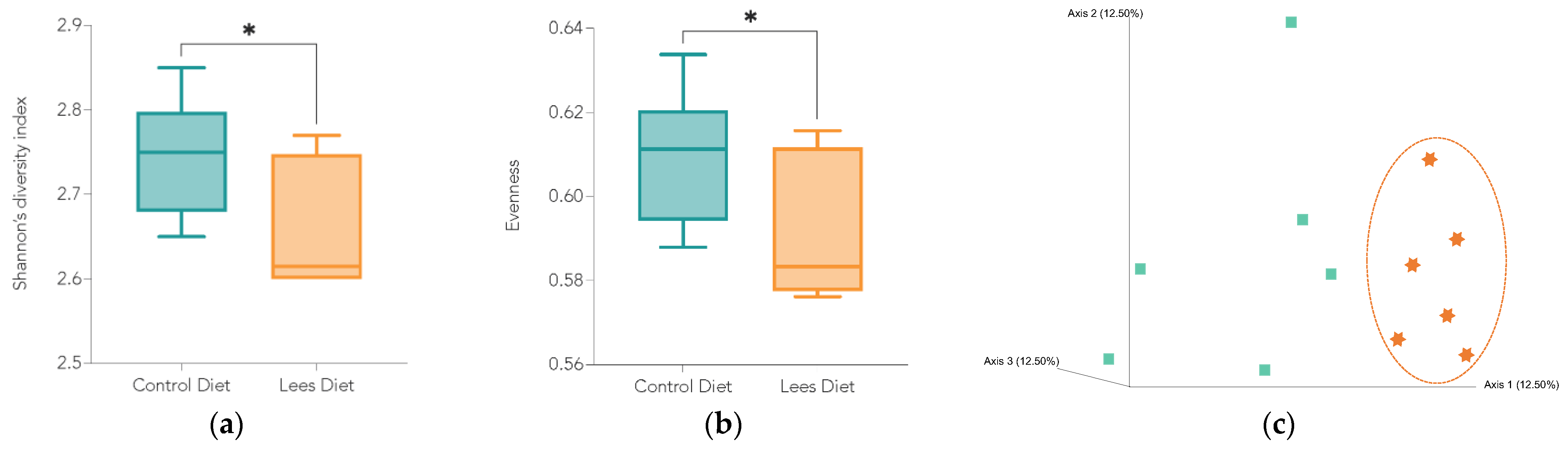
3.2. Effect of Cava Lees on Gut Microbiota Composition
Bacterial Diversity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Siró, I.; Kápolna, E.; Kápolna, B.; Lugasi, A. Functional Food. Product Development, Marketing and Consumer Acceptance—A Review. Appetite 2008, 51, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaluvadi, S.; Hotchkiss, A.T.; Yam, K.L. Chapter–36—Gut Microbiota: Impact of Probiotics, Prebiotics, Synbiotics, Pharmabiotics, and Postbiotics on Human Health. In Probiotics, Prebiotics, and Synbiotics; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 515–523. ISBN 978-0-12-802189-7. [Google Scholar]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, M.Á.; Casquete, R.; Córdoba, M.d.G.; Ruíz-Moyano, S.; Benito, M.J.; Pérez-Nevado, F.; Martín, A. Chemical Composition and Functional Properties of Dietary Fibre Concentrates from Winemaking By-Products: Skins, Stems and Lees. Foods 2021, 10, 1510. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; Nuñez-Gómez, V.; Navarro-González, I.; Sánchez-Martínez, L.; García-Alonso, J.; Periago, M.J.; González-Barrio, R. Raspberry Dietary Fibre: Chemical Properties, Functional Evaluation and Prebiotic in Vitro Effect. LWT 2020, 134, 110140. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA) Scientific Opinion on Dietary Reference Values for Carbohydrates and Dietary Fibre. EFSA J. 2010, 8, 1462. [CrossRef] [Green Version]
- Troilo, M.; Difonzo, G.; Paradiso, V.M.; Summo, C.; Caponio, F. Bioactive Compounds from Vine Shoots, Grape Stalks, and Wine Lees: Their Potential Use in Agro-Food Chains. Foods 2021, 10, 342. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health Benefits of Dietary Fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef]
- Ciudad-Mulero, M.; Fernández-Ruiz, V.; Matallana-González, M.C.; Morales, P. Chapter Two—Dietary Fiber Sources and Human Benefits: The Case Study of Cereal and Pseudocereals. In Advances in Food and Nutrition Research; Ferreira, I.C.F.R., Barros, L., Eds.; Functional Food Ingredients from Plants; Academic Press: Cambridge, MA, USA, 2019; Volume 90, pp. 83–134. [Google Scholar]
- Satija, A.; Hu, F.B. Cardiovascular Benefits of Dietary Fiber. Curr. Atheroscler Rep. 2012, 14, 505–514. [Google Scholar] [CrossRef]
- Stephen, A.M.; Champ, M.M.-J.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary Fibre in Europe: Current State of Knowledge on Definitions, Sources, Recommendations, Intakes and Relationships to Health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, B.; Peláez, C.; Martínez-Cuesta, M.C.; Parajó, J.C.; Alonso, J.L.; Requena, T. Emerging Prebiotics Obtained from Lemon and Sugar Beet Byproducts: Evaluation of Their in Vitro Fermentability by Probiotic Bacteria. LWT 2019, 109, 17–25. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Bajerska, J.; Zawirska-Wojtasiak, R.; Górecka, D. White Grape Pomace as a Source of Dietary Fibre and Polyphenols and Its Effect on Physical and Nutraceutical Characteristics of Wheat Biscuits. J. Sci. Food Agric. 2013, 93, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Mildner-Szkudlarz, S.; Zawirska-Wojtasiak, R.; Szwengiel, A.; Pacyński, M. Use of Grape By-Product as a Source of Dietary Fibre and Phenolic Compounds in Sourdough Mixed Rye Bread. Int. J. Food Sci. Technol. 2011, 46, 1485–1493. [Google Scholar] [CrossRef]
- Rivas, M.Á.; Benito, M.J.; Ruíz-Moyano, S.; Martín, A.; Córdoba, M.d.G.; Merchán, A.V.; Casquete, R. Improving the Viability and Metabolism of Intestinal Probiotic Bacteria Using Fibre Obtained from Vegetable By-Products. Foods 2021, 10, 2113. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine. 2021 Statistical Report on World Vitiviniculture; International Organisation of Vine and Wine: Paris, France, 2022. [Google Scholar]
- De Iseppi, A.; Lomolino, G.; Marangon, M.; Curioni, A. Current and Future Strategies for Wine Yeast Lees Valorization. Food Res. Int. 2020, 137, 109352. [Google Scholar] [CrossRef]
- Hernández-Macias, S.; Comas-Basté, O.; Jofré, A.; Bover-Cid, S.; Latorre-Moratalla, M.L.; Vidal-Carou, M.C. Growth-Promoting Effect of Cava Lees on Lactic Acid Bacteria Strains: A Potential Revalorization Strategy of a Winery By-Product. Foods 2021, 10, 1636. [Google Scholar] [CrossRef]
- Hernández-Macias, S.; Ferrer-Bustins, N.; Comas-Basté, O.; Jofré, A.; Latorre-Moratalla, M.; Bover-Cid, S.; Vidal-Carou, M. del C. Revalorization of Cava Lees to Improve the Safety of Fermented Sausages. Foods 2021, 10, 1916. [Google Scholar] [CrossRef]
- Alarcón, M.; López-Viñas, M.; Pérez-Coello, M.S.; Díaz-Maroto, M.C.; Alañón, M.E.; Soriano, A. Effect of Wine Lees as Alternative Antioxidants on Physicochemical and Sensorial Composition of Deer Burgers Stored during Chilled Storage. Antioxidants 2020, 9, 687. [Google Scholar] [CrossRef]
- Felix, M.; Martínez, I.; Sayago, A.; Recamales, M.Á.F. Wine Lees: From Waste to O/W Emulsion Stabilizer. Innov. Food Sci. Emerg. Technol. 2021, 74, 102810. [Google Scholar] [CrossRef]
- Martín-Garcia, A.; Riu-Aumatell, M.; López-Tamames, E. Revalorization of Cava (Spanish Sparkling Wine) Lees on Sourdough Fermentation. Fermentation 2022, 8, 133. [Google Scholar] [CrossRef]
- Martín-Garcia, A.; Riu-Aumatell, M.; López-Tamames, E. By-Product Revalorization: Cava Lees Can Improve the Fermentation Process and Change the Volatile Profile of Bread. Foods 2022, 11, 1361. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The Metagenomics RAST Server—A Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Li, S.; Qiao, E.; Tang, Z.; Jin, E.; Jin, G.; Gu, Y. Effects of Boron on Structure and Antioxidative Activities of Spleen in Rats. Biol. Trace Elem. Res. 2014, 158, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Altunkaynak, B.Z.; Ozbek, E.; Altunkaynak, M.E. A Stereological and Histological Analysis of Spleen on Obese Female Rats, Fed with High Fat Diet. Saudi Med. J. 2007, 28, 353–357. [Google Scholar]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of Targeted Delivery of Propionate to the Human Colon on Appetite Regulation, Body Weight Maintenance and Adiposity in Overweight Adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate Mediates a Microbiome–Brain–β-Cell Axis to Promote Metabolic Syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial Degradation of Complex Carbohydrates in the Gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, J.; Henning, S.M.; Lee, R.; Hsu, M.; Grojean, E.; Pisegna, R.; Ly, A.; Heber, D.; Li, Z. Dietary Pomegranate Extract and Inulin Affect Gut Microbiome Differentially in Mice Fed an Obesogenic Diet. Anaerobe 2017, 48, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Chen, J.; Miao, S.; Deng, K.; Liu, J.; Zeng, S.; Zheng, B.; Lu, X. Lotus Seed Oligosaccharides at Various Dosages with Prebiotic Activity Regulate Gut Microbiota and Relieve Constipation in Mice. Food Chem. Toxicol. 2019, 134, 110838. [Google Scholar] [CrossRef]
- Guo, X.; Cheng, M.; Zhang, X.; Cao, J.; Wu, Z.; Weng, P. Green Tea Polyphenols Reduce Obesity in High-Fat Diet-Induced Mice by Modulating Intestinal Microbiota Composition. Int. J. Food Sci. Technol. 2017, 52, 1723–1730. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia—A New Functional Genus with Potential Probiotic Properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Chase, A.B.; Weihe, C.; Orchanian, S.B.; Riedel, S.F.; Hendrickson, C.L.; Lay, M.; Sewall, J.M.; Martiny, J.B.H.; Whiteson, K. High-Fiber, Whole-Food Dietary Intervention Alters the Human Gut Microbiome but Not Fecal Short-Chain Fatty Acids. mSystems 2021, 6, e00115-21. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The Gut Microbiota of Laying Hens and Its Manipulation with Prebiotics and Probiotics To Enhance Gut Health and Food Safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]
- Han, X.; Ding, S.; Ma, Y.; Fang, J.; Jiang, H.; Li, Y.; Liu, G. Lactobacillus Plantarum and Lactobacillus Brevis Alleviate Intestinal Inflammation and Microbial Disorder Induced by ETEC in a Murine Model. Oxidative Med. Cell. Longev. 2021, 2021, e6867962. [Google Scholar] [CrossRef]
- Liévin-Le Moal, V.; Servin, A.L. Anti-Infective Activities of Lactobacillus Strains in the Human Intestinal Microbiota: From Probiotics to Gastrointestinal Anti-Infectious Biotherapeutic Agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary Fiber Intervention on Gut Microbiota Composition in Healthy Adults: A Systematic Review and Meta-Analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed]
- Babu, L.; Reddy, P.; Murali, H.S.; Batra, H.V. Optimization and Evaluation of a Multiplex PCR for Simultaneous Detection of Prominent Foodborne Pathogens of Enterobacteriaceae. Ann. Microbiol. 2013, 63, 1591–1599. [Google Scholar] [CrossRef]
- Mattos, G.N.; Tonon, R.V.; Furtado, A.A.L.; Cabral, L.M.C. Grape by-product extracts against microbial proliferation and lipid oxidation: A review. J. Sci. Food Agric. 2017, 97, 1055–1064. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Label | Lees Dose (mg/kg/Day) | Administered Volume (mL/kg/Day) | Animal Number | |
|---|---|---|---|---|---|
| Male | Female | ||||
| 1 | Control | 0 | 10 | 1–6 | 7–12 |
| 2 | Lees | 2000 1 | 10 | 13–18 | 19–24 |
| Organ | Control Diet | Lees Diet | p-Value |
|---|---|---|---|
| Thymus | 0.73 ± 0.14 | 0.68 ± 0.10 | 0.398 |
| Liver | 11.91 ± 3.71 | 11.21 ± 3.68 | 0.650 |
| Spleen | 0.82 ± 0.17 | 0.94 ± 0.24 | 0.158 |
| Kidneys | 2.11 ± 0.59 | 2.06 ± 0.45 | 0.802 |
| Family | Control Diet | Lees Diet | p-Value |
|---|---|---|---|
| Lachnospiraceae | 10.87 ± 1.91 | 15.22 ± 1.87 | 0.02 |
| Lactobacillaceae | 9.71 ± 0.83 | 11.71 ± 1.09 | 0.03 |
| Ruminococcaceae | 8.55 ± 1.53 | 11.00 ± 1.02 | 0.02 |
| Species | Control Diet | Lees Diet | p-Value |
|---|---|---|---|
| Lactobacillus brevis | 240 ± 124 | 671 ± 443 | 0.02 |
| Limosilactobacillus fermentum | 327 ± 226 | 1144 ± 905 | 0.02 |
| Lacticaseibacillus paracasei | 115 ± 68 | 370 ± 290 | 0.04 |
| Lactiplantibacillus plantarum | 636 ± 422 | 2072 ± 967 | 0.04 |
| Ligilactobacillus ruminis | 1109 ± 775 | 5224 ± 1628 | 0.02 |
| Latilactobacillus sakei | 166 ± 105 | 509 ± 286 | 0.04 |
| Ligilactobacillus salivarius | 1721 ± 1143 | 7684 ± 2820 | 0.02 |
| Family | Initial (t = 0 Days) | End (t = 14 Days) | p-Value |
|---|---|---|---|
| Enterobacteriaceae | 1249 ± 472 | 736 ± 352 | 0.07 |
| Clostridiaceae | 47,330 ± 14,339 | 37,557 ± 22,313 | 0.47 |
| Campylobacteraceae | 707 ± 555 | 366 ± 170 | 0.31 |
| Listeriaceae | 561 ± 144 | 360 ± 164 | 0.04 |
| Staphylococcaceae | 423 ± 134 | 260 ± 96 | 0.03 |
| Vibrionaceae | 354 ± 62 | 219 ± 118 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Garcia, A.; Gonzalez-Linares, J.; Riu-Aumatell, M.; López-Tamames, E. Potential Prebiotic Effect of Cava Lees: Changes in Gut Microbiota. Fermentation 2022, 8, 657. https://doi.org/10.3390/fermentation8110657
Martín-Garcia A, Gonzalez-Linares J, Riu-Aumatell M, López-Tamames E. Potential Prebiotic Effect of Cava Lees: Changes in Gut Microbiota. Fermentation. 2022; 8(11):657. https://doi.org/10.3390/fermentation8110657
Chicago/Turabian StyleMartín-Garcia, Alba, Javier Gonzalez-Linares, Montserrat Riu-Aumatell, and Elvira López-Tamames. 2022. "Potential Prebiotic Effect of Cava Lees: Changes in Gut Microbiota" Fermentation 8, no. 11: 657. https://doi.org/10.3390/fermentation8110657