
Thorough Investigation of the Effects of Cultivation Factors on Polyhydroalkanoates (PHAs) Production by Cupriavidus necator from Food Waste-Derived Volatile Fatty Acids
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Volatile Fatty Acids from Acidogenic Fermentation of Food Waste
2.3. Bacterial Cultivation
2.4. Analytical Methods
2.5. Kinetics Analysis of Biomass Production and PHA Accumulation
2.6. Characterisation of PHA
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effect of Individual VFA Concentrations on Bacterial Growth
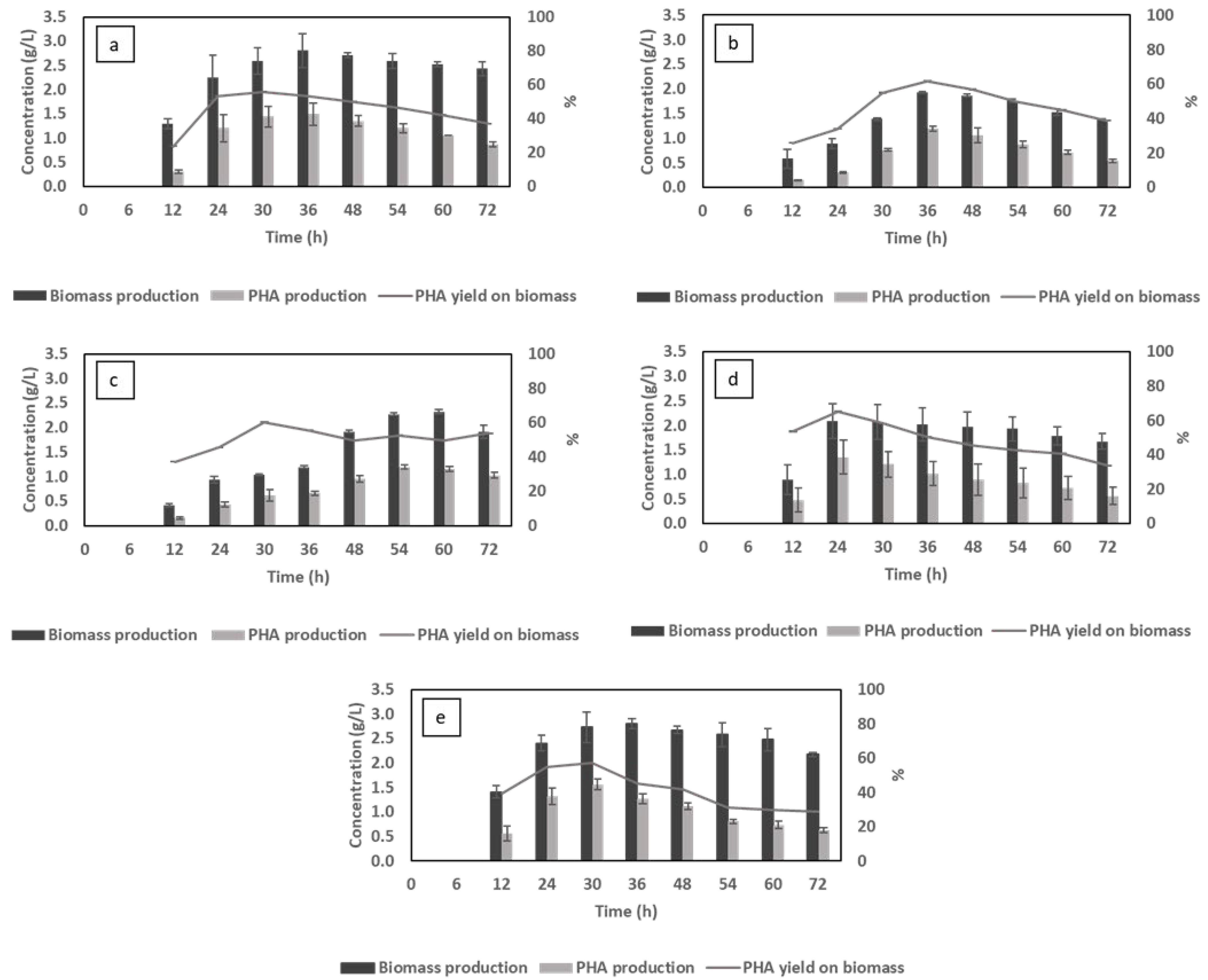
3.2. Effect of Different Cultivation Factors on Biomass and PHA Production
3.2.1. The Effect of pH
3.2.2. The Effect of Aeration Rate
3.2.3. The Effect of C/N Ratio
3.2.4. The Effect of Nitrogen Source
3.3. C. necator Cultivation on VFAs in CSTR
3.3.1. Yields of Biomass Production and PHA Accumulation
3.3.2. PHA Characterisation
Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
Nuclear Magnetic Resonance (NMR) Analysis
3.3.3. Thermogravimetric and Differential Scanning Calorimetry Analysis (TGA and DSC)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skoczinski, P.; Krause, L.; Raschka, A.; Dammer, L.; Carus, M. Chapter One-Current Status and Future Development of Plastics: Solutions for a Circular Economy and Limitations of Environmental Degradation. In Methods in Enzymology; Weber, G., Bornscheuer, U.T., Wei, R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 1–26. [Google Scholar]
- Li, P.; Wang, X.; Su, M.; Zou, X.; Duan, L.; Zhang, H. Characteristics of plastic pollution in the environment: A review. Bull. Environ. Contam. Toxicol. 2021, 107, 577–584. [Google Scholar] [CrossRef]
- Vu, D.H.; Åkesson, D.; Taherzadeh, M.J.; Ferreira, J.A. Recycling strategies for polyhydroxyalkanoate-based waste materials: An overview. Bioresour. Technol. 2020, 298, 122393. [Google Scholar] [CrossRef]
- Adeleye, A.T.; Odoh, C.K.; Enudi, O.C.; Banjoko, O.O.; Osiboye, O.O.; Odediran, E.T.; Louis, H. Sustainable synthesis and applications of polyhydroxyalkanoates (phas) from biomass. Process Biochem. 2020, 96, 174–193. [Google Scholar] [CrossRef]
- Raza, Z.A.; Abid, S.; Banat, I.M. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. Int. Biodeterior. Biodegrad. 2018, 126, 45–56. [Google Scholar] [CrossRef]
- De Donno Novelli, L.; Sayavedra, S.M.; Rene, E.R. Polyhydroxyalkanoate (pha) production via resource recovery from industrial waste streams: A review of techniques and perspectives. Bioresour. Technol. 2021, 331, 124985. [Google Scholar] [CrossRef]
- Koller, M.; Atlić, A.; Dias, M.; Reiterer, A.; Braunegg, G. Microbial PHA Production from Waste Raw Materials. In Plastics from Bacteria: Natural Functions and Applications; Chen, G.G.-Q., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 85–119. [Google Scholar]
- Silva, L.F.; Taciro, M.K.; Ramos, M.E.M.; Carter, J.M.; Pradella, J.G.; Gomez, J.G. Poly-3-hydroxybutyrate (p3hb) production by bacteria from xylose, glucose and sugarcane bagasse hydrolysate. J. Ind. Microbiol. Biotechnol. 2004, 31, 245–254. [Google Scholar] [CrossRef]
- Solaiman, D.K.Y.; Ashby, R.D.; Foglia, T.A.; Marmer, W.N. Conversion of agricultural feedstock and coproducts into poly(hydroxyalkanoates). Appl. Microbiol. Biotechnol. 2006, 71, 783–789. [Google Scholar] [CrossRef]
- Raza, Z.A.; Tariq, M.R.; Majeed, M.I.; Banat, I.M. Recent developments in bioreactor scale production of bacterial polyhydroxyalkanoates. Bioprocess Biosyst. Eng. 2019, 42, 901–919. [Google Scholar] [CrossRef]
- Davis, R.; Kataria, R.; Cerrone, F.; Woods, T.; Kenny, S.; O’Donovan, A.; Guzik, M.; Shaikh, H.; Duane, G.; Gupta, V.K.; et al. Conversion of grass biomass into fermentable sugars and its utilization for medium chain length polyhydroxyalkanoate (mcl-pha) production by pseudomonas strains. Bioresour. Technol. 2013, 150, 202–209. [Google Scholar] [CrossRef]
- Ruiz, C.; Kenny, S.T.; P, R.B.; Walsh, M.; Narancic, T.; O’Connor, K.E. High cell density conversion of hydrolysed waste cooking oil fatty acids into medium chain length polyhydroxyalkanoate using pseudomonas putida kt2440. Catalysts 2019, 9, 468. [Google Scholar] [CrossRef]
- Harirchi, S.; Wainaina, S.; Sar, T.; Nojoumi, S.A.; Parchami, M.; Parchami, M.; Varjani, S.; Khanal, S.K.; Wong, J.; Awasthi, M.K.; et al. Microbiological insights into anaerobic digestion for biogas, hydrogen or volatile fatty acids (vfas): A review. Bioengineered 2022, 13, 6521–6557. [Google Scholar] [CrossRef]
- Wainaina, S.; Awasthi, M.K.; Sarsaiya, S.; Chen, H.; Singh, E.; Kumar, A.; Ravindran, B.; Awasthi, S.K.; Liu, T.; Duan, Y.; et al. Resource recovery and circular economy from organic solid waste using aerobic and anaerobic digestion technologies. Bioresour. Technol. 2020, 301, 122778. [Google Scholar] [CrossRef]
- Ramos-Suarez, M.; Zhang, Y.; Outram, V. Current perspectives on acidogenic fermentation to produce volatile fatty acids from waste. Rev. Environ. Sci. Bio/Technol. 2021, 20, 439–478. [Google Scholar] [CrossRef]
- Liu, C.C.; Zhang, L.L.; An, J.; Chen, B.; Yang, H. Recent strategies for efficient production of polyhydroxyalkanoates by micro-organisms. Lett. Appl. Microbiol. 2016, 62, 9–15. [Google Scholar] [CrossRef]
- Agustín Martinez, G.; Bertin, L.; Scoma, A.; Rebecchi, S.; Braunegg, G.; Fava, F. Production of polyhydroxyalkanoates from dephenolised and fermented olive mill wastewaters by employing a pure culture of Cupriavidus necator. Biochem. Eng. J. 2015, 97, 92–100. [Google Scholar] [CrossRef]
- Cheah, Y.-K.; Vidal-Antich, C.; Dosta, J.; Mata-Álvarez, J. Volatile fatty acid production from mesophilic acidogenic fermentation of organic fraction of municipal solid waste and food waste under acidic and alkaline ph. Environ. Sci. Pollut. Res. Int. 2019, 26, 35509–35522. [Google Scholar] [CrossRef] [Green Version]
- Kachrimanidou, V.; Ioannidou, S.M.; Ladakis, D.; Papapostolou, H.; Kopsahelis, N.; Koutinas, A.A.; Kookos, I.K. Techno-economic evaluation and life-cycle assessment of poly(3-hydroxybutyrate) production within a biorefinery concept using sunflower-based biodiesel industry by-products. Bioresour. Technol. 2021, 326, 124711. [Google Scholar] [CrossRef]
- Pervez, M.N.; Bilgiç, B.; Mahboubi, A.; Uwineza, C.; Zarra, T.; Belgiorno, V.; Naddeo, V.; Taherzadeh, M.J. Double-stage membrane-assisted anaerobic digestion process intensification for production and recovery of volatile fatty acids from food waste. Sci. Total Environ. 2022, 825, 154084. [Google Scholar] [CrossRef]
- Reddy Prasad, D.M.; Pendyala, R.; Senthilkumar, R.; Azri, M.H.B. Microbial production of poly (3-hydroxybutyrate) (phb) from rubber seed oil using Cupriavidus necator h16. IOP Conf. Ser. Earth Environ. Sci. 2019, 398, 012008. [Google Scholar] [CrossRef]
- Sukruansuwan, V.; Napathorn, S. Use of agro-industrial residue from the canned pineapple industry for polyhydroxybutyrate production by Cupriavidus necator strain a-04. Biotechnol. Biofuels 2018, 11, 202. [Google Scholar] [CrossRef]
- Du, G.; Si, Y.; Yu, J. Inhibitory effect of medium-chain-length fatty acid on synthesis of polyhydroxyalkanoates from volatile fatty acid by ralstonia eutrophus. Biotechnol. Lett. 2001, 23, 1613–1617. [Google Scholar] [CrossRef]
- Yun, J.; Sawant, S.; Kim, B.S. Production of polyhydroxyalkanoates by ralstonia eutropha from volatile fatty acids. Korean J. Chem. Eng. 2013, 30, 2223–2227. [Google Scholar] [CrossRef]
- Vu, D.H.; Wainaina, S.; Taherzadeh, M.J.; Åkesson, D.; Ferreira, J.A. Production of polyhydroxyalkanoates (phas) by bacillus megaterium using food waste acidogenic fermentation-derived volatile fatty acids. Bioengineered 2021, 12, 2480–2498. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.H.; Kummerlöwe, C.; Kammer, H.-W. Crystallization and melting behavior of poly(3-hydroxybutyrate)-based blends. Macromol. Chem. Phys. 2004, 205, 664–675. [Google Scholar] [CrossRef]
- Jaramillo-Sánchez, R.; Alcaraz-Zapata, W. Limitations on production methods for phas obtention: A systematic review. DYNA 2020, 87, 193–203. [Google Scholar]
- Wilbanks, B.; Trinh, C.T. Comprehensive characterization of toxicity of fermentative metabolites on microbial growth. Biotechnol. Biofuels 2017, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Aremu, M.O.; Ishola, M.M.; Taherzadeh, M.J. Polyhydroxyalkanoates (phas) production from volatile fatty acids (vfas) from organic wastes by pseudomonas oleovorans. Fermentation 2021, 7, 287. [Google Scholar] [CrossRef]
- Setiadi, T.; Aznury, M.; Trianto, A.; Pancoro, A. Production of polyhydroxyalkanoate (pha) by ralstonia eutropha jmp 134 with volatile fatty acids from palm oil mill effluent as precursors. Water Sci. Technol. 2015, 72, 1889–1895. [Google Scholar] [CrossRef]
- Minocha, S.C. PH of the Medium and the Growth and Metabolism of Cells in Culture. In Cell and Tissue Culture in Forestry: General Principles and Biotechnology; Bonga, J.M., Durzan, D.J., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 125–141. [Google Scholar]
- Shehadul Islam, M.; Aryasomayajula, A.; Selvaganapathy, P.R. A review on macroscale and microscale cell lysis methods. Micromachines 2017, 8, 83. [Google Scholar] [CrossRef]
- Jin, Q.; Kirk, M.F. PH as a primary control in environmental microbiology: 1. Thermodynamic perspective. Front. Environ. Sci. 2018, 6, 21. [Google Scholar] [CrossRef]
- Sánchez-Clemente, R.; Igeño, M.I.; Población, A.G.; Guijo, M.I.; Merchán, F.; Blasco, R. Study of ph changes in media during bacterial growth of several environmental strains. Proceedings 2018, 2, 1297. [Google Scholar]
- Ratzke, C.; Gore, J. Modifying and reacting to the environmental ph can drive bacterial interactions. PLoS Biol. 2018, 16, e2004248. [Google Scholar] [CrossRef] [PubMed]
- Aramvash, A.; Shahabi, Z.A.; Aghjeh, S.D.; Ghafari, M.D. Statistical physical and nutrient optimization of bioplastic polyhydroxybutyrate production by Cupriavidus necator. Int. J. Environ. Sci. Technol. 2015, 12, 2307–2316. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, D.; Yashchuk, O.; Hermida, É.B. Evaluation of culture medium on poly(3-hydroxybutyrate) production by Cupriavidus necator atcc 17697: Application of the response surface methodology. Heliyon 2019, 5, e01374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, R.A.P.; Oehmen, A.; Reis, M.A.M. The impact of biomass withdrawal strategy on the biomass selection and polyhydroxyalkanoates accumulation of mixed microbial cultures. New Biotechnol. 2022, 66, 8–15. [Google Scholar] [CrossRef]
- Smith, B. How Does PH Level Affect Enzyme Activity? 2017. Available online: https://sciencing.com/role-enzymes-chemical-reactions-5553131.html (accessed on 15 September 2022).
- Foundation, N. Bacterial Responses to PH.; Wiley: Hoboken, NJ, USA, 2008. [Google Scholar]
- Catherine, M.-C.; Guwy, A.; Massanet-Nicolau, J. Effect of acetate concentration, temperature, ph and nutrient concentration on polyhydroxyalkanoates (pha) production by glycogen accumulating organisms. Bioresour. Technol. Rep. 2022, 20, 101226. [Google Scholar] [CrossRef]
- Matsakas, A.; Patel, K. Aerobic Metabolism. In Encyclopedia of Exercise Medicine in Health and Disease; Mooren, F.C., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 29–32. [Google Scholar]
- Gao, J.; Ramsay, J.A.; Ramsay, B.A. Fed-batch production of poly-3-hydroxydecanoate from decanoic acid. J. Biotechnol. 2016, 218, 102–107. [Google Scholar] [CrossRef]
- Wang, X.; Oehmen, A.; Freitas, E.B.; Carvalho, G.; Reis, M.A. The link of feast-phase dissolved oxygen (do) with substrate competition and microbial selection in pha production. Water Res. 2017, 112, 269–278. [Google Scholar] [CrossRef]
- Lefebvre, G.; Rocher, M.; Braunegg, G. Effects of low dissolved-oxygen concentrations on poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) production by alcaligenes eutrophus. Appl. Environ. Microbiol. 1997, 63, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Shantini, K.; Yahya, A.R.; Amirul, A.A. Influence of feeding and controlled dissolved oxygen level on the production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymer by cupriavidus sp. Usmaa2-4 and its characterization. Appl. Biochem. Biotechnol. 2015, 176, 1315–1334. [Google Scholar] [CrossRef]
- Cavalheiro, J.M.B.T.; Raposo, R.S.; de Almeida, M.C.M.D.; Cesário, M.T.; Sevrin, C.; Grandfils, C.; da Fonseca, M.M.R. Effect of cultivation parameters on the production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) and poly(3-hydroxybutyrate-4-hydroxybutyrate-3-hydroxyvalerate) by Cupriavidus necator using waste glycerol. Bioresour. Technol. 2012, 111, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K.; Kettl, K.-H.; Titz, M.; Koller, M.; Schnitzer, H.; Narodoslawsky, M. Comparison of ecological footprint for biobased pha production from animal residues utilizing different energy resources. Clean Technol. Environ. Policy 2013. 15, 525–536. [CrossRef]
- Zhang, C.C.; Zhou, C.Z.; Burnap, R.L.; Peng, L. Carbon/nitrogen metabolic balance: Lessons from cyanobacteria. Trends Plant Sci. 2018, 23, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Muhammadi; Shabina; Afzal, M.; Hameed, S. Bacterial polyhydroxyalkanoates-eco-friendly next generation plastic: Production, biocompatibility, biodegradation, physical properties and applications. Green Chem. Lett. Rev. 2015, 8, 56–77. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.S.S.; Silva, C.E.; Carvalho, G.; Reis, M.A. Strategies for efficiently selecting pha producing mixed microbial cultures using complex feedstocks: Feast and famine regime and uncoupled carbon and nitrogen availabilities. New Biotechnol. 2017, 37, 69–79. [Google Scholar] [CrossRef]
- Zhou, W.; Colpa, D.I.; Geurkink, B.; Euverink, G.-J.W.; Krooneman, J. The impact of carbon to nitrogen ratios and ph on the microbial prevalence and polyhydroxybutyrate production levels using a mixed microbial starter culture. Sci. Total Environ. 2022, 811, 152341. [Google Scholar] [CrossRef]
- Sangkharak, K.; Prasertsan, P. Nutrient optimization for production of polyhydroxybutyrate from halotolerant photosynthetic bacteria cultivated under aerobic-dark condition. Electron. J. Biotechnol. 2008, 11, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Sarkar, B.; Samantaray, D.P.; Daware, A.; Maity, S.; Pattnaik, S.; Bhattacharjee, S. Bioconversion of fish solid waste into phb using bacillus subtilis based submerged fermentation process. Environ. Technol. 2017, 38, 3201–3208. [Google Scholar] [CrossRef]
- Ahn, J.; Jho, E.H.; Nam, K. Effect of c/n ratio on polyhydroxyalkanoates (pha) accumulation by Cupriavidus necator and its implication on the use of rice straw hydrolysates. Environ. Eng. Res. 2015, 20, 246–253. [Google Scholar] [CrossRef]
- Budde, C.F.; Riedel, S.L.; Hübner, F.; Risch, S.; Popović, M.K.; Rha, C.; Sinskey, A.J. Growth and polyhydroxybutyrate production by ralstonia eutropha in emulsified plant oil medium. Appl. Microbiol. Biotechnol. 2011. 89, 1611–1619. [CrossRef]
- da Cruz Pradella, J.G.; Ienczak, J.L.; Delgado, C.R.; Taciro, M.K. Carbon source pulsed feeding to attain high yield and high productivity in poly(3-hydroxybutyrate) (phb) production from soybean oil using Cupriavidus necator. Biotechnol. Lett. 2012, 34, 1003–1007. [Google Scholar] [CrossRef]
- Benesova, P.; Kucera, D.; Marova, I.; Obruca, S. Chicken feather hydrolysate as an inexpensive complex nitrogen source for pha production by Cupriavidus necator on waste frying oils. Lett. Appl. Microbiol. 2017, 65, 182–188. [Google Scholar] [CrossRef]
- Koller, M.; Bona, R.; Hermann, C.; Horvat, P.; Martinz, J.; Neto, J.; Pereira, L.; Varila, P.; Braunegg, G. Biotechnological production of poly(3-hydroxybutyrate) with wautersia eutropha by application of green grass juice and silage juice as additional complex substrates. Biocatal. Biotransformation 2005, 23, 329–337. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Oborna, J.; Marova, I. Application of protease-hydrolyzed whey as a complex nitrogen source to increase poly(3-hydroxybutyrate) production from oils by Cupriavidus necator. Biotechnol. Lett. 2014, 36, 775–781. [Google Scholar] [CrossRef]
- Arumugam, A.; Senthamizhan, S.G.; Ponnusami, V.; Sudalai, S. Production and optimization of polyhydroxyalkanoates from non-edible calophyllum inophyllum oil using Cupriavidus necator. Int. J. Biol. Macromol. 2018, 112, 598–607. [Google Scholar] [CrossRef]
- Dañez, J.C.A.; Requiso, P.J.; Alfafara, C.G.; Nayve, F.R.P. Optimization of fermentation factors for polyhydroxybutyrate (phb) production using bacillus megaterium pncm 1890 in simulated glucose-xylose hydrolysates from agricultural residues. Philipp. J. Sci. 2020, 149, 163–175. [Google Scholar]
- Loo, Y.C.; Sudesh, K. Polyhydroxyalkanoates: Bio-based microbial plastics and their properties. Malays. Polym. J. (MPJ) 2007, 2, 31–57. [Google Scholar]
- Riedel, L.S.; Bader, J.; Brigham, C.J.; Budde, C.F.; Yusof, Z.A.; Rha, C.; Sinskey, A.J. Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) by ralstonia eutropha in high cell density palm oil fermentations. Biotechnol. Bioeng. 2012, 109, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Kulpreecha, S.; Boonruangthavorn, A.; Meksiriporn, B.; Thongchul, N. Inexpensive fed-batch cultivation for high poly(3-hydroxybutyrate) production by a new isolate of bacillus megaterium. J. Biosci. Bioeng. 2009, 107, 240–245. [Google Scholar] [CrossRef]
- Kumari, A. Chapter 4-beta oxidation of fatty acids. In Sweet Biochemistry; Kumari, A., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 17–19. [Google Scholar]
- Randriamahefa, S.; Renard, E.; Guérin, P.; Langlois, V. Fourier transform infrared spectroscopy for screening and quantifying production of phas by pseudomonas grown on sodium octanoate. Biomacromolecules 2003, 4, 1092–1097. [Google Scholar] [CrossRef]
- Trakunjae, C.; Boondaeng, A.; Apiwatanapiwat, W.; Kosugi, A.; Arai, T.; Sudesh, K.; Vaithanomsat, P. Enhanced polyhydroxybutyrate (phb) production by newly isolated rare actinomycetes rhodococcus sp. Strain bsrt1-1 using response surface methodology. Sci. Rep. 2021, 11, 1896. [Google Scholar]
- Bluhm, T.L.; Hamer, G.K.; Marchessault, R.H.; Fyfe, C.A.; Veregin, R.P. Isodimorphism in bacterial poly(β-hydroxybutyrate-co-β-hydroxyvalerate). Macromolecules 1986. 19, 2871–2876. [CrossRef]
- Kamiya, N.; Sakurai, M.; Inoue, Y.; Chujo, R.; Doi, Y. Study of cocrystallization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by solid-state high-resolution carbon-13 nmr spectroscopy and differential scanning calorimetry. Macromolecules 1991, 24, 2178–2182. [Google Scholar] [CrossRef]
- William, J.O.; David, L.V.; Terry, L.B.; Robert, H.M. Cocrystallization in random copolymers of poly(β-hydroxybutyrate-co-β-hydroxyvalerate) and its effect on crystalline morphology. Can. J. Chem. 1995, 73, 2094–2100. [Google Scholar]
- VanderHart, D.L.; Orts, W.J.; Marchessault, R.H. 13c nmr determination of the degree of cocrystallization in random copolymers of poly(Beta.-hydroxybutyrate-co-.Beta.-hydroxyvalerate). Macromolecules 1995, 28, 6394–6400. [Google Scholar]
- Bonthrone, K.M.; Clauss, J.; Horowitz, D.M.; Hunter, B.K.; Sanders, J.K.M. The biological and physical chemistry of polyhydroxyalkanoates as seen by nmr spectroscopy. FEMS Microbiol. Lett. 1992, 103, 269–277. [Google Scholar] [CrossRef]
- Ntaikou, I.; Koumelis, I.; Kamilari, M.; Iatridi, Z.; Tsitsilianis, C.; Lyberatos, G. Effect of nitrogen limitation on polyhydroxyalkanoates production efficiency, properties and microbial dynamics using a soil-derived mixed continuous culture. Int. J. Biobased Plast. 2019, 1, 31–47. [Google Scholar] [CrossRef]
- Rodrigues, P. Impact of different bacterial strains on the production, composition, and properties of novel polyhydroxyalkanoates using crude palm oil as substrate. Chem. Biochem. Eng. Q. 2018, 32, 141–150. [Google Scholar] [CrossRef]
- Dietrich, K.; Dumont, M.-J.; del Rio, L.F.; Orsat, V. Producing phas in the bioeconomy—towards a sustainable bioplastic. Sustain. Prod. Consum. 2017, 9, 58–70. [Google Scholar] [CrossRef]
- Nygaard, D.; Yashchuk, O.; Noseda, D.G.; Araoz, B.; Hermida, É.B. Improved fermentation strategies in a bioreactor for enhancing poly(3-hydroxybutyrate) (phb) production by wild type Cupriavidus necator from fructose. Heliyon 2021, 7, e05979. [Google Scholar] [CrossRef]
- Li, D.; Li, J.; Ma, X. Accumulation of bioplastic polyhydroxyalkanoate with different substrate forms from pretreated waste lignocellulose hydrolysate. Ind. Crops Prod. 2021, 172, 114061. [Google Scholar] [CrossRef]
- Rosengart, A.; Cesário, M.T.; de Almeida, M.C.M.D.; Raposo, R.S.; Espert, A.; de Apodaca, E.D.; da Fonseca, M.M.R. Efficient p(3hb) extraction from burkholderia sacchari cells using non-chlorinated solvents. Biochem. Eng. J. 2015, 103, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Scattering methods and the properties of polymer materials. Mater. Today 2005, 8, 59. [CrossRef]
- Nair, A.M.; Annamalai, K.; Kannan, S.K.; Kuppusamy, S. Characterization of polyhydroxyalkanoates produced by bacillus subtilis isolated from soil samples. Malaya J. Biosci. 2014, 1, 8–12. [Google Scholar]
- Zhila, N.O.; Sapozhnikova, K.Y.; Kiselev, E.G.; Vasiliev, A.D.; Nemtsev, I.V.; Shishatskaya, E.I.; Volova, T.G. Properties of degradable polyhydroxyalkanoates (phas) synthesized by a new strain, Cupriavidus necator ibp/sfu-1, from various carbon sources. Polymers 2021, 13, 3142. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejski, W.; Klinowski, J. Kinetics of Cross-Polarization in Solid-State NMR: A Guide for Chemists. Chem. Rev. 2002, 102, 613–628. [Google Scholar] [CrossRef]
- Fitted Using the Python curve_fit Routine, Version 2.7, Python Software Foundation. Available online: www.python.org (accessed on 9 October 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (g/L) | |
|---|---|
| Acetic acid | 1.72 |
| Propionic acid | 0.42 |
| Butyric acid | 1.31 |
| Isobutyric acid | 0.15 |
| Valeric acid | 0.1 |
| Isovaleric acid | 0 |
| Caproic acid | 0.75 |
| Ammonium | 0.27 |
| Total COD | 13.2 |
| Carbon Source | Conditions | Maximum Biomass Production (g/L) | Maximum PHA Accumulation (g/L) | Maximum PHA Yield on Biomass (%) | Maximum Biomass Yield on Substrate (%) | Maximum PHA Yield on Substrate (%) |
|---|---|---|---|---|---|---|
| Synthetic VFAs | (NH4)2SO4, pH control, C/N 6, 1 vvm | 2.80 (36 h) | 1.49 (36 h) | 55.74 (30 h) | 82.08 (12 h) | 17.54 (24 h) |
| (NH4)2SO4, no pH control, C/N 6, 1 vvm | 1.98 (36 h) | 1.29 (36 h) | 65.31 (36 h) | 58.20 (24 h) | 31.81 (30 h) | |
| (NH4)2SO4, pH control, C/N 12, 1 vvm | 2.25 (24 h) | 1.15 (24 h) | 50.94 (24 h) | 45.53 (24 h) | 10.30 (24 h) | |
| (NH4)2SO4, pH control, C/N 6, 0.25 vvm | 2.32 (60 h) | 1.20 (54 h) | 60.04 (30 h) | 72.05 (24 h) | 34.32 (30 h) | |
| Urea, pH control, C/N 6, 1 vvm | 2.80 (36 h) | 1.51 (30 h) | 56.50 (30 h) | 73.07 (24 h) | 18.12 (24 h) | |
| Urea, no pH control, C/N 6, 1 vvm | 2.16 (36 h) | 1.60 (36 h) | 75.34 (30 h) | 62.51 (24 h) | 24.76 (24 h) | |
| Real VFAs | pH control, C/N 12, 1 vvm | 2.26 (36 h) | 1.02 (36 h) | 50.60 (24 h) | 72.12 (30 h) | 20.42 (24 h) |
| Glucose | (NH4)2SO4, pH control, C/N 6, 1 vvm | 2.45 (36 h) | 1.66 (30 h) | 67.87 (30 h) | 53.59 (24 h) | 16.24 (24 h) |
| Mole% | Total | ||
|---|---|---|---|
| Sample | HB | HV | Crystallinity * |
| VFA/NH4SO4 | 93.8 | 6.2 | 62.2 |
| VFA/urea | 91.7 | 8.3 | 58.3 |
| Glucose batch | 95.9 | 4.1 | 66.5 |
| Real VFA stream | 93.5 | 6.5 | 61.4 |
| Samples | Tonset (°C) | Tmax (°C) | Total Mass Loss (%) | Tm (°C) | Melting Enthalpy (J/g) | Degree of Crystallinity (%) |
|---|---|---|---|---|---|---|
| Glucose, (NH4)2SO4 | 218.4 | 316.8 | 98.5 | 176.1 | 66.4 | 45.5 |
| Synthetic VFA, (NH4)2SO4 | 212.5 | 317.7 | 97.5 | 154.7 | 37.2 | 25.5 |
| Synthetic VFA, Urea | 213.9 | 315.1 | 96.3 | 156.3 | 31.7 | 21.7 |
| Real VFA stream | 201.1 | 305.7 | 81.2 | 151.9 | 15.8 | 10.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, D.H.; Mahboubi, A.; Root, A.; Heinmaa, I.; Taherzadeh, M.J.; Åkesson, D. Thorough Investigation of the Effects of Cultivation Factors on Polyhydroalkanoates (PHAs) Production by Cupriavidus necator from Food Waste-Derived Volatile Fatty Acids. Fermentation 2022, 8, 605. https://doi.org/10.3390/fermentation8110605
Vu DH, Mahboubi A, Root A, Heinmaa I, Taherzadeh MJ, Åkesson D. Thorough Investigation of the Effects of Cultivation Factors on Polyhydroalkanoates (PHAs) Production by Cupriavidus necator from Food Waste-Derived Volatile Fatty Acids. Fermentation. 2022; 8(11):605. https://doi.org/10.3390/fermentation8110605
Chicago/Turabian StyleVu, Danh H., Amir Mahboubi, Andrew Root, Ivo Heinmaa, Mohammad J. Taherzadeh, and Dan Åkesson. 2022. "Thorough Investigation of the Effects of Cultivation Factors on Polyhydroalkanoates (PHAs) Production by Cupriavidus necator from Food Waste-Derived Volatile Fatty Acids" Fermentation 8, no. 11: 605. https://doi.org/10.3390/fermentation8110605