
Simultaneous Production of Biohydrogen (bioH2) and Poly-Hydroxy-Alkanoates (PHAs) by a Photoheterotrophic Consortium Bioaugmented with Syntrophomonas wolfei
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
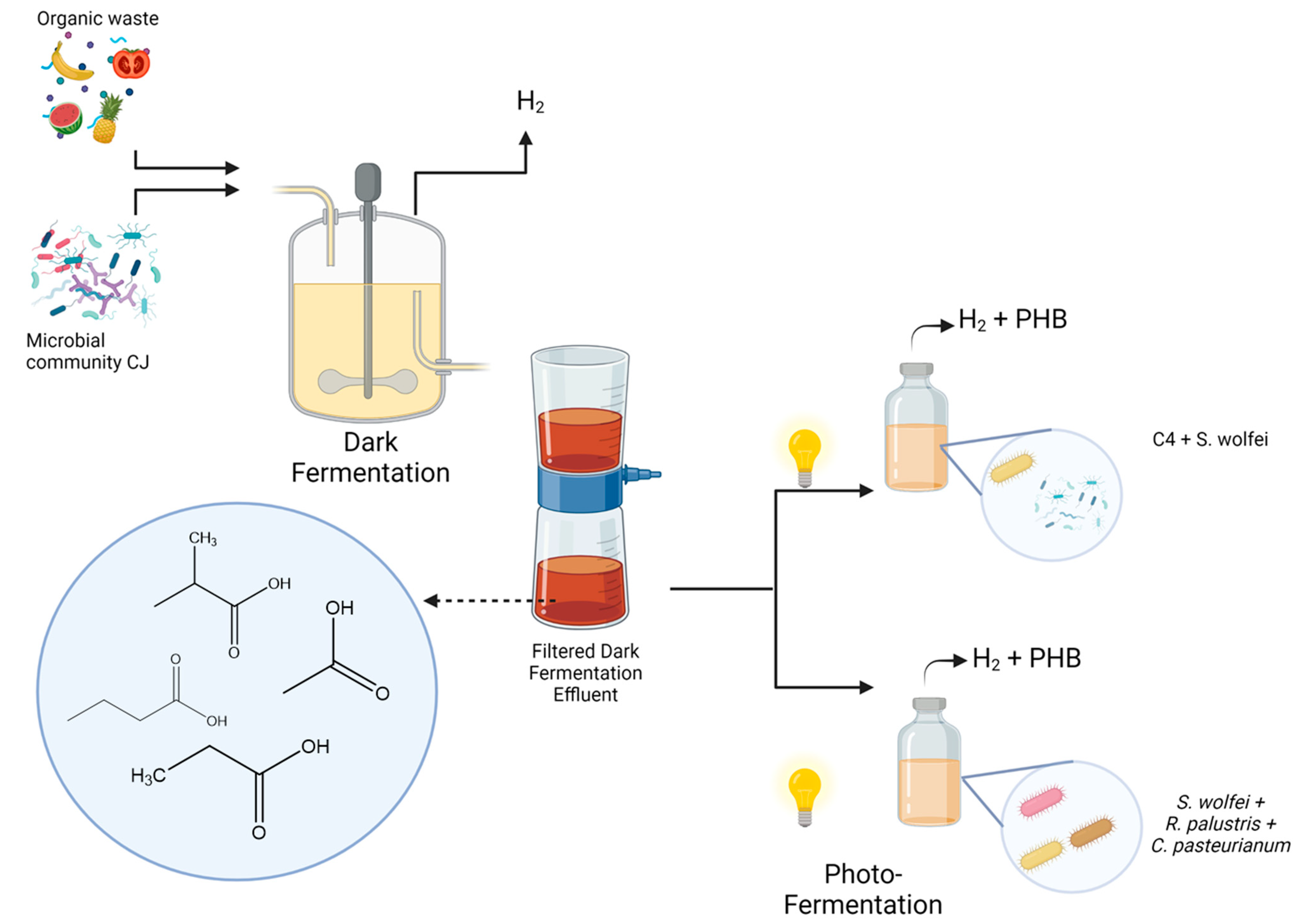
2.1. Inoculums for Dark- and Photofermentation Processes
2.1.1. Dark Fermentation with a Lactose-Adapted Consortium
2.1.2. R. palustris ATCC 17001 and the NPC-C4 Growth Stage
2.1.3. C. pasteurianum ATCC 6013
2.1.4. S. wolfei ATCC BAA 1933
2.2. Experimental Conditions
2.2.1. Dark Fermentation Process
2.2.2. Photofermentation Process
2.3. Chemical Analysis
2.3.1. Biomass (Protein) Quantification
2.3.2. bioH2 Gas Production
2.3.3. Nitrogen Quantification
2.3.4. OA Quantification
2.3.5. PHA Accumulation
Extraction
Biopolymer Characterization
3. Results
3.1. Individual Strain Cultures
3.1.1. C. Pasteurian
3.1.2. R. palustris
3.1.3. S. wolfei
3.2. Consortiums, Bioaugmented and Designed Cultures
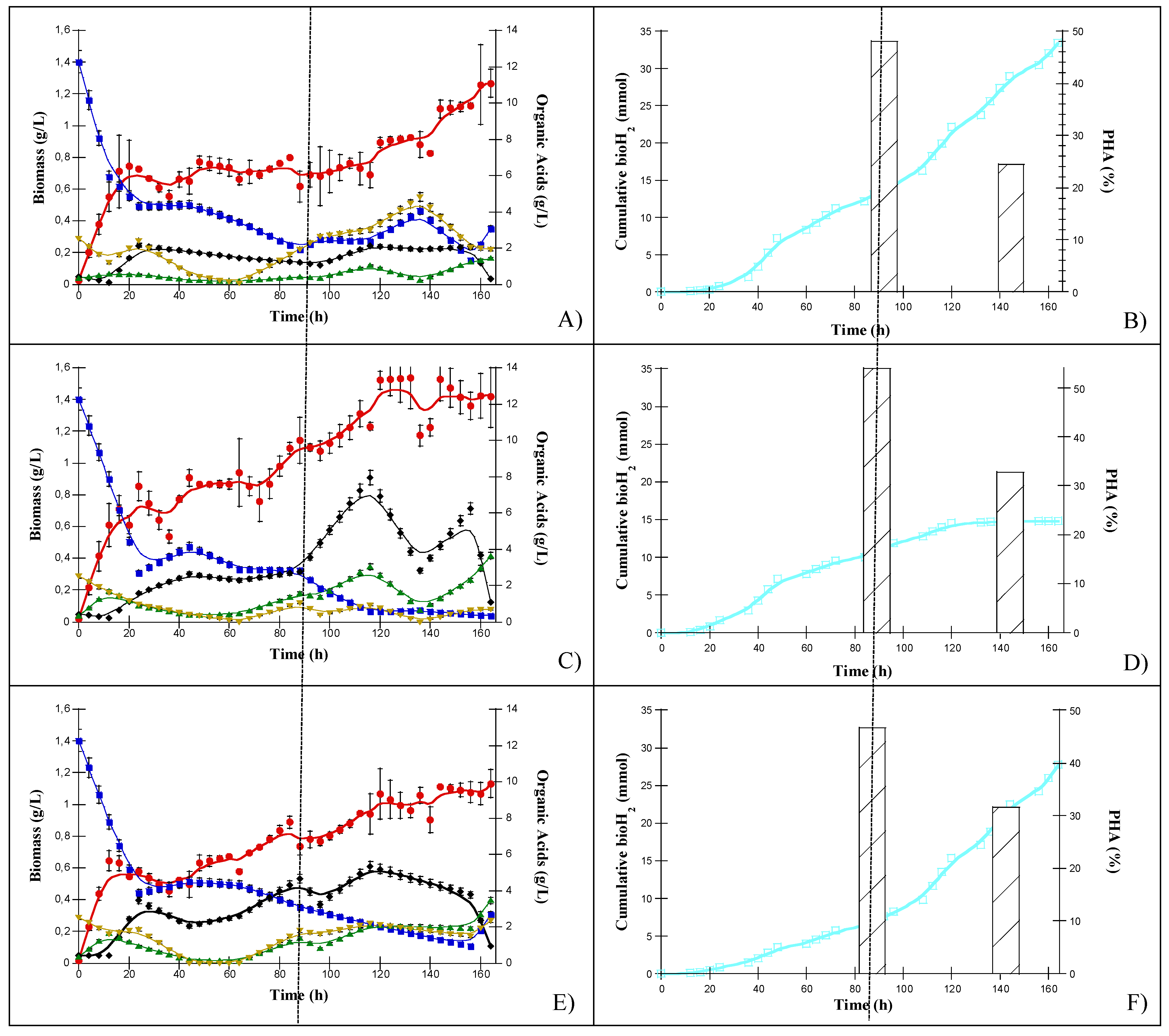
3.2.1. Natural Photoheterotrophic Consortium “NPC-C4”
3.2.2. Bioaugmented Natgural Photoheterotrophic Consortium with S. wolfei “BNPC-C4”
3.2.3. Designed Consortium
4. Discussion
5. Conclusions
Supplementary Materials
 ), H2VPR of R. palustris (
), H2VPR of R. palustris ( ) and H2VPR of S. wolfei (
) and H2VPR of S. wolfei ( ); (C) PHA C. pasteurianum (blue solid column), R. palustris (pink column), S. wolfei (crosse black column); and by the microbial consortiums: (B) H2VPR of “NPC-C4” (
); (C) PHA C. pasteurianum (blue solid column), R. palustris (pink column), S. wolfei (crosse black column); and by the microbial consortiums: (B) H2VPR of “NPC-C4” ( ), H2VPR of “BNPC-C4” (
), H2VPR of “BNPC-C4” ( ) and H2VPR of the “design consortium” (
) and H2VPR of the “design consortium” ( ); (D) PHA “NPC-C4” (solid golden column), “BNPC-C4” (brown column), “design consortium” (cross green column).
); (D) PHA “NPC-C4” (solid golden column), “BNPC-C4” (brown column), “design consortium” (cross green column). Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations and Acronyms
References
- Schwartz, E.; Fritsch, J.; Friedrich, B. H2-Metabolizing Prokaryotes. Prokaryotes: Prokaryotic Physiol. Biochem. 2013, 2, 119–199. [Google Scholar] [CrossRef]
- Niño-Navarro, C.; Chairez, I.; Christen, P.; Canul-Chan, M.; Garcia-Peña, E.I. Enhanced Hydrogen Production by a Sequential Dark and Photo Fermentation Process: Effects of Initial Feedstock Composition, Dilution and Microbial Population. Renew. Energy 2020, 147, 924–936. [Google Scholar] [CrossRef]
- Cheng, D.; Ngo, H.H.; Guo, W.; Chang, S.W.; Nguyen, D.D.; Bui, X.T.; Wei, W.; Ni, B.; Varjani, S.; Hoang, N.B. Enhanced Photo-Fermentative Biohydrogen Production from Biowastes: An Overview. Bioresour. Technol. 2022, 357, 127341. [Google Scholar] [CrossRef]
- Zong, W.; Yu, R.; Zhang, P.; Fan, M.; Zhou, Z. Efficient Hydrogen Gas Production from Cassava and Food Waste by a Two-Step Process of Dark Fermentation and Photo-Fermentation. Biomass Bioenergy 2009, 33, 1458–1463. [Google Scholar] [CrossRef]
- Liu, B.F.; Ren, N.Q.; Tang, J.; Ding, J.; Liu, W.Z.; Xu, J.F.; Cao, G.L.; Guo, W.Q.; Xie, G.J. Bio-Hydrogen Production by Mixed Culture of Photo- and Dark-Fermentation Bacteria. Int. J. Hydrogen Energy 2010, 35, 2858–2862. [Google Scholar] [CrossRef]
- Lo, Y.C.; Chen, C.Y.; Lee, C.M.; Chang, J.S. Photo Fermentative Hydrogen Production Using Dominant Components (Acetate, Lactate, and Butyrate) in Dark Fermentation Effluents. Int. J. Hydrogen Energy 2011, 36, 14059–14068. [Google Scholar] [CrossRef]
- Hitit, Z.Y.; Lazaro, C.Z.; Hallenbeck, P.C. Hydrogen Production by Co-Cultures of Clostridium butyricum and Rhodospeudomonas palustris: Optimization of Yield Using Response Surface Methodology. Int. J. Hydrogen Energy 2017, 42, 6578–6589. [Google Scholar] [CrossRef]
- McKinlay, J.B.; Oda, Y.; Ruhl, M.; Posto, A.L.; Sauer, U.; Harwood, C.S. Non-Growing Rhodopseudomonas palustris Increases the Hydrogen Gas Yield from Acetate by Shifting from the Glyoxylate Shunt to the Tricarboxylic Acid Cycle. J. Biol. Chem. 2014, 289, 1960–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, S.; Brigham, C. Inexpensive and Waste Raw Materials for PHAProduction. In The Handbook of Polyhydroxyalkanoates; Koller, M., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 203–220. [Google Scholar]
- McKinlay, J.B.; Harwood, C.S. Photobiological Production of Hydrogen Gas as a Biofuel. Curr. Opin. Biotechnol. 2010, 21, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Vincenzini, M.; Marchini, A.; Ena, A.; de Philippis, R. H2 and Poly-b-Hydroxybutyrate, Two Alternative Chemicals from Purple Non Sulfur Bacteria. Biotechnol. Lett. 1997, 19, 759–762. [Google Scholar] [CrossRef]
- Thulasidharan, D.; Arumugam, A.; Uppuluri, K.B. Research and Economic Perspectives on an Integrated Biorefinery Approach for the Simultaneous Production of Polyhydroxyalkanoates and Biohydrogen. Int. J. Biol. Macromol. 2021, 193, 1937–1951. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-Y.; Shi, Z.-Y.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. Enhanced Co-Production of Hydrogen and Poly-(R)-3-Hydroxybutyrate by Recombinant PHB Producing E. Coli over-Expressing Hydrogenase 3 and Acetyl-CoA Synthetase. Metab. Eng. 2012, 14, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Rafa, N.; Mofijur, M.; Badruddin, I.A.; Inayat, A.; Ali, M.S.; Farrok, O.; Yunus Khan, T.M. Biohydrogen Production From Biomass Sources: Metabolic Pathways and Economic Analysis. Front. Energy Res. 2021, 9, 753878. [Google Scholar] [CrossRef]
- Fernández-Dacosta, C.; Posada, J.A.; Kleerebezem, R.; Cuellar, M.C.; Ramirez, A. Microbial Community-Based Polyhydroxyalkanoates (PHAs) Production from Wastewater: Techno-Economic Analysis and Ex-Ante Environmental Assessment. Bioresour. Technol. 2015, 185, 368–377. [Google Scholar] [CrossRef]
- Manikandan, N.A.; Pakshirajan, K.; Pugazhenthi, G. Techno-Economic Assessment of a Sustainable and Cost-Effective Bioprocess for Large Scale Production of Polyhydroxybutyrate. Chemosphere 2021, 284, 131371. [Google Scholar] [CrossRef] [PubMed]
- Montiel-Corona, V.; Revah, S.; Morales, M. Hydrogen Production by an Enriched Photoheterotrophic Culture Using Dark Fermentation Effluent as Substrate: Effect of Flushing Method, Bicarbonate Addition, and Outdoor–Indoor Conditions. Int. J. Hydrogen Energy 2015, 40, 9096–9105. [Google Scholar] [CrossRef]
- Xu, C.; Yu, H. Insights into Constructing a Stable and Efficient Microbial Consortium. Chin. J. Chem. Eng. 2021, 30, 112–120. [Google Scholar] [CrossRef]
- Guerra-Blanco, P.; Cortes, O.; Poznyak, T.; Chairez, I.; García-Peña, E.I. Polyhydroxyalkanoates (PHA) Production by Photoheterotrophic Microbial Consortia: Effect of Culture Conditions over Microbial Population and Biopolymer Yield and Composition. Eur. Polym. J. 2018, 98, 94–104. [Google Scholar] [CrossRef]
- Lopez-Romero, J.; Salgado-Manjarrez, E.; Torres, L.; Garcia-Peña, E.I. Enhanced Carotenoid Production by Rhodopseudomonas palustris ATCC 17001 under Low Light Conditions. J. Biotechnol. 2020, 323, 159–165. [Google Scholar] [CrossRef]
- Cabrol, L.; Marone, A.; Tapia-Venegas, E.; Steyer, J.-P.; Ruiz-Filippi, G.; Trably, E. Microbial Ecology of Fermentative Hydrogen Producing Bioprocesses: Useful Insights for Driving the Ecosystem Function. FEMS Microbiol. Rev. 2017, 41, 158–181. [Google Scholar] [CrossRef]
- Ergal, İ.; Bochmann, G.; Fuchs, W.; Rittmann, S.K.-M. Design and Engineering of Artificial Microbial Consortia for Biohydrogen Production. Curr. Opin. Biotechnol. 2022, 73, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.E.; Oda, Y.; Harwood, C.S. Regulation of Uptake Hydrogenase and Effects of Hydrogen Utilization on Gene Expression in Rhodopseudomonas palustris. J. Bacteriol. 2006, 188, 6143–6151. [Google Scholar] [CrossRef] [Green Version]
- Hitit, Z.Y.; Zampol Lazaro, C.; Hallenbeck, P.C. Increased Hydrogen Yield and COD Removal from Starch/Glucose Based Medium by Sequential Dark and Photo-Fermentation Using Clostridium butyricum and Rhodopseudomonas palustris. Int. J. Hydrogen Energy 2017, 42, 18832–18843. [Google Scholar] [CrossRef]
- Kao, P.-M.; Hsu, B.-M.; Huang, K.-H.; Tao, C.-W.; Chang, C.-M.; Ji, W.-T. Biohydrogen Production by Immobilized Co-Culture of Clostridium butyricum and Rhodopseudomonas palustris. Energy Procedia 2014, 61, 834–837. [Google Scholar] [CrossRef] [Green Version]
- Jurado-Marban, V.H.; Tapia-Bustos, M.A.; Gonzalez-Garcia, R.A.; Salgado, E.; Garcia-Peña, E.I. Hydrogen Production by a Mixed Photoheterotrophic Culture: Correlation between Gene Expression Analysis and Physiological Behavior. Int. J. Hydrogen Energy 2019, 44, 641–651. [Google Scholar] [CrossRef]
- Gomez-Romero, J.; Gonzalez-Garcia, A.; Chairez, I.; Torres, L.; Garcia-Peña, E.I. Selective Adaptation of an Anaerobic Microbial Community: Biohydrogen Production by Co-Digestion of Cheese Whey and Vegetables Fruit Waste. Int. J. Hydrogen Energy 2014, 39, 12541–12550. [Google Scholar] [CrossRef]
- Cheng, Y.; Wei, H.; Sun, R.; Tian, Z.; Zheng, X. Rapid Method for Protein Quantitation by Bradford Assay after Elimination of the Interference of Polysorbate 80. Anal. Biochem. 2016, 494, 37–39. [Google Scholar] [CrossRef]
- Govindaraju, A.; McKinlay, J.B.; LaSarre, B. Phototrophic Lactate Utilization by Rhodopseudomonas palustris Is Stimulated by Coutilization with Additional Substrates. Appl. Environ. Microbiol. 2019, 85, e00048-19. [Google Scholar] [CrossRef] [Green Version]
- Diez-Gonzalez, F.; Russell, J.B.; Hunter, J.B. The Role of an NAD-Independent Lactate Dehydrogenase and Acetate in the Utilization of Lactate by Clostridium acetobutylicum Strain P262. Arch. Microbiol. 1995, 164, 36–42. [Google Scholar] [CrossRef]
- Buckel, W.; Thauer, R.K. Flavin-Based Electron Bifurcation, A New Mechanism of Biological Energy Coupling. Chem. Rev. 2018, 118, 3862–3886. [Google Scholar] [CrossRef]
- Demmer, J.K.; Pal Chowdhury, N.; Selmer, T.; Ermler, U.; Buckel, W. The Semiquinone Swing in the Bifurcating Electron Transferring Flavoprotein/Butyryl-CoA Dehydrogenase Complex from Clostridium difficile. Nat. Commun. 2017, 8, 1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Lu, W.-B.; Liu, C.-H.; Chang, J.-S. Improved Phototrophic H2 Production with Rhodopseudomonas palustris WP3-5 Using Acetate and Butyrate as Dual Carbon Substrates. Bioresour. Technol. 2008, 99, 3609–3616. [Google Scholar] [CrossRef] [PubMed]
- Cardeña, R.; Valdez-Vazquez, I.; Buitrón, G. Effect of Volatile Fatty Acids Mixtures on the Simultaneous Photofermentative Production of Hydrogen and Polyhydroxybutyrate. Bioprocess Biosyst. Eng. 2017, 40, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Anand, A.; Dubey, V.K.; Moholkar, V.S. Metabolic Flux Network Analysis of Hydrogen Production from Crude Glycerol by Clostridium pasteurianum. Bioresour. Technol. 2017, 242, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.J.; Rocha, J.M.S.; Tramper, J.; Wijffels, R.H. Acetate as a Carbon Source for Hydrogen Production by Photosynthetic Bacteria. J. Biotechnol. 2001, 85, 25–33. [Google Scholar] [CrossRef]
- Coma, M.; Vilchez-Vargas, R.; Roume, H.; Jauregui, R.; Pieper, D.H.; Rabaey, K. Product Diversity Linked to Substrate Usage in Chain Elongation by Mixed-Culture Fermentation. Environ. Sci. Technol. 2016, 50, 6467–6476. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Popp, D.; Sträuber, H.; Harms, H.; Kleinsteuber, S. Draft Genome Sequences of Three Clostridia Isolates Involved in Lactate-Based Chain Elongation. Microbiol. Resour. Announc. 2020, 9, e00679-20. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Hoshino, Y.; Nakase, K.; Usuda, Y. Reduction of Hydrogen Peroxide Stress Derived from Fatty Acid Beta-Oxidation Improves Fatty Acid Utilization in Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Pinske, C.; Jaroschinsky, M.; Linek, S.; Kelly, C.L.; Sargent, F.; Sawers, R.G. Physiology and Bioenergetics of [NiFe]-Hydrogenase 2-Catalyzed H2 -Consuming and H2 -Producing Reactions in Escherichia coli. J. Bacteriol. 2015, 197, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Schut, G.J.; Adams, M.W.W. The Iron-Hydrogenase of Thermotoga maritima Utilizes Ferredoxin and NADH Synergistically: A New Perspective on Anaerobic Hydrogen Production. J. Bacteriol. 2009, 191, 4451–4457. [Google Scholar] [CrossRef]
- Weghoff, M.C.; Bertsch, J.; Müller, V. A Novel Mode of Lactate Metabolism in Strictly Anaerobic Bacteria. Environ. Microbiol. 2015, 17, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Beaty, P.S.; McInerney, M.J. Effects of Organic Acid Anions on the Growth and Metabolism of Syntrophomonas wolfei in Pure Culture and in Defined Consortia. Appl. Environ. Microbiol. 1989, 55, 977–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detman, A.; Mielecki, D.; Pleśniak, Ł.; Bucha, M.; Janiga, M.; Matyasik, I.; Chojnacka, A.; Jędrysek, M.-O.; Błaszczyk, M.K.; Sikora, A. Methane-Yielding Microbial Communities Processing Lactate-Rich Substrates: A Piece of the Anaerobic Digestion Puzzle. Biotechnol. Biofuels 2018, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Detman, A.; Mielecki, D.; Chojnacka, A.; Salamon, A.; Błaszczyk, M.K.; Sikora, A. Cell Factories Converting Lactate and Acetate to Butyrate: Clostridium Butyricum and Microbial Communities from Dark Fermentation Bioreactors. Microb. Cell Fact 2019, 18, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, J.R.; Sims, D.R.; Han, C.; Kim, E.; Lykidis, A.; Lapidus, A.L.; McDonnald, E.; Rohlin, L.; Culley, D.E.; Gunsalus, R.; et al. The Genome of Syntrophomonas wolfei: New Insights into Syntrophic Metabolism and Biohydrogen Production. Environ. Microbiol. 2010, 12, 2289–2301. [Google Scholar] [CrossRef]
- Beaty, P.S.; McInerney, M.J. Growth of Syntrophomonas wolfei in Pure Culture on Crotonate. Arch. Microbiol. 1987, 147, 389–393. [Google Scholar] [CrossRef]
- McInerney, M.J.; Bryant, M.P.; Hespell, R.B.; Costerton, J.W. Syntrophomonas wolfei Gen. Nov. Sp. Nov., an Anaerobic, Syntrophic, Fatty Acid-Oxidizing Bacterium. Appl. Environ. Microbiol. 1981, 41, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.-C.; Chen, S.-D.; Chen, C.-Y.; Huang, T.-I.; Lin, C.-Y.; Chang, J.-S. Combining Enzymatic Hydrolysis and Dark–Photo Fermentation Processes for Hydrogen Production from Starch Feedstock: A Feasibility Study. Int. J. Hydrogen Energy 2008, 33, 5124–5233. [Google Scholar] [CrossRef]
- Su, H.; Cheng, J.; Zhou, J.; Song, W.; Cen, K. Improving Hydrogen Production from Cassava Starch by Combination of Dark and Photo Fermentation. Int. J. Hydrogen Energy 2009, 34, 1780–1786. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Yang, M.-H.; Yeh, K.-L.; Liu, C.-H.; Chang, J.-S. Biohydrogen Production Using Sequential Two-Stage Dark and Photo Fermentation Processes. Int. J. Hydrogen Energy 2008, 33, 4755–4762. [Google Scholar] [CrossRef]
- Kremp, F.; Roth, J.; Müller, V. The Sporomusa Type Nfn Is a Novel Type of Electron-Bifurcating Transhydrogenase That Links the Redox Pools in Acetogenic Bacteria. Sci. Rep. 2020, 10, 14872. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture | Total OAs Removed (%) | Total COD Removed (%) | Final Biomass Concentration (g/L) | Cumulative bioH2 (mmol) | H2VPR a (mL/Lh) | H2 Yield PF (mL/gOA) | Overall H2 Yield DF-PF b (mL/gCOD) | PHA Production (%) 88 and 134 h | |
|---|---|---|---|---|---|---|---|---|---|
| C. pasteurianum | 57 | 50 | 1.3 | 33 | 27 | 158.8 | 495.4 [15.4] c | 48.8 | 24.5 |
| R. palustris | 63 | 55 | 1.5 | 15 | 20 | 81.9 | 428.8 [13.4] c | 54.0 | 32.8 |
| S. wolfei | 39 | 28 | 1.1 | 28 | 20 | 94.4 | 553.9 [17.3] c | 46.6 | 31.6 |
| C4 | 57 | 52 | 1.3 | 26 | 28 | 128.2 | 467.9 [14.6] c | 32.2 | 48.3 |
| C4 + S.wolfei | 59 | 52 | 1.2 | 32 | 28 | 161.8 | 487.8 [15.2] c | 29.0 | 50.1 |
| Designed consortium d | 53 | 43 | 1.3 | 45 | 40 | 195.4 | 561.9 [17.5] c | 41.1 | 50.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, A.; Salgado, E.; Vanegas, Z.; Niño-Navarro, C.; Cortés, O.; Chairez, I.; García-Peña, E.I. Simultaneous Production of Biohydrogen (bioH2) and Poly-Hydroxy-Alkanoates (PHAs) by a Photoheterotrophic Consortium Bioaugmented with Syntrophomonas wolfei. Fermentation 2022, 8, 644. https://doi.org/10.3390/fermentation8110644
González A, Salgado E, Vanegas Z, Niño-Navarro C, Cortés O, Chairez I, García-Peña EI. Simultaneous Production of Biohydrogen (bioH2) and Poly-Hydroxy-Alkanoates (PHAs) by a Photoheterotrophic Consortium Bioaugmented with Syntrophomonas wolfei. Fermentation. 2022; 8(11):644. https://doi.org/10.3390/fermentation8110644
Chicago/Turabian StyleGonzález, Axayacatl, Edgar Salgado, Zaira Vanegas, Cristina Niño-Navarro, Omar Cortés, Isaac Chairez, and Elvia I. García-Peña. 2022. "Simultaneous Production of Biohydrogen (bioH2) and Poly-Hydroxy-Alkanoates (PHAs) by a Photoheterotrophic Consortium Bioaugmented with Syntrophomonas wolfei" Fermentation 8, no. 11: 644. https://doi.org/10.3390/fermentation8110644