3.1. Acidity, Microbiological and Chromaticity Characteristics of Wholemeal Wheat Flour Samples
The pH, total titratable acidity (TTA), colour and microbiological characteristics of the wheat grain wholemeal are shown in
Table 1. Significant differences between the control group (non-fermented) and fermented dough samples were found when comparing the acidity, and the lowest pH was achieved with the wholemeal ‘Gaja’ and DS8535—2 (purple wheat) samples (on average, 6.00). However, higher TTA values were found in ‘Gaja’ and DS8888-3-6 (waxy wheat) samples (on average, 1.22° N), in comparison with DS8548—7 (blue wheat) and DS8535—2 (purple wheat) (on average, 1.01° N). When comparing the pH of non-treated and fermented samples, in all the fermented groups, the lowest pH was reached in samples fermented with
Pediococcus acidilactici strain. In addition, most of the latter samples (fermented ones) showed the highest TTA, as expected. The test between the subjects showed that the LAB strain used for fermentation was a significant factor in TTA (
p ≤ 0.0001). Moreover, a moderate positive correlation was established (r = 0.552,
p ≤ 0.001) between the TTA and type of LAB. The concentration of organic acids in fermented wholemeal flours depend on numerous factors, including LAB strain used, processing parameters and flour composition. [
30]. However, controlling excessive acidity is one of the major challenges in fermented wheat, especially when they are used as a sourdough for breadmaking. Previous studies reported the influence of the interaction of processing factors on fermented cereal acidity parameters, microorganism growth, volatile compounds and amino acids formation [
30]. However, the changes in waxy and coloured wheat grain during the fermentation process were not described. Thus, this study demonstrated that the cereal variety is not a significant factor in the acidity parameters (pH and TTA) of the fermented wholemeal wheat flours when using
Pediococcus acidilactici,
Liquorilactobacillus uvarum and
Lactiplantibacillus plantarum. It was also possible to conclude that all these LAB strains are suitable for acidic fermentation of the new varieties of wholemeal wheat grains under scrutiny in this research study.
When analysing the colour characteristics of non-fermented and fermented wholemeal wheat samples, with all the tested LAB strains, the fermented ‘Gaja’ samples showed higher L* (lightness) coordinates in comparison with non-fermented ones. However, opposite tendencies of the waxy, blue and purple wholemeal wheat cereals were found, as in the latter samples, L* decreased after fermentation (except DS8888-3-6 fermented with
Lactiplantibacillus plantarum). Cereal variety proved to be a significant factor in the L* coordinates of the samples (
p = 0.023). Different trends were established when comparing a* (redness) coordinates. After fermentation, a* coordinates showed tendencies to increase in all fermented ‘Gaja’ samples, in wholemeal waxy wheat fermented with
Pediococcus acidilactici, wholemeal waxy wheat cereal fermented with
Liquorilactobacillus uvarum and in wholemeal purple wheat fermented with
Pediococcus acidilactici. However, fermentation decreased a* coordinates in all the fermented wholemeal blue wheat cereal samples. Comparing the b* (yellowness) coordinates, they were lower in both fermented wholemeal coloured wheat in comparison with non-fermented ones. However, in traditional wheat ‘Gaja’, fermentation with
Liquorilactobacillus uvarum and
Lactiplantibacillus plantarum increased b* coordinates by 76.5 and 108.7%, respectively. Opposite trends were found in wholemeal waxy wheat samples, as fermentation with
Pediococcus acidilactici and
Liquorilactobacillus uvarum decreased b* values by 27.1 and 20.2%, respectively, in comparison with non-fermented ones. Variation in colour components in the cereal grain depends on genetic factors, growing conditions, as well as on technological processes [
12]. Yellow C-glycosides of flavones are mainly present in the outer layer of cereal grains. However, carotenoids are located in the endosperm [
12]. In addition, there is a great diversity of anthocyanin composition among various cereals [
31], and the stability of these compounds can be influenced by substrate pH, temperature, the presence of ascorbic acid, oxygen and enzymes, among other factors [
32]. This study showed that the LAB strain used for fermentation, cereal variety and the interaction between these factors had a significant impact on a* coordinates of wholemeal cereal samples (
p = 0.021,
p ≤ 0.023
p ≤ 0.001, respectively), and in most of the cases, fermented samples showed lower redness and yellowness.
Comparing LAB viable counts in fermented wholemeal wheat flours, the purple wheat samples fermented with
Pediococcus acidilactici and
Liquorilactobacillus uvarum showed the highest numbers of viable LAB (on average, 8.83 log
10 CFU/g). As a matter of fact, technological microorganisms play a major role in developing the characteristics of fermented cereal flours [
33]. A moderate positive correlation was established between the LAB viable counts and TTA (r = 0.552,
p ≤ 0.001). In addition, the type of LAB strain used for fermentation was a significant factor on samples TTA (
p ≤ 0.0001). Fermented cereal dough is a stressful ecosystem that is characterized by a low pH, high carbohydrate concentrations, oxygen limitation and viable cell counts of LAB ≥ 10
8 CFU/g [
34]. In this study, all the fermented wholemeal wheat flour samples showed LAB viable counts higher than 8.00 log
10 CFU/g, and the type of LAB strain used for fermentation was a significant factor in LAB viable counts of wholemeal fermented flours (
p ≤ 0.0001). Finally, despite the colour intensity of the colored wheats being reduced during fermentation, all the tested wholemeal wheat flour samples showed desirable low pH values and a high number of viable LAB after fermentation.
3.2. Amino Acids (AA), γ-Aminobutyric Acids (GABA) and Biogenic Amines (BA) of Wholemeal Wheat Flour Samples
Essential amino acid concentrations of the wholemeal wheat grain samples are shown in
Table 2. When comparing non-fermented samples, no significant differences were found in threonine, methionine, phenylalanine and histidine concentrations. However, in comparison with traditional and waxy wheat samples, higher valine concentration in both colored wholemeal wheat cereals was established (on average, by 14.6% higher). In contrast to these findings, in comparison with both colored wheat, traditional (‘Gaja’) and waxy wheat samples showed higher concentrations of leucine/isoleucine. Simultaneously, the highest concentration of lysine was found in ‘Gaja’ samples (0.360 µmol/g). In most of the cases, fermentation increased essential amino acid concentration in wholemeal wheat flour samples, but methionine in DS8888-3-6 La.p and in DS8548-7 La.p, and histidine in ‘Gaja’Li.u, DS8888-3-6 La.p. and DS8548-7 La.p. The LAB strain used for fermentation proved to constitute a significant factor on all the essential amino acid concentrations in wholemeal wheat grains (
p ≤ 0.0001). On the other hand, wheat variety was a significant factor on threonine, methionine, valine, lysine and histidine concentrations (
p = 0.002,
p ≤ 0.0001,
p = 0.002,
p = 0.034,
p ≤ 0.0001, respectively), and interaction of these factors was showed to be significant on methionine, valine and histidine contents (
p ≤ 0.0001,
p ≤ 0.0001, and
p ≤ 0.01, respectively).
Nonessential amino acid and GABA concentrations of the wholemeal wheat grain samples are shown in
Table 3. Regarding the nonessential amino acid concentrations in non-fermented samples, ‘Gaja’ and waxy wholemeal wheat flours showed higher arginine content, in comparison with both wholemeal coloured wheat, and wheat variety has shown to be a significant factor on arginine content (
p = 0.21).
The highest glutamine concentration was found in wholemeal blue wheat (2.52 µmol/g), and the wheat variety was a significant factor in glutamine content (p ≤ 0.0001). The highest glutamic acid and glycine concentration was obtained in the ‘Gaja’ samples. The highest serine content was found in wholemeal waxy wheat, and the highest alanine content was established in wholemeal blue wheat. Wheat variety was a significant factor in the content of most of these amino acids, but alanine (p = 0.015, p = 0.028 and p = 0.009, respectively). The aspartic acid concentration in ‘Gaja’ and purple wheat samples was, on average, 15.5% lower in comparison with wholemeal waxy and blue wheat grain samples. The lowest proline concentration was found in the ‘Gaja’ samples (0.285 µmol/g). In all the cases, tyrosine content in non-fermented samples was lower than 0.020 µmol/g. When comparing fermented samples, different tendencies were found, and the type of LAB strain used for fermentation represented a significant factor in arginine, glutamine, aspartic acid, glycine, alanine, proline and tyrosine content (p ≤ 0.0001, p ≤ 0.0001, p ≤ 0.0001, p = 0.004, p ≤ 0.0001, p ≤ 0.0001, p ≤ 0.0001, respectively). The interaction between LAB strain used for fermentation and wheat variety was significant with regards to the arginine, glutamine, glutamic acid, alanine, proline and tyrosine concentrations (p ≤ 0.0001, p ≤ 0.0001, p = 0.027, p = 0.005, p = 0.003, p = 0.039, respectively).
Despite wheat grains being a good source of protein, the amounts of essential amino acids (especially lysine) are low [
35]. Finally, the quality of the protein must be taken into consideration because the cereal-based diet tends to be deficient in some essential amino acids. Having in mind that the amino acid profile (especially essential amino acids) shows the quality of protein, the use of fermentation in this study showed prospective results, as most of the essential amino acid concentrations in wholemeal wheat cereal samples increased. Cereal grain proteins supply 47% of total global protein intake [
36], and cereal-based proteins are a predominant protein source in developing countries [
37]. For this reason, finding technological solutions to increase the biological value of wheat cereal varieties becomes very important. Protein content in wheat grains can vary due to genetic differences, agronomic factors, cultivation conditions, etc., but essential amino acids are the most important parameter from nutritional and physiological points of view [
36,
38,
39]. Better knowledge of the amino acid profile of different wheat varieties is beneficial not only to wheat farmers but also to food and feed manufacturers [
40]. The recommended daily intake of total essential amino acids is 83.5 mg/kg of body weight [
41]. In addition to lysine, threonine, which is present at low concentrations in cereals, is very important from a nutritional standpoint [
42]. Recommended daily intake for threonine is about 19 mg/kg [
43]. Recommended daily intake for histidine is 8 and 12 mg/kg of body weight/day in adults [
44,
45,
46]. Phenylalanine is essential for the growth and development of children [
47]. Leucine stimulates muscle protein synthesis [
44,
48]. Recommended daily intake for leucine is 39 mg/kg body weight per day [
41].
It was reported that in wheat cereal grains, the predominant nonessential amino acid is glutamic, which concentration can range from 24.79 to 37.05 g/100 g protein [
49]. Glutamic acid plays a major role in amino acid metabolism in the body and shows neurotransmitter function [
47]. The mean daily intake of glutamic acid is 15 g/day [
44]. Moreover, usually, wheat grains show low content of cysteine, methionine and histidine [
50].
The results of this study showed that the quality of wholemeal wheat grain proteins can be significantly improved trough the modification of the amino acid profile by using fermentation biotechnology with selected LAB strains. However, it should be pointed out that for a feed industry, these results can be of great interest because one can obtain products with higher biological added value for feed purposes. However, for the food industry, fermented wholemeal wheat grain should be further tested (e.g., in bread production), and the quality, as well as the food safety parameters of the end-products, must be taken into consideration because, during the thermal treatment, various (desirable and not) changes may occur.
When comparing GABA concentration in non-fermented samples, the highest content was found in wholemeal ‘Gaja’ and waxy wheat samples (on average, 2.47 µmol/g). Conversely, fermentation increased GABA concentration in all wholemeal cereal doughs. In addition, the LAB strain used for fermentation, the wheat variety and the interaction of these factors were shown to be significant on GABA concentration in wholemeal wheat grain samples (p ≤ 0.0001, p = 0.002 and p = 0.004, respectively).
GABA is a non-protein amino acid compound produced during the decarboxylation of L-glutamic acid [
51]. GABA shows many desirable effects in the body, including neurotransmission and regulation of brain metabolism [
52].
Due to the activity of LAB (decarboxylation, deamination, transamination, etc.), the production of several functional compounds in cereal grains may occur [
53,
54,
55,
56,
57], including GABA [
58,
59]. It was reported that during the fermentation with LAB, GABA could be formed [
60,
61,
62], and, as a matter of fact, fermented food is a good source of dietary GABA [
53]. The selection of cereal varieties, based on their nutritional and functional potential, may be very useful in improving the features of food and feed products. Finally, our study showed that fermentation with selected LAB increases GABA concentration in all the tested wholemeal wheat cereal samples and can be a very prospective technology to increase the functional value of cereals.
Biogenic amine concentration in wholemeal wheat cereal samples is shown in
Table 4. Phenylethylamine was found in all the wholemeal cereal samples (fermented and non-fermented), and a higher concentration of this compound was disclosed in non-fermented samples of the wholemeal ‘Gaja’, waxy and blue wheat flour (on average, by 14.2, 24.1 and 27.2%, respectively, higher in comparison with fermented ones). However, opposite trends were found in wholemeal purple wheat cereal samples, and the lowest phenylethylamine concentration was found in samples fermented with
Lactiplantibacillus plantarum (30.9 mg/kg). Putrescine was observed in non-fermented ‘Gaja’ samples, in waxy wheat cereal samples fermented with
Liquorilactobacillus uvarum, and in wholemeal waxy, blue and purple wheat cereals fermented with
Lactiplantibacillus plantarum. Waxy wheat samples fermented with
Lactiplantibacillus plantarum showed 8.3 and 2.3 times higher concentrations of putrescine, in comparison with wholemeal blue and purple wheat, respectively, fermented in the same conditions. Cadaverine was formed in all samples fermented with
Liquorilactobacillus uvarum and
Lactiplantibacillus plantarum strains. When comparing cadaverine concentration between different cereal groups fermented with the same LAB strain, the highest concentration was found in purple wheat samples fermented with
Liquorilactobacillus uvarum (122.6 mg/kg). Furthermore, the cereal group consisting of wholemeal ‘Gaja’, waxy and purple wheat samples fermented with
Lactiplantibacillus plantarum showed, on average, a concentration as high as 120.0 mg/kg cadaverine. Histamine was just formed in wholemeal waxy, blue and purple wheat cereal samples fermented with
Lactiplantibacillus plantarum, and the highest concentration of histamine was reached in wholemeal purple wheat samples (66.3 mg/kg).
Tyramine (35.6 mg/kg) was formed in wholemeal purple wheat cereal samples fermented with Lactiplantibacillus plantarum, and it also displayed the highest spermidine concentration (51.8 mg/kg). When comparing spermidine in all wholemeal cereal groups, non-fermented wholemeal ‘Gaja’, waxy and blue flours contained spermidine concentrations above 20.0 mg/kg. Nevertheless, spermidine was not found in non-fermented whole purple wheat samples. In opposite to these trends, spermidine was found in all the fermented whole purple wheat samples, and its content ranged from 24.5 to 51.8 mg/kg (in fermented samples with Pediococcus acidilactici and Lactiplantibacillus plantarum, respectively). Spermine was just found in wholemeal purple wheat flour fermented with Lactiplantibacillus plantarum (20.2 mg/kg). Multivariate analyses of between-subjects effects indicated that the LAB strain used for fermentation, the wheat variety and their interaction were significant factors in the concentration of all biogenic amines (p ≤ 0.0001).
Increased concentrations of cadaverine, putrescine and spermidine in products are associated with microbial activity [
63]. There are a few mechanisms of action for the accumulation of biogenic amines in food, which are explained by their possible synthesis via the decarboxylation of amino acids, as well as by transamination of aldehydes or ketones [
64,
65]. There are published works highlighting the negative effects of tyramine and phenylethylamine on human [
66] and animal health [
67]. In addition to the negative effect on health, putrescine and cadaverine are characterised as specifically smelly molecules [
68], whose presence in food or feed may reduce their acceptability. The total amount of biogenic amines in food is very important because they can increase toxicity [
66,
69]. Arginine can be metabolised via bacterial action to putrescine; lysine can be converted to cadaverine, histidine to histamine, tyrosine to tryptophan and phenylalanine to tyramine, tryptamine and β-phenylethylamine, respectively [
70]. Histamine is the most toxic biogenic amine [
71,
72,
73]. Tyramine is related to migraines, and cadaverine and putrescine can increase the toxic effect of other biogenic amines [
74]. The Food and Drug Administration (FDA) suggested that the maximum permitted histamine in food should be limited to 500mg/kg, and a level of >1000 mg/kg should be considered, for safety for consumption purposes, as unfit for human consumption [
75,
76]. The European Union (EU) regulation suggests values between 100 and 200 mg of histamine per kilogram of food for the
Scomberesocidae and
Scombridae fish families [
77], as well as the highest concentration of 300 mg/kg for the total biogenic amine content in fish and derivatives.
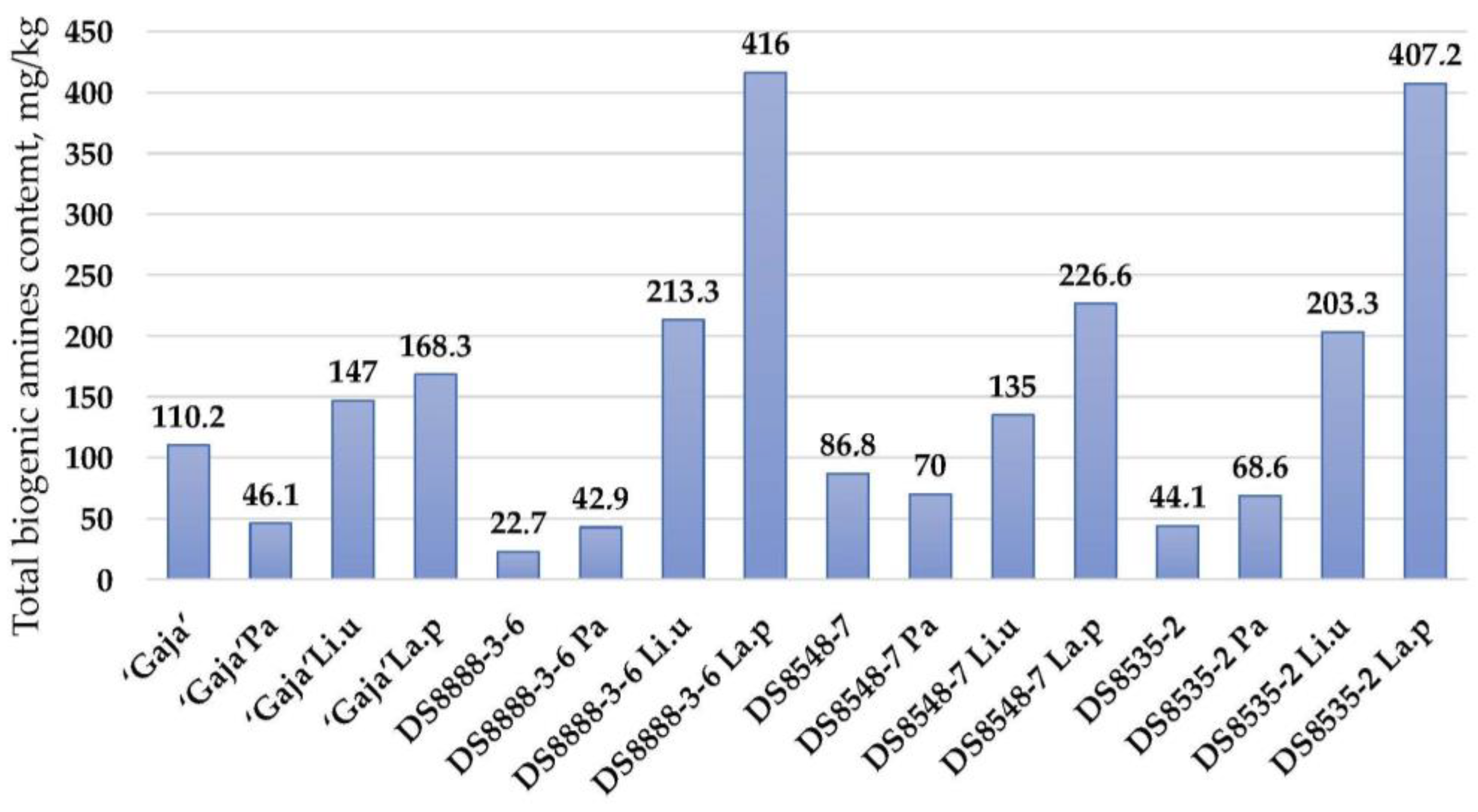
According to the results obtained in this study, the formation of biogenic amines in whole wheat cereal products should be taken into consideration, as the total levels of biogenic amine ranged from 22.7 (in non-fermented DS8888-3-6 samples) to 416 mg/kg (in samples DS8548-7 fermented with
Lactiplantibacillus plantarum) (
Figure 2). Moderate positive correlations were found between cadaverine concentration, TTA and viable LAB viable counts (r = 0.308,
p = 0.033 and r = 0.581,
p = 0.0001, respectively). However, a moderate positive correlation between histamine concentration and LAB viable counts was revealed (r = 0.306,
p = 0.034).
3.3. Macro- and Microelements of Wholemeal Wheat Flour Samples
Macroelement concentrations in wholemeal wheat cereal samples are shown in
Table 5. Significant differences were found in the Na content when comparing the macroelement concentrations in non-fermented and fermented groups, which, in all the cases, was lower in non-fermented samples. These differences can be explained by the addition of the LAB suspension, which contained a physiological (NaCl) solution.
When comparing the varieties of wholemeal wheat cereals under scrutiny, the highest Mg and K concentrations were obtained in wholemeal ‘Gaja’ wheat. However, the highest Ca content was found in wholemeal waxy wheat samples. Multivariate test of between-subject effects showed that the LAB strain used for fermentation and the interaction with the wheat variety had a significant impact on the Na concentration in wholemeal cereals (p ≤ 0.0001) and showed as well that the wheat variety was a significant factor on the concentration of all the analysed macroelements (p ≤ 0.0001).
Essential microelement concentrations of the wholemeal wheat cereal samples (except Co, Ni and Se, whose concentrations were lower than 0.010, 0.5 and 0.20 mg/kg, respectively) are given in
Table 6. A concentration higher than 0.010 mg/100 g Cr was only detected in wholemeal waxy wheat samples (on average, 0.154 mg/100 g), and fermentation was not a significant factor towards the Cr content. Wholemeal blue and purple wheat samples showed the highest Mn concentration (on average, 16.2 mg/kg), and, comparatively, in wholemeal ‘Gaja’ and waxy wheats, the Mn concentration, on average, was 21.0 and 53.5% lower, respectively. The highest Fe content was found in wholemeal ‘Gaja’ (on average, 24.8 mg/kg), and s waxy, blue and purple wheats showed, on average, 35.1% lower Fe concentrations. Moreover, wholemeal ‘Gaja’ showed the highest Cu content (on average, 2.19 mg/kg), and wholemeal waxy, blue and purple wheat samples showed, on average, 47.9, 55.6 and 63.2% lower Cu concentrations, respectively. The highest concentration of Zn was found in wholemeal purple wheats (8.31 mg/kg), and wholemeal ‘Gaja’, waxy and blue wheats showed, on average, 21.1, 33.7 and 20.0% lower Zn contents, respectively. A multivariate test of between-subject effects showed that the LAB strain used for fermentation was a significant factor in Cr concentration in wholemeal cereals (
p = 0.007). On the other hand, wheat variety was a significant factor in Cr, Mn and Cu contents (
p ≤ 0.0001), and LAB strain and wheat variety interaction were significant factors in Cr concentration in wholemeal wheat samples (
p ≤ 0.0001).
Non-essential microelement concentrations of the wholemeal wheat cereal samples (except V, Mo, Ag, Sb, Cs, Ti, Al and Li, whose concentrations were lower than 2, 0.5, 2, 0.5, 2, 2, 5 and 0.050 mg/kg, respectively) are shown in
Table 7. Wholemeal ‘Gaja’ samples showed higher concentrations of As and Rb in comparison with wholemeal waxy, blue and purple wheat flours, in which these microelements were absent. The highest concentration of Sr was found in wholemeal blue wheat samples (2.58 mg/kg), and the wholemeal ‘Gaja’ samples showed the highest Cd content (0.54 mg/kg). The highest Ba concentration was found in wholemeal blue wheat samples, and the highest Pb content was disclosed in purple wheat wholemeal.
A multivariate test of between-subject effects revealed that the wheat variety was a significant factor on all the analysed non-essential microelement contents (p ≤ 0.0001) in wholemeal cereals. This finding can lead to the conclusion that different cereal varieties present different capacities to accumulate these elements from the environment.
Cereal grains are an important source of macro- and microelements [
78]. Macro- and microelements are essential compounds for human and animal nutrition [
79]. The mineral and trace element contents of many raw plant-based materials are known to be related to the cultivar and variety of plant, soil and weather conditions, fertilisers used, etc. [
80,
81]. The levels of some elements can vary in a large range [
78]. From the nutritional standpoint, major attention has been given to the elements Fe, Ca, Cu and Zn [
82].
However, other trace elements are also very important, and micronutrient malnutrition afflicts billions of people [
83]. According to World Health Organization (WHO), 46% of children (from 5 to 14 years-old) have Fe deficiency anaemia, 48% of pregnant women are Fe deficient [
84], and, on average, 200 million people lack essential trace elements [
85]. The food industry tries to solve this problem by the addition of nutrients to food, but they do not ensure nutrient absorption from the end product [
86]. If nutrient-rich wheat cereal grain can be bred, the problems mentioned above might be solved. It was reported that coloured wheat grains contain numerous nutrients which are beneficial to human health [
87,
88,
89], and black wheat showed higher contents of iron, zinc, manganese, copper, selenium, magnesium, potassium and phosphorus when compared to traditional ones [
90]. Likewise, organic chromium content in black wheat was, on average, four times higher in comparison with traditional ones [
90]. Correspondingly, it was reported that Se concentration in purple wheat is higher than in control by, on average, 173.6% [
83].
Finally, the determination of the nutrient content in cereals, such as essential elements, is necessary to perceive their dietary intake. Calcium is an element in bone tissues. Potassium and Na play an important role in maintaining osmotic pressure and in the transmission of nervous impulses, and Mg is a cofactor in more than 300 enzymatic reactions. Trace elements (Fe, Cu, Co, Zn, Mn, Mo) are also needed for the proper functioning of the organism but are required in smaller quantities [
91,
92,
93,
94,
95]. These trace elements are involved in numerous biological processes: Fe is involved in oxygen transport, Co is an element forming part of cobalamin or vitamin B12, and Mo, Mn, Zn or Cu are components in many enzymes [
96,
97,
98]. This study, considering the importance of cereals in the diet, demonstrates the importance of creating databases of new wheat varieties, which can be a good source of micro- and macroelements in human and animal diets.
3.4. Fatty Acid Composition of Wholemeal Wheat Flour Samples
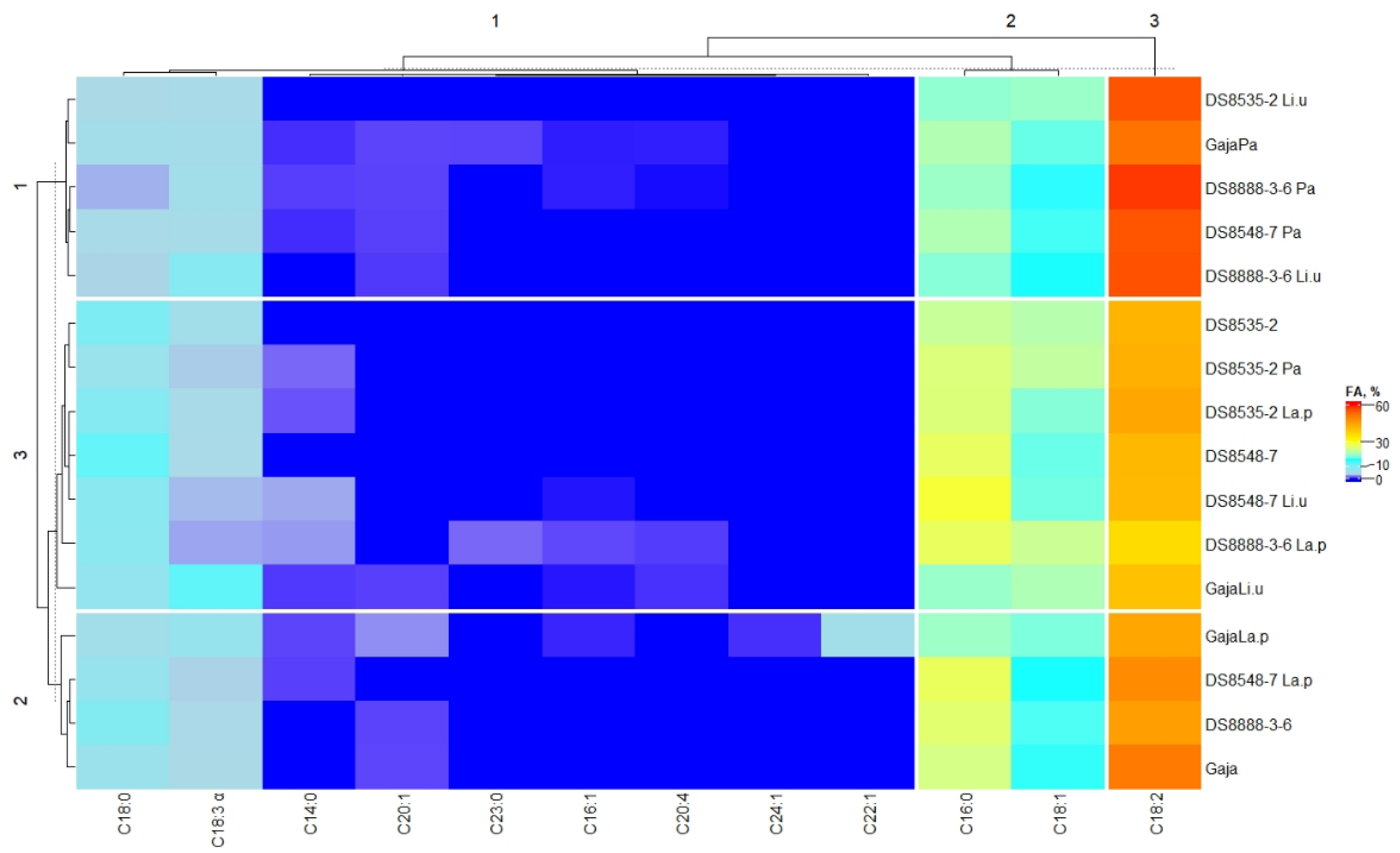
Fatty acid composition (expressed as a percentage of the total fatty acid content) of the wholemeal wheat cereal samples is given in
Table 8. Moreover, the detailed fatty acid profile is given in
Supplementary Table S1. A detailed fatty acid profile of the wheat cereal wholemeal samples is in the heatmap of
Figure 3.
The highest saturated fatty acids content was found in wholemeal blue wheat flour (38.14% from the total fatty acid content) when comparing the non-fermented samples. Nonetheless, in most of the cases, fermentation reduced the saturated fatty acid content of wholemeal wheat samples—except wholemeal waxy and purple wheat fermented with Lactiplantibacillus plantarum and wholemeal blue wheat fermented with Liquorilactobacillus uvarum. A multivariate test of between-subjects effects showed that the LAB strain used for fermentation, wheat variety and the interaction of these factors were significant in the saturated fatty acid content of wholemeal wheat cereals (p ≤ 0.0001).
Different tendencies of the monounsaturated fatty content were obtained. In addition, when comparing non-fermented samples, the highest content was displayed in wholemeal purple wheat samples (21.33% from the total fatty acids content). However, after fermentation with Lactiplantibacillus plantarum and Liquorilactobacillus uvarum, the monounsaturated fatty content in wholemeal purple wheat was reduced. Conversely, in samples fermented with Pediococcus acidilactici, an increase in monounsaturated fatty acid content was detected. Furthermore, fermentation with any of the tested LAB strains increased monounsaturated fatty acid content in wholemeal ‘Gaja’ wheat cereal. Moreover, it also increased monounsaturated fatty acid content in wholemeal waxy wheat cereal samples fermented with Lactiplantibacillus plantarum, in wholemeal blue wheat cereal samples fermented with Liquorilactobacillus uvarum fermented, and, finally, in wholemeal purple wheat cereal samples fermented with Pediococcus acidilactici. A multivariate test of between-subjects effects showed that the type of LAB strain used for fermentation and its interaction with the wholemeal wheat variety had a significant effect on the monounsaturated fatty acid content of wholemeal wheat flours (p = 0.038).
When analysing polyunsaturated fatty acids in non-fermented samples, the highest content was found in wholemeal ‘Gaja’ wheat cereal. In turn, in fermented samples, different tendencies were established, and, in most of the cases, fermentation increased the polyunsaturated fatty acid content of wholemeal wheat cereal samples, viz.: in wholemeal ‘Gaja’, waxy and blue wheat cereal samples fermented with Pediococcus acidilactici; in wholemeal waxy and purple wheat cereal samples fermented with Liquorilactobacillus uvarum; and in wholemeal blue and purple wheat cereal samples fermented with Lactiplantibacillus plantarum fermented. However, analysed factors were not significant in the polyunsaturated fatty acid content of wholemeal cereal samples.
Comparing the omega-3 (ω-3), omega 6 (ω-6) and omega 9 (ω-9) content, fermentation reduced ω-3 content in wholemeal waxy, blue and purple wheat cereal samples fermented with Lactiplantibacillus plantarum, as well as in wholemeal blue and purple wheat cereal samples fermented with Liquorilactobacillus uvarum, and in wholemeal purple wheat cereal samples fermented with Pediococcus acidilactici. A lower content of ω-6 after fermentation was found in wholemeal ‘Gaja’ and blue wheat cereal samples fermented with Liquorilactobacillus uvarum, and in wholemeal ‘Gaja’ and waxy wheat cereal samples fermented with Lactiplantibacillus plantarum. Moreover, fermentation with Lactiplantibacillus plantarum reduced ω-9 content in both wholemeal colored wheat samples; fermentation with Pediococcus acidilactici and Liquorilactobacillus uvarum reduced ω-9 content in wholemeal waxy wheat; and finally, fermentation with Pediococcus acidilactici and Liquorilactobacillus uvarum was a significant factor on reducing ω-9 content in wholemeal blue and purple wheat samples, respectively. Multivariate test of between-subjects effects showed that the LAB strain used for fermentation and its interaction with wholemeal wheat variety interaction was significant on ω-6 content of wholemeal wheat cereal samples (p ≤ 0.050). However, analysed factors and their interaction were not significant on ω-3 and ω-9 concentrations of wholemeal cereal samples.
When analysing the content of individual fatty acids in wholemeal wheat samples, the results unveiled that the dominant fatty acids in all wholemeal wheat cereal samples were linoleic acid (C18:2), palmitic acid (C16:0), octadec-9-enoic acid (C18:1 cis, trans), stearic acid (C18:0) and α-linolenic acid (C18:3 α) (
Figure 3). Moreover, fermented samples revealed a wider variety of individual fatty acids. In most of the fermented samples, tetradecanoic acid (C14:0) was formed, with the highest content observed in DS8548-7 Li.u wholemeal samples (2.24% of the total fatty acid content). Moreover, six fermented samples showed, in their fatty acid profile, the presence of palmitoleic acid (C16:1), and the highest concentration of C16:1 was found in DS8888-3-6 La.p cereal wholemeal (0.682% from the total fatty acid content). Cis-5,8,11,14-eicosatetraenoic acid (C20:4) accounted for just 4 out of 16 analysed wholemeal samples, and its content ranged from 0.051 to 0.527% of the total fatty acid content (in DS8888-3-6 Pa and DS8888-3-6 La.p, respectively). Cis-13-docosenoic acid (C22:1) (4.88% from the total fat content) and cis-15-tetracosenoic acid (C24:1) (0.367% from the total fat content) were formed just in ‘Gaja’La.p samples. Moreover, tricosanoic acid (C23:0) was just found in ‘Gaja’Pa and DS8888-3-6 La.p wholemeal cereal samples (0.628 and 1.17% from the total fatty acid content, respectively). Multivariate test of between-subjects effects showed that the type of LAB strain used for fermentation was a significant factor on the concentrations of C14:0, C16:1, C18:0, C18:2, C20:4, C22:1, C23:0 and C24:1 in wholemeal cereals (
p ≤ 0.0001,
p ≤ 0.0001,
p = 0.034,
p = 0.023,
p ≤ 0.0001,
p ≤ 0.0001,
p ≤ 0.0001,
p ≤ 0.0001, respectively). Moreover, wholemeal wheat variety was a significant factor on the content of C14:0, C16:1, C18:3 α, C20:1, C20:4, C22:1, C23:0 and C24:1 (
p ≤ 0.0001,
p ≤ 0.0001,
p = 0.002,
p ≤ 0.0001,
p ≤ 0.0001,
p ≤ 0.0001,
p ≤ 0.0001 and
p ≤ 0.0001, respectively). However, the interaction of LAB strain and wheat variety was significant on the concentrations of C14:0, C16:1, C18:0, C18:2, C18:3 α, C20:1, C20:4, C22:1, C23:0 and C24:1 in wholemeal wheat samples (
p ≤ 0.0001,
p ≤ 0.0001,
p = 0.004,
p ≤ 0.0001,
p ≤ 0.001,
p ≤ 0.0001,
p ≤ 0.0001,
p ≤ 0.0001, respectively).
It was reported that cereal grains are rich in unsaturated fatty acids as well as are a good source of essential polyunsaturated fatty acids (namely, linoleic and linolenic acids) [
99]. Wheat lipids also contain saturated fatty acids [palmitic (20.5%) and stearic acids (1.5%)]. The fatty acids composition of wheat grain makes from a nutritional point of view the cereals balanced foodstuffs [
99].
Dominant unsaturated fatty acids in wheat are C18:2, C18:1, C18:3 and C16:1 (which includes two essential ones to know: linoleic and linolenic acids) [
99]. Essential fatty acids are involved in many metabolic processes, including cholesterol metabolism [
100,
101,
102]. The fatty acid profile of wheat is dependent on many factors, including genetics [
103], environmental conditions and agronomic practices [
100,
103,
104]. It was reported that durum and hard red wheat have a higher lipid content and distinct fatty acid profile in comparison with soft white wheat [
99]. Moreover, cold weather can increase the total lipid concentration and the unsaturated fatty acid content in wheat cereal grains [
105]. Bottari et al. reported more than 60 fatty acids in wheat cereal, with even numbers of carbon atoms from C12 to C30 as well as C15 and C17. The major fatty acids were saturated and unsaturated C16 and C18 and, particularly, C16:0, C18:1 and C18:2, which together represented around 90% of the total [
106]. However, the main fatty acids in wheat cereals are C16:0, C18:0, C18:1, C18:2 and C18:3, and other fatty acid contents represent, on average, 1–2%. The differences in wheat fatty acid profile reported in different studies are explained by the different genetic, climatic and agronomical conditions, as well as the interaction between genotype × year × treatment as the main factor towards the variability of the fatty acid concentration observed in durum wheat samples [
104]. Armanino et al. reported that the fatty acid profile of durum wheat is influenced by the cultivar [
103]. In addition, variation in saturated and unsaturated fatty acids within the same wheat cereal grain variety can be obtained, and both biotic and abiotic stresses are related to these differences [
105,
107]. Finally, the fatty acid composition and level of unsaturation also vary according to the type of cereal, for example, when comparing maize and rye flours [
108,
109,
110,
111,
112]. Regarding the fatty acid content, this study showed that in addition to the wheat variety, the process of fermentation is a significant factor on most of the fatty acid content in wholemeal wheat cereal grain. Finally, the changes of fatty acid profile during the biotechnological processes should also be taken into consideration to achieve the best technological and nutritional characteristics of the cereal-based end product.

 ,
, 




{kind=link}
{kind=link}
{kind=link}