

Schizosaccharomyces pombe in the Brewing Process: Mixed-Culture Fermentation for More Complete Attenuation of High-Gravity Wort


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Yeast Isolation and Storage
2.3. Wort Production (Standard-Gravity Wort)
2.4. Wort Production (High-Gravity Wort)
2.5. Propagation of Yeast Cultures
2.6. Inoculation of Wort
2.7. Monitoring and Modelling of Fermentation Kinetics
2.8. Determination of Free Amino Nitrogen (FAN) Content
2.9. Alcohol by Volume (ABV), pH, Specific Gravity (SG), Extract, and Real Degree of Fermentation (RDF)
2.10. Determination of Yeast Metabolite Concentration
2.11. Statistical Analysis
3. Results
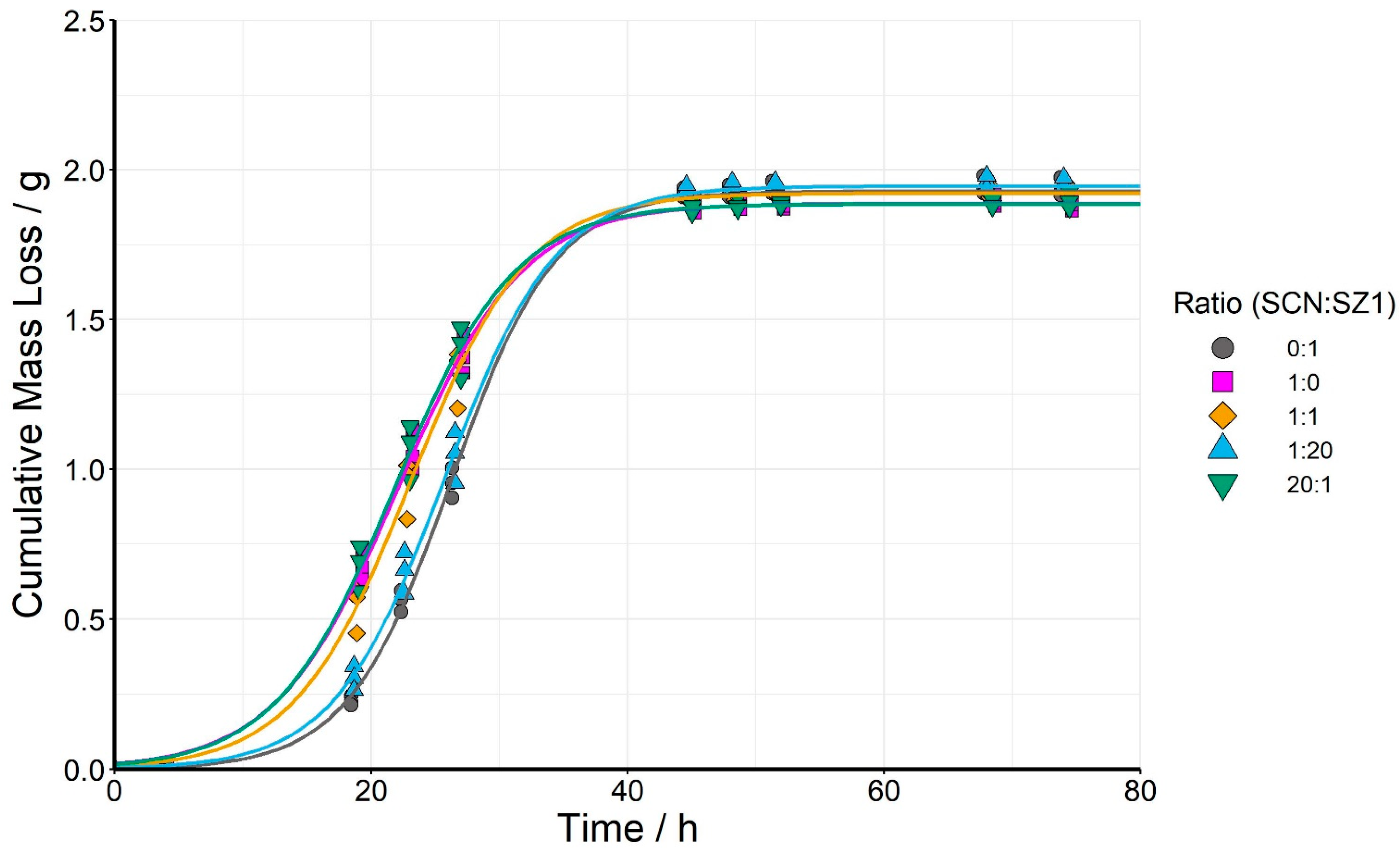
3.1. Fermentation of Standard-Gravity Wort by S. cerevisiae (SCN) and S. pombe (SZ1) Monocultures, and Three Mixed Cultures at 20:1, 1:1, and 1:20 Initial Inoculation Ratio
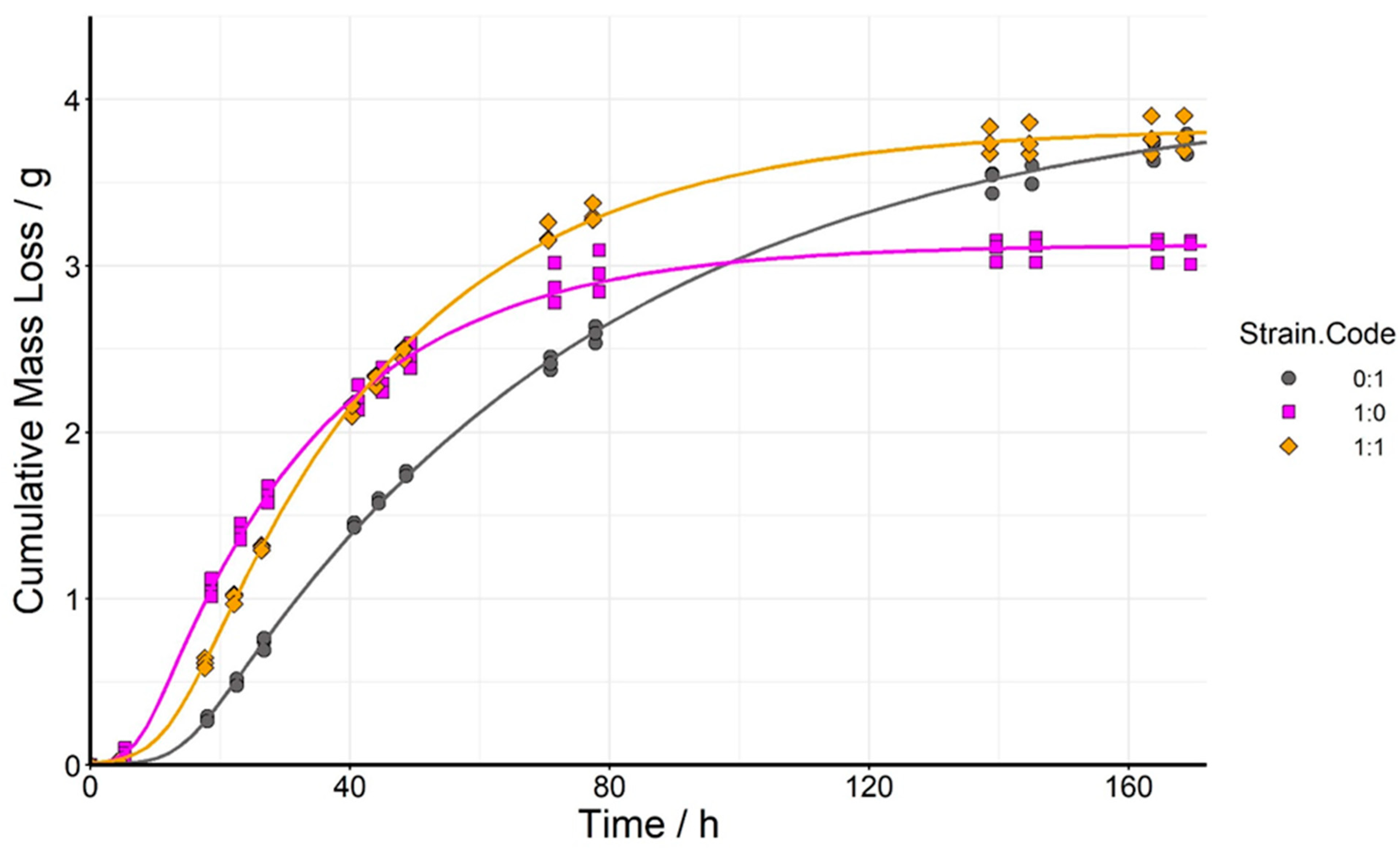
3.2. Fermentation of High-Gravity Wort by S. cerevisiae (SCN) and S. pombe (SZ1) Monocultures and by Mixed Culture at 1:1 Initial Inoculation Ratio
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stewart, G.G. Brewing Intensification Through the Lens of the Craft Brewer. MBAA Technol. Q. 2016, 53, 192–199. [Google Scholar] [CrossRef]
- Puligundla, P.; Smogrovicova, D.; Sarathi, V.; Obulam, R.; Ko, S. Very High Gravity (VHG) Ethanolic Brewing and Fermentation: A Research Update. J. Ind. Microbiol. Biotechnol. 2011, 38, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Hackstaff, B.W. Various Aspects of High Gravity Brewing. MBAA Technol. Q. 1978, 15, 1–7. [Google Scholar]
- Boulton, C.; Quain, D. Brewing Yeast and Fermentation; Blackwell Science Ltd.: Oxford, UK, 2001; ISBN 0632054751. [Google Scholar]
- Saerens, S.M.G.; Verbelen, P.J.; Vanbeneden, N.; Thevelein, J.M.; Delvaux, F.R. Monitoring the Influence of High-Gravity Brewing and Fermentation Temperature on Flavour Formation by Analysis of Gene Expression Levels in Brewing Yeast. Appl. Microbiol. Biotechnol. 2008, 80, 1039–1051. [Google Scholar] [CrossRef]
- Lei, H.; Zhao, H.; Yu, Z.; Zhao, M. Effects of Wort Gravity and Nitrogen Level on Fermentation Performance of Brewer’s Yeast and the Formation of Flavor Volatiles. Appl. Biochem. Biotechnol. 2012, 166, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Pratt, P.L.; Bryce, J.H.; Stewart, G.G. The Effects of Osmotic Pressure and Ethanol on Yeast Viability and Morphology. J. Inst. Brew. 2003, 109, 218–228. [Google Scholar] [CrossRef]
- Cooper, D.J.; Stewart, G.G.; Bryce, J.H. Some Reasons Why High Gravity Brewing Has a Negative Effect on Head Retention. J. Inst. Brew. 1998, 104, 83–87. [Google Scholar] [CrossRef]
- Liu, J.J.; Ding, W.T.; Zhang, G.C.; Wang, J.Y. Improving Ethanol Fermentation Performance of Saccharomyces cerevisiae in Very High-Gravity Fermentation through Chemical Mutagenesis and Meiotic Recombination. Appl. Microbiol. Biotechnol. 2011, 91, 1239–1246. [Google Scholar] [CrossRef]
- Ekberg, J.; Rautio, J.; Mattinen, L.; Vidgren, V.; Londesborough, J.; Gibson, B.R. Adaptive Evolution of the Lager Brewing Yeast Saccharomyces pastorianus for Improved Growth under Hyperosmotic Conditions and Its Influence on Fermentation Performance. FEMS Yeast Res. 2013, 13, 335–349. [Google Scholar] [CrossRef] [Green Version]
- Krogerus, K.; Holmström, S.; Gibson, B. Enhanced Wort Fermentation with De Novo Lager Hybrids. Appl. Environ. Microbiol. 2018, 84, e02302-17. [Google Scholar] [CrossRef] [Green Version]
- Christofoleti-Furlan, R.M.; Portugal, C.B.; Varize, C.S.; Muynarsk, E.S.M.; Alcarde, A.R.; Basso, L.C. Unraveling Brazilian Bioethanol Yeasts as Novel Starters for High-Gravity Brewing. Food Res. Int. 2020, 135, 109282. [Google Scholar] [CrossRef] [PubMed]
- Araújo, T.M.; Souza, M.T.; Diniz, R.H.S.; Yamakawa, C.K.; Soares, L.B.; Lenczak, J.L.; de Castro Oliveira, J.V.; Goldman, G.H.; Barbosa, E.A.; Campos, A.C.S.; et al. Cachaça Yeast Strains: Alternative Starters to Produce Beer and Bioethanol. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 1749–1766. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, V.; Radecka, D.; Aerts, G.; Verstrepen, K.J.; Lievens, B.; Thevelein, J.M. Phenotypic Landscape of Non-Conventional Yeast Species for Different Stress Tolerance Traits Desirable in Bioethanol Fermentation. Biotechnol. Biofuels 2017, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Chaves-López, C.; Di Fabio, F.; Schirone, M.; Felis, G.E.; Torriani, S.; Paparella, A.; Suzzi, G. Molecular Identification and Osmotolerant Profile of Wine Yeasts That Ferment a High Sugar Grape Must. Int. J. Food Microbiol. 2009, 130, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Meurens, N.A.A.; De Troostembergh, J.-C.M.P.; Horbach, B.; Kruse, M.J.C. Alcoholic Fermentation Process in the Presence of a High Alcohol Tolerant Yeast and a Maltotriose Positive Yeast. International Patent Application No. WO 2013/181496 Al, 5 December 2013. [Google Scholar]
- van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Abee, T.; Smid, E.J. Performance of Non-Conventional Yeasts in Co-Culture with Brewers’ Yeast for Steering Ethanol and Aroma Production. Microb. Biotechnol. 2017, 10, 1591–1602. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the Brewing Process: A New Approach to Enhance Bioflavour and to Reduce Ethanol Content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef]
- Callejo, M.J.; García Navas, J.J.; Alba, R.; Escott, C.; Loira, I.; González, M.C.; Morata, A. Wort Fermentation and Beer Conditioning with Selected Non-Saccharomyces Yeasts in Craft Beers. Eur. Food Res. Technol. 2019, 245, 1229–1238. [Google Scholar] [CrossRef]
- Benito, S. The Impacts of Schizosaccharomyces on Winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Benito, Á.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.-Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe Strains Have Characteristics That Are Beneficial for Winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderón, F.; Suárez-Lepe, J.A. New Applications for Schizosaccharomyces pombe in the Alcoholic Fermentation of Red Wines. Int. J. Food Sci. Technol. 2012, 47, 2101–2108. [Google Scholar] [CrossRef]
- Thomas, M; Method of Preparing a Low-Extract Beer. United States Patent No. 2,146,240, 3 March 1936.
- EBC ANALYTICA 3.2.1.1 Methylene Blue/Violet Stain. Available online: https://brewup.eu/ebc-analytica/yeast-analysis/methylene-blue-violet-stain/3.2.1.1 (accessed on 25 November 2021).
- Reid, S.J.; Josey, M.; Macintosh, A.J.; Maskell, D.L. Predicting Fermentation Rates in Ale, Lager and Whisky Predicting Fermentation Rates in Ale, Lager and Whisky. Fermentation 2021, 7, 13. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- EBC ANALYTICA 8.10.1 Free Amino Nitrogen in Wort by Spectrophotometry-Manual Method (IM). Available online: https://brewup.eu/ebc-analytica/wort/free-amino-nitrogen-in-wort-by-spectrophotometry-manual-method-im/8.10.1 (accessed on 26 November 2021).
- EBC ANALYTICA 8.3 Extract of Wort. Available online: https://brewup.eu/ebc-analytica/wort/extract-of-wort/8.3 (accessed on 10 November 2022).
- EBC ANALYTICA 9.5 Real Degree of Fermentation of Beer. Available online: https://brewup.eu/ebc-analytica/beer/real-degree-of-fermentation-of-beer/9.5 (accessed on 25 November 2021).
- Rollero, S.; Bloem, A.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Altered Fermentation Performances, Growth, and Metabolic Footprints Reveal Competition for Nutrients between Yeast Species Inoculated in Synthetic Grape Juice-Like Medium. Front. Microbiol. 2018, 9, 196. [Google Scholar] [CrossRef] [PubMed]
- Preiss, R.; Tyrawa, C.; Krogerus, K.; Garshol, L.M.; Van Der Merwe, G. Traditional Norwegian Kveik Are a Genetically Distinct Group of Domesticated Saccharomyces cerevisiae Brewing Yeasts. Front. Microbiol. 2018, 9, 2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daute, M.; Jack, F.; Harrison, B.; Walker, G. Experimental Whisky Fermentations: Influence of Wort Pretreatments. Foods 2021, 10, 2755. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Comitini, F.; Ciani, M. Torulaspora delbrueckii Contribution in Mixed Brewing Fermentations with Different Saccharomyces cerevisiae Strains. Int. J. Food Microbiol. 2017, 259, 7–13. [Google Scholar] [CrossRef]
- Toh, D.W.K.; Chua, J.Y.; Liu, S.Q. Impact of Simultaneous Fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii on Volatile and Non-Volatile Constituents in Beer. LWT-Food Sci. Technol. 2018, 91, 26–33. [Google Scholar] [CrossRef]
- Tonsmeire, M. Commercial Methods of Souring in American Sour Beer: Innovative Techniques for Mixed Fermentations, 1st ed.; Brewers Publications: Colorado, CO, USA, 2014; pp. 228–285. [Google Scholar]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The Soul of Beer’s Aroma-A Review of Flavour-Active Esters and Higher Alcohols Produced by the Brewing Yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Hazelwood, L.A.; Daran, J.M.; Van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Omura, F.; Miyajima, K.; Ashikari, T. Control of Higher Alcohol Production by Manipulation of the BAP2 Gene in Brewing Yeast. J. Am. Soc. Brew. Chem. 2001, 59, 157–162. [Google Scholar] [CrossRef]
- Lilly, M.; Bauer, F.F.; Styger, G.; Lambrechts, M.G.; Pretorius, I.S. The Effect of Increased Branched-Chain Amino Acid Transaminase Activity in Yeast on the Production of Higher Alcohols and on the Flavour Profiles of Wine and Distillates. FEMS Yeast Res. 2006, 6, 726–743. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.; Caldeira, M.; Câmara, J.S. Development of a Dynamic Headspace Solid-Phase Microextraction Procedure Coupled to GC-QMSD for Evaluation the Chemical Profile in Alcoholic Beverages. Anal. Chim. Acta 2008, 609, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miks, M.H.; De Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The Molecular Biology of Fruity and Floral Aromas in Beer and Other Alcoholic Beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [Green Version]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-Active Volatile Compounds in Beer: Production, Regulation and Control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Meilgaard, M.C. Flavor Chemistry of Beer: Part II: Flavor and Threshold of 239 Aroma Volatiles. MBAA Technol. Q. 1975, 12, 151–168. [Google Scholar]
- Piddocke, M.P.; Kreisz, S.; Heldt-Hansen, H.P.; Nielsen, K.F.; Olsson, L. Physiological Characterization of Brewer’s Yeast in High-Gravity Beer Fermentations with Glucose or Maltose Syrups as Adjuncts. Appl. Microbiol. Biotechnol. 2009, 84, 453–464. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, S.; Heponiemi, P.; Heinonen, M.; Marsol-Vall, A.; Ma, X.; Yang, B.; Laaksonen, O. Effect of Saccharomyces cerevisiae and Schizosaccharomyces pombe Strains on Chemical Composition and Sensory Quality of Ciders Made from Finnish Apple Cultivars. Food Chem. 2021, 345, 128833. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii Strains in Mixed and Sequential Fermentations to Improve Red Wine Sensory Quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Meier-Dörnberg, T.; Kory, O.I.; Jacob, F.; Michel, M.; Hutzler, M. Saccharomyces cerevisiae Variety Diastaticus Friend or Foe?-Spoilage Potential and Brewing Ability of Different Saccharomyces cerevisiae Variety Diastaticus Yeast Isolates by Genetic, Phenotypic and Physiological Characterization. FEMS Yeast Res. 2018, 18, foy023. [Google Scholar] [CrossRef] [Green Version]
- Tubb, R.S.; Searle, B.A.; Goodey, A.R.; Brown, A.J.P. Rare Mating and Transformation for Construction of Novel Brewing Yeasts. In Proceedings of the 18th Congress of the European Brewery Convention, Brussels, Belgium, 1981; pp. 487–496. [Google Scholar]
- Krogerus, K.; Seppänen-Laakso, T.; Castillo, S.; Gibson, B. Inheritance of Brewing-Relevant Phenotypes in Constructed Saccharomyces cerevisiae × Saccharomyces eubayanus Hybrids. Microb. Cell Fact. 2017, 16, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Younis, O.S.; Stewart, G.G. The Science of Beer Effect of Malt Wort, Very-High-Gravity Malt Wort, and Very-High-Gravity Adjunct Wort on Volatile Production in Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2018, 57, 39–45. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ratio (SCN:SZ1) | Lag Phase (h) | End of Exponential Phase (h) | Maximum Rate of Fermentation (g/h) |
|---|---|---|---|
| 1:0 | 12.6 ± 0.3 a | 31.7 ± 0.7 ab | 0.099 ± 0.003 a |
| 20:1 | 12.6 ± 0.7 a | 31.2 ± 1.0 a | 0.102 ± 0.003 ab |
| 1:1 | 14.1 ± 0.7 a | 32.1 ± 1.5 ab | 0.107 ± 0.004 bc |
| 1:20 | 17.1 ± 0.7 b | 34.4 ± 1.0 b | 0.113 ± 0.001 cd |
| 0:1 | 18.1 ± 0.3 b | 34.4 ± 0.8 b | 0.119 ± 0.002 d |
| Ratio (SCN:SZ1) | Residual FAN (mg/L) | FAN Consumption (%) | pH | FG | RDF (%) | ABV (v/v %) |
|---|---|---|---|---|---|---|
| 1:0 | 49 ± 1 a | 76.3 ± 0.6 b | 4.16 ± 0.03 b | 1.00632 ± 0.00003 c | 84.8 ± 0.2 a | 5.56 ± 0.08 a |
| 20:1 | 44 ± 1 a | 78.7 ± 0.4 b | 4.08 ± 0.02 ab | 1.00516 ± 0.00008 b | 87.3 ± 0.2 b | 5.59 ± 0.02 a |
| 1:1 | 46 ± 4 a | 77.7 ± 2.0 b | 3.93 ± 0.11 a | 1.00443 ± 0.00007 a | 89.1 ± 0.2 d | 5.74 ± 0.12 a |
| 1:20 | 47 ± 5 a | 77.5 ± 2.3 b | 4.21 ± 0.04 b | 1.00442 ± 0.00015 a | 88.9 ± 0.1 d | 5.62 ± 0.13 a |
| 0:1 | 62 ± 2 b | 70.2 ± 0.9 a | 4.12 ± 0.03 b | 1.00496 ± 0.00010 b | 87.8 ± 0.2 c | 5.65 ± 0.01 a |
| Ratio (SCN:SZ1) | 1:0 | 20:1 | 1:1 | 1:20 | 0:1 | Aroma Threshold (mg/L) | |
|---|---|---|---|---|---|---|---|
| Compound | Concentration (mg/L) | Reference | |||||
| Acetaldehyde | 3.15 ± 1.10 a | 3.42 ± 0.22 a | 22.0 ± 20.6 a | 20.6 ± 25.7 a | 21.0 ± 4.0 a | 25 | [42] |
| Methanol | 7.50 ± 0.98 a | 6.95 ± 0.64 a | 28.1 ± 22.6 a | 8.01 ± 4.27 a | 4.14 ± 0.86 a | 10,000 | [43] |
| Propan-1-ol | 17.9 ± 4.2 b | 9.51 ± 1.86 ab | 6.00 ± 8.01 ab | 4.88 ± 4.23 a | ND a | 600 | [36] |
| Butan-1-ol | 0.151 ± 0.023 a | 0.150 ± 0.015 a | 0.203 ± 0.066 a | 0.136 ± 0.028 a | 0.0807 ± 0.0699 a | 450 | [43] |
| Isobutanol | 19.7 ± 2.0 b | 17.8 ± 1.0 b | 31.4 ± 5.2 c | 20.2 ± 4.4 b | 1.75 ± 0.27 a | 100 | [36] |
| Isoamyl alcohol | 36.2 ± 4.3 b | 31.4 ± 1.9 b | 64.2 ± 11.6 c | 40.5 ± 5.4 b | 6.13 ± 1.37 a | 50–65 | [36] |
| 2-phenylethyl alcohol | ND a | ND a | 26.0 ± 24.2 a | ND a | ND a | 40 | [36] |
| Isoamyl acetate | 2.95 ± 0.33 a | 3.39 ± 0.19 ab | 3.91 ± 0.30 b | 3.21 ± 0.07 a | 3.83 ± 0.15 b | 1.2–2.0 | [36] |
| Ethyl acetate | 3.54 ± 0.99 a | 2.35 ± 0.29 a | 4.36 ± 1.02 a | 4.46 ± 2.55 a | 4.80 ± 1.50 a | 25–30 | [36] |
| Phenylethyl acetate | ND | ND | ND | ND | ND | 0.2–3.8 | [36] |
| Ethyl hexanoate | 0.102 ± 0.053 a | 0.143 ± 0.071 a | 0.0835 ± 0.0258 a | 0.0572 ± 0.0135 a | 0.147 ± 0.076 a | 0.2–0.23 | [36] |
| Ethyl octanoate | 0.0364 ± 0.0235 a | 0.0673 ± 0.0203 a | 0.122 ± 0.105 a | 0.0357 ± 0.0219 a | 0.0468 ± 0.0031 a | 0.9–1.0 | [36] |
| Ethyl lactate | 0.954 ± 0.831 a | 1.16 ± 1.07 a | 1.14 ± 1.97 a | 0.104 ± 0.179 a | 1.11 ± 1.14 a | 154 | [43] |
| Ratio (SCN:SZ1) | Lag Time (h) | End of Exponential Phase (h) | Maximum Rate of Fermentation (g/h) |
|---|---|---|---|
| 1:0 | 6.3 ± 0.3 a | 41.7 ± 0.8 a | 0.088 ± 0.001 c |
| 1:1 | 10.1 ± 0.4 b | 57.0 ± 1.7 b | 0.082 ± 0.001 b |
| 0:1 | 13.1 ± 0.4 c | 87.8 ± 1.9 c | 0.054 ± 0.000 a |
| Ratio (SCN:SZ1) | Residual FAN (mg/L) | FAN Consumption (%) | pH | FG | RDF (%) | ABV (v/v %) |
|---|---|---|---|---|---|---|
| 1:0 | 46 ± 1 a | 77.7 ± 0.4 a | 3.91 ± 0.07 b | 1.02308 ± 0.00057 b | 69.8 ± 0.5 a | 8.46 ± 0.03 a |
| 1:1 | 41 ± 2 a | 80.0 ± 1.1 a | 3.55 ± 0.02 a | 1.00975 ± 0.00051 a | 87.0 ± 0.6 b | 10.32 ± 0.02 b |
| 0:1 | 47 ± 1 a | 77.1 ± 0.3 a | 3.43 ± 0.01 a | 1.00858 ± 0.00220 a | 88.5 ± 2.7 b | 10.45 ± 0.09 b |
| Ratio (SCN:SZ1) | 1:0 | 1:1 | 0:1 | Aroma Threshold (mg/L) | |
|---|---|---|---|---|---|
| Compound | Concentration (mg/L) | Reference | |||
| Acetaldehyde | 9.98 ± 8.58 a | 6.81 ± 1.43 a | 7.68 ± 1.07 a | 25 | [42] |
| Methanol | 45.9 ± 7.4 a | 66.0 ± 8.2 b | 42.6 ± 3.2 a | 10,000 | [43] |
| Propan-1-ol | ND a | ND a | 4.77 ± 5.80 a | 600 | [36] |
| Butan-1-ol | 11.1 ± 0.0 a | 12.8 ± 0.5 b | 11.8 ± 0.2 a | 450 | [43] |
| Isobutanol | 62.3 ± 5.1 b | 59.4 ± 8.9 b | 18.1 ± 1.6 a | 100 | [36] |
| Isoamyl alcohol | 145 ± 11 b | 280 ± 45 c | 30.7 ± 2.6 a | 50–65 | [36] |
| 2-phenylethyl alcohol | ND | ND | ND | 40 | [36] |
| Isoamyl acetate | 1.58 ± 0.02 a | 2.06 ± 0.11 b | 1.54 ± 0.02 a | 1.2–2.0 | [36] |
| Ethyl acetate | 6.80 ± 1.34 a | 9.07 ± 1.37 a | 6.18 ± 1.09 a | 25–30 | [36] |
| Phenylethyl acetate | 0.140 ± 0.016 a | 0.369 ± 0.062 b | 0.0394 ± 0.0682 a | 0.2–3.8 | [36] |
| Ethyl hexanoate | 0.194 ± 0.030 a | 0.210 ± 0.066 a | 0.150 ± 0.006 a | 0.2–0.23 | [36] |
| Ethyl octanoate | 0.159 ± 0.001 a | 0.174 ± 0.003 b | 0.171 ± 0.005 b | 0.9–1.0 | [36] |
| Ethyl lactate | 11.7 ± 10.4 a | 7.73 ± 13.40 a | 5.52 ± 9.57 a | 154 | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pownall, B.; Reid, S.J.; Hill, A.E.; Jenkins, D. Schizosaccharomyces pombe in the Brewing Process: Mixed-Culture Fermentation for More Complete Attenuation of High-Gravity Wort. Fermentation 2022, 8, 643. https://doi.org/10.3390/fermentation8110643
Pownall B, Reid SJ, Hill AE, Jenkins D. Schizosaccharomyces pombe in the Brewing Process: Mixed-Culture Fermentation for More Complete Attenuation of High-Gravity Wort. Fermentation. 2022; 8(11):643. https://doi.org/10.3390/fermentation8110643
Chicago/Turabian StylePownall, Barnaby, Struan J. Reid, Annie E. Hill, and David Jenkins. 2022. "Schizosaccharomyces pombe in the Brewing Process: Mixed-Culture Fermentation for More Complete Attenuation of High-Gravity Wort" Fermentation 8, no. 11: 643. https://doi.org/10.3390/fermentation8110643