
Molecular Identification and Biochemical Characterization of Novel Marine Yeast Strains with Potential Application in Industrial Biotechnology

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Environmental Sampling
2.2. Screening and Isolation
2.3. Molecular Identification
2.4. Phylogenetic Analysis
2.5. Biochemical Characterization
2.6. Cytotoxicity of Yeast Strains on HEK293 Cell Viability
2.7. Statistical Analysis
3. Results and Discussions
3.1. Screening and Isolation of Marine Yeast
3.2. DNA Identification
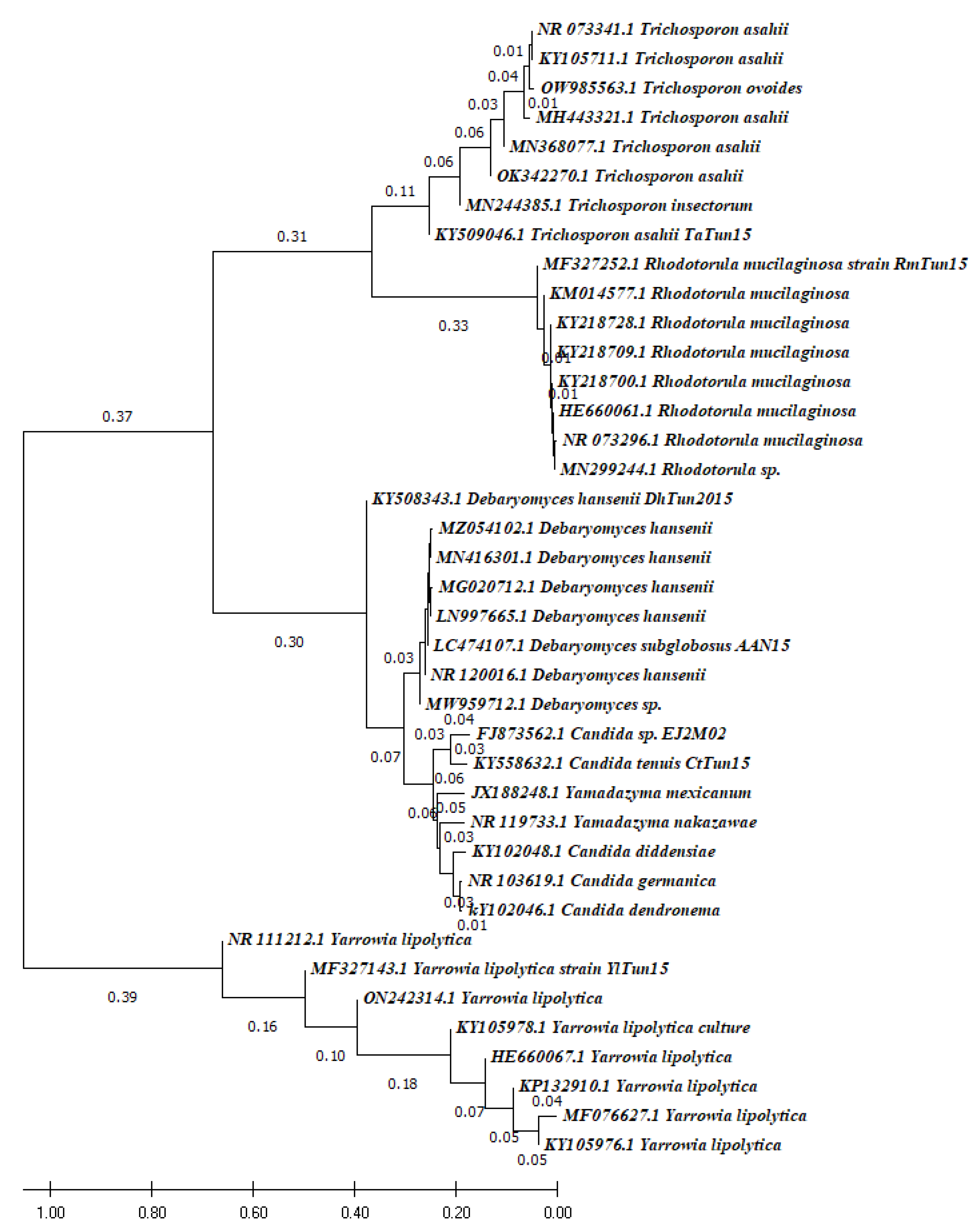
3.3. Phylogenic Analysis
3.4. Biochemical Characterization
3.4.1. Ash Content
3.4.2. Mineral Profile
3.4.3. Carbohydrate Content
3.4.4. Lipid Content
3.4.5. Fatty Acid Profile
3.4.6. Protein Content
3.4.7. Amino Acid Content
3.4.8. Biogenic Amine Content
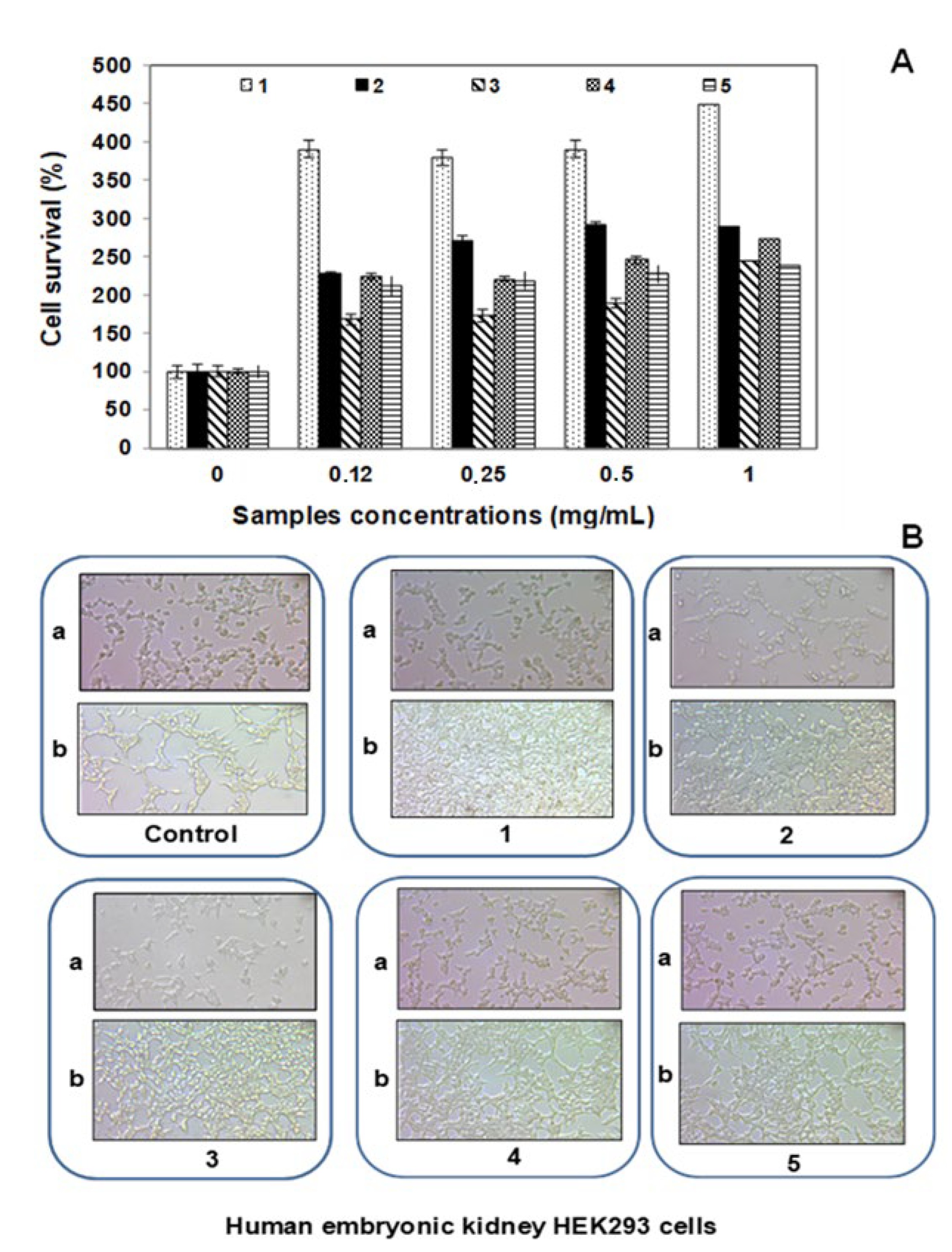
3.5. Cytotoxicity Effect of F5 Peptide on HEK293 Cells
3.6. Potential Application of Marine Yeast
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gadanho, M.; Sampaio, P.J. Occurrence and diversity of yeasts in the mid-Atlantic ridge hydrothermal fields near the Azores Archipelago. Microb. Ecol. 2005, 50, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Kutty, S.N.; Philip, R. Marine yeast-a review. Yeast 2008, 25, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.M.; Liu, T.T.; Chi, Z.; Liu, G.L.; Wang, Z.P. Occurrence and diversity of yeasts in the mangrove ecosystems in Fujian, Guangdong and Hainan provinces of China. Ind. J. Microbiol. 2012, 52, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Nagano, Y.; Nagahama, T. Fungal diversity in deep-sea extreme environments. Fungal Ecol. 2012, 5, 463–471. [Google Scholar] [CrossRef]
- Kandasamy, K.; Alikunhi, N.M.; Sunbramanian, M. Yeast in marine and estuarine environments. J. Yeast Fungal Res. 2012, 3, 74–82. [Google Scholar]
- Zhang, X.; Hua, M.X.; Song, C.L.; Chi, Z.M. Occurrence and diversity of marine yeasts in Antarctica environments. J. Ocean. Univ. China. 2012, 11, 70–74. [Google Scholar] [CrossRef]
- Chi, Z.; Liu, G.; Lu, Y.; Jiang, H.; Chi, Z.-M. Bio-products produced by marine yeasts and their potential applications. Bioresour. Technol. 2016, 202, 244–252. [Google Scholar] [CrossRef]
- Ahmed, I.; Haroon, A.; Khan, M.N.; Asadullah Shah, H.; Nadeem, A.; Saeed, F.; Ullah Shah, S.R.; Abbasi, M.A.; Buzdar, M.A. Occurrence and biodiversity of marine yeast in mangrove ecosystem of Shabi Creek, Gwadar-Pakistan. Pure Appl. Biol. 2019, 8, 680–687. [Google Scholar] [CrossRef]
- Arias, C.R.; Burns, J.K.; Friedrich, L.M.; Goodrich, R.M.; Parish, M.E. Yeast species associated with orange juice: Evaluation of different identification methods. Appl. Environ. Microb. 2002, 68, 1955–1961. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.; Rodriguez, M.; Sanchez, B.; Aranda, E.; Cordoba, J. DNA typing methods for differentiation of yeasts related to dry-cured meat products. Int. J. Food Microbiol. 2006, 107, 48–58. [Google Scholar] [CrossRef]
- Senses-Ergul, S.; Ágoston, R.; Belák, Á.; Deák, T. Characterization of some yeasts isolated from foods by traditional and molecular tests. Int. J. Food Microbiol. 2006, 108, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Akpınar, O.; Ucar, F.; Yalcın, H.T. Screening and regulation of alkaline extracellular protease and ribonuclease production of Yarrowia lipolytica strains isolated and identified from different cheeses in Turkey. Ann. Microbiol. 2011, 61, 907–915. [Google Scholar] [CrossRef]
- Lopandic, K.; Tiefenbrunner, W.; Gangl, H.; Mandl, K.; Berger, S.; Leitner, G.; Abd-Ellah, G.A.; Querol, A.; Gardner, R.C.; Sterflinger, K.; et al. Molecular profiling of yeasts isolated during spontaneous fermentations of Austrian wines. FEMS Yeast Res. 2008, 8, 1063–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of the ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. A Van Leeuw. 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Fell, J.W. Collection and identification of marine yeast in Methods. Microbiology 2001, 30, 347–356. [Google Scholar]
- Nagahama, T. Yeast Biodiversity in Freshwater, marine and deep-sea environments. Chap12. Biodivers. Ecophysiol. Yeasts 2006, 241–262. [Google Scholar] [CrossRef]
- Kaur, J.; Kaur, S.; Kumar, M.; Krishnan, P.; Priya Minhas, A. Studies on production, optimization and machine learning-based prediction of biosurfactant from Debaryomyces hansenii CBS767. Int. J. Sci. Environ. Technol. 2021, 19, 8465–8478. [Google Scholar] [CrossRef]
- Loeto, D.; Jongman, M.; Lekote, L.; Muzila, M.; Mokomane, M.; Motlhanka, K.; Zhou, N. Biosurfactant production by halophilic yeasts isolated from extreme environments in Botswana. FEMS Microbiol. Lett. 2021, 368, fnab146. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M. The Superiority of Yeast Secondary Metabolites, from Industrial applications, biological activities to Pharmaceutical potential. J. Pharmacogn. Phytochem. 2022, 14, 1. [Google Scholar]
- Hassabo, A.A.; Ibrahim, E.I.; Ali, B.A.; Emam, H.E. Anticancer effects of biosynthesized Cu2O nanoparticles using marine yeast. Biocatal. Agric. Biotechnol. 2022, 39, 102261. [Google Scholar] [CrossRef]
- Hassan, W.M.S.; Abdrabo, M.A.A.; Elsayis, A. Microbial Pigments: Sources and Applications in the Marine Environment. Egypt. J. Aquat. Biol. Fish. 2022, 26, 99–124. [Google Scholar]
- Li, H.; Huang, L.; Zhang, Y.; Yan, Y. Production, characterization and immunomodulatory activity of an extracellular polysaccharide from Rhodotorula mucilaginosa YL-1 isolated from sea salt field. Mar. Drugs. 2020, 18, 595. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kwon, K.K.; Kang, S.G.; Cha, S.S.; Kim, S.J.; Lee, J.H. Approaches for novel enzyme discovery from marine environments. Curr. Opin. Biotechnol. 2010, 21, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Zaky, A.S.; Tucker, G.A.; Daw, Z.Y.; Du, C. Marine yeast isolation and industrial application. FEMS Yeast Res. 2014, 14, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Zaky, A.S. Introducing a Marine Biorefinery System for the Integrated Production of Biofuels, High-Value-Chemicals, and Co-Products: A Path Forward to a Sustainable Future. Processes 2021, 9, 1841. [Google Scholar] [CrossRef]
- Gupta, A.; Vongsvivut, J.; Barraow, C.J.; Puri, M. Molecular identification of marine yeast and its spectroscopic analysis establishes unsaturated fatty acid accumulation. J. Biosci. Bioeng. 2012, 114, 411–417. [Google Scholar] [CrossRef]
- Sarkar, A.; Bhaskara Rao, K.V. Marine yeast: A potential candidate for Biotechnological applications-a review. Asian J. Microbiol. Biotech. Env. Sci. 2016, 18, 627–634. [Google Scholar]
- Anupama, R.; Ravindra, P. Value-added food. Biotechnol. Adv. 2000, 18, 459–479. [Google Scholar] [CrossRef]
- Chi, Z.; Yan, K.; Gao, L.; Li, J.; Wang, X.; Wang, L. Diversity of marine yeasts with high protein content and evaluation of their nutritive compositions. J. Mar. Biol. Assoc. UK 2008, 88, 1347–1352. [Google Scholar] [CrossRef]
- Bessadok, B.; Masri, M.; Brück, T.; Sadok, S. Characterization of the Crude Alkaline Extracellular Protease of Yarrowia lipolytica YlTun15. J. FisheriesSciences.Com. 2017, 11, 19–24. [Google Scholar] [CrossRef]
- Bessadok, B.; Santulli, A.; Brück, T.; Sadok, S. Species disparity response to mutagenesis of marine yeasts for the potential production of biodiesel. Biotechnol Biofuels. 2019, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vita-Marquez, M.A.; Lira, S.P.; Berlick, R.G.S. A multi-screening approch for marine-derived fungal metabolite and the isolation of cyclodepsiptides from Beauveria felina. Quim. Nova. 2008, 31, 1099–1103. [Google Scholar] [CrossRef] [Green Version]
- Masuda, K.; Guo, X.-F.; Uryu, N. Isolation of marine yeasts collected from the Pacific Ocean Showing a high production of γ-Aminobutyric acid. Biosci. Biotechnol. Biochem. 2008, 72, 3265–3272. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Yanagida, F.; Chen, L.-Y. Isolation of marine yeasts from coastal waters of northeastern Taiwan. Aquat. Biol. 2009, 8, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Rhishipal, R.; Philip, R. Selection of marine yeasts for the generation of single cell protein from prawn-shell waste. Bioresour. Technol. 1998, 65, 255–256. [Google Scholar] [CrossRef]
- Minegishi, H.; Miura, T.; Yoshida, Y.; Usami, R.; Abe, F. Phylogenetic analysis of pectin degrading yeasts from deep-sea environments. J. Jpn. Soc. Extrem. 2006, 5, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Burgaud, G.; le Calvez, T.; Arzur, D.; Vandenkoornhuyse, P.; Barbier, G. Diversity of culturable marine filamentous fungi from deep-sea hydrothermal vents. Environ. Microbiol. 2009, 11, 1588–1600. [Google Scholar] [CrossRef]
- Burgaud, G.; Arzur, D.; Durand, L.; Cambon-Bonavita, M.A.; Barbier, G. Marine culturable yeasts in deep-sea hydrothermal vents: Species, richness and association with fauna. FEMs Microbiol. Ecolo. 2010, 73, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Bharathi, S.; Saravanan, D.; Radhakrishnan, M.; Balagurunathan, R. Bioprocessing of marine teast with special reference to Inulinase production. Int. J. Chem. Tech Res. 2011, 3, 1514–1519. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: New York, NY, USA, 1989. [Google Scholar]
- Josefa, M.C.J.; Lyibia, M.C.; Sergio, M.R. Molecular characterization and ecological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995.
- Norme Tunisienne: Standard NT.09.193; Qualité de l’eau—Dosage D’éléments Choisis par Spectroscopie D’émission Optique avec Plasma Induit par Haute Fréquence (ICP-OES). National Institute of Standardization and Industrial Property: Cité El Khadra, Tunis, 2010.
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Van Hille, R.P.; Harrison, S.T.L. Selection of Direct Transesterification as the Preferred Method for Assay of Fatty Acid Content of Microalgae. Lipids 2010, 45, 1053–1060. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Bartolomeo, M.; Maisao, F. Validation of a reversed-phase HPLC Method for quantitative amino acid analysis. J. Biomol. Tech. 2006, 17, 131–137. [Google Scholar]
- Ouijifard, A.; Seyfadadi, J.; Kenari, A.A.; Rezaei, M. Growth and apparent digestibility of nutrient, fatty acids and amino acids in Pacific white shrimp, Litopenaeusvannamei, fed diets with rice protein concentrate as total and partial replacement of fish meal. J. Aquac. 2012, 342–343, 56–61. [Google Scholar] [CrossRef]
- International Standard Organization: ISO 19343; Microbiology of the food chain—Detection and quantification of histamine in fish and fishery products—HPLC method. International Organization for Standardization: Geneva, Switzerland, 2017.
- Elloumi-Mseddi, J.; Jemel-Oualha, I.; Beji, A.; Hakim, B.; Aifa, S. Effect of estradiol and clomiphene citrate on Erk activation in breast cancer cells. J. Recept. Signal. Transduct. Res. 2015, 35, 202–206. [Google Scholar] [CrossRef]
- Mikucki, J.A.; Liu, Y.; Delwiche, M.; Colwell, F.S.; Boone, D.R. Isolation of methanogen from deep sediments that contain methane hydrate, and description of Methanoculleussubmarinus sp. Nov. Appl. Environ. Microbiol. 2003, 69, 3311–3316. [Google Scholar] [CrossRef] [Green Version]
- Hirimuthugoda, N.Y.; Chi, Z.; Wu, L. Des levures probiotiques présentant une activité phytases, découvertes dans l’appareil digestif d’holothuries. La Bêche-De-Mer Bull. De La CPS 2007, 26, 31–33. [Google Scholar]
- Wang, X.H.; Chi, Z.M.; Yue, L.; Li, J. Purification and characterization of killer toxin from a marine yeast Pichia anomala YF07b against the pathogenic yeast in crab. Curr. Microbiol. 2007, 55, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Chi, Z.; Li, J.; Gao, L.; Gong, F. Inulinase production by the marine yeast Cryptococcus aureus G7a and inulin hydrolysis by the crude inulinase. Process Biochem. 2007, 42, 805–811. [Google Scholar] [CrossRef]
- Shen, H.; Wei, Y.; Wang, X.; Xu, C.; Shao, X. The marine yeast Sporidioboluspararoseus ZMY-1 has antagonistic properties against Botrytis cinerea in vitro and in strawberry fruit. Postharvest Biol. Technol. 2019, 150, 1–8. [Google Scholar] [CrossRef]
- Balan, S.; Ganesh Kumar, C.; Jayalakshmi, S. Physicochemical, structural and biological evaluation of Cybersan (trigalactomargarate), a new glycolipid biosurfactant produced by a marine yeast, Cyberlindnerasaturnus strain SBPN-27. Process Biochem. 2019, 80, 171–180. [Google Scholar] [CrossRef]
- Fotedar, R.; Kolecka, A.; Boekhout, T.; Fell, J.W.; Anand, A.; Al Malaki, A.; Zeyara, A.; Al Marri, M. Naganishiaqatarensis sp. Nov., a novel basidomycetous yeast species from a hypersaline marine environment in Qatar. Int. J. Syst. Evol. Microbiol. 2018, 68, 2924–2929. [Google Scholar] [CrossRef]
- Dhaliwal, M.K.; Chandra, N. Isolation of carotenoids producing marine red yeasts. Indian J. Geo-Mar. Sci. 2014, 45, 1029–1034. [Google Scholar]
- Bogusławska-Wąs, E.; Dłubała, A.; Laskowska, M. The role of Rhodotorula mucilaginosa in selected biological process of wild fish. Fish Physiol. Biochem. 2018, 45, 511–521. [Google Scholar] [CrossRef] [Green Version]
- El-Baz, A.F.; El-Enshasy, H.A.; Shetaia, Y.M.; Mahrous, H.; Othman, N.Z.; Yousef, A.E. Semi-industrial scale production of a new yeast with probiotic traits Cryptococcus sp. YMHS, isolated from the red sea. Probiotics Antimicrob. Proteins 2017, 10, 77–88. [Google Scholar] [CrossRef]
- De Leo, F.; Lo Giudice, A.; Alaimo, C.; De Carlo, G.; Rappazzo, A.C.; Graziano, M.; De Domenico, E.; Urzì, C. Occurrence of the black yeast Hortaea werneckii in the Mediterranean Sea. Extremophiles 2019, 23, 9–17. [Google Scholar] [CrossRef]
- Filippousi, R.; Antoniou, D.; Tryfinopoulou, P.; Nisiotou, A.A.; Nychas, G.J.; Koutinas, A.A.; Papanikolaou, S. Isolation, identification and screening of yeasts towards their ability to assimilate biodiesel-derived crude glycerol: Microbial production of polyols, endopolysaccharides and lipid. J. Appl. Microbiol. 2019, 127, 1080–1100. [Google Scholar] [CrossRef] [PubMed]
- Scorzetti, G.; Fell, J.W.; Fonseca, A.; Statzell-Tallman, A. Systematics of basidiomycetous yeasts: A comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS Yeast Res. 2002, 2, 495–517. [Google Scholar] [CrossRef]
- Suh, S.O.; Blackwell, M.; Kurtzman, C.P.; Lachance, M.A. Phylogenetics of Saccharomycetales, the ascomycete yeasts. Mycologia 2006, 98, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, T.; Hamamoto, M.; Nakase, T.; Takami, H.; Horikoshi, K. Distribution and identification of red yeasts in deep-sea environments around the northwest Pacific Ocean. Antonie Van Leeuwenhoek 2001, 80, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, T.; Abdel-Wahab, M.A.; Nogi, Y.; Miyazaki, M.; Uematsu, K.; Hamamoto, M.; Horikoshi, K. Dipodascus tetrasporeus sp. nov., an ascosporogenous yeast isolated from deep-sea sediments in the japan trench. Int. J. Syst. Evol. Microbiol. 2008, 58, 1040–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khambhaty, Y.; Upadhyay, D.; Kriplani, Y.; Joshi, N.; Mody, K.; Gandhi, M.R. Bioenerg Res Bioethanol from macroalgal biomass: Utilization of marine yeast for production of the same. BioEnergy Res. 2013, 6, 188–195. [Google Scholar] [CrossRef]
- Kathiresan, K.; Saravanakumar, K.; Senthilraja, P. Bio-ethanol production by marine yeasts isolated from coastal mangrove sediment. Int. Multidiscip. Res. J. 2011, 1, 19–24. [Google Scholar]
- Burgaud, G.; Arzur, D.; Sampaio, J.P.; Barbier, G. Candida oceani sp. nov., a novel yeast isolated from a Mid-Atlantic Ridge hydrothermal vent (−2300 meters). Antonie Van Leeuwenhoek 2011, 100, 75–82. [Google Scholar] [CrossRef]
- Zaky, A.S.; Greetham, D.; Louis, E.J.; Tucker, G.A.; Du, C. A New Isolation and Evaluation Method for Marine-Derived Yeast spp. with Potential Applications in Industrial Biotechnology. J. Microbiol. Biotechnol. 2016, 26, 1891–1907. [Google Scholar] [CrossRef] [Green Version]
- Raggi, P.; Lopez, P.; Diaz, A.; Carrasco, D.; Silva, A.; Velez, A.; Navarrete, P.A. Debaryomyces hansenii and, Rhodotorula mucilaginosa comprised the yeast core gut microbiota of wild and reared carnivorous salmonids, croaker and yellowtail. Environ. Microbiol. 2014, 16, 2791–2803. [Google Scholar] [CrossRef]
- Leyton, A.; Flores, L.; Mäki-Arvel, P.; Lienqueo, M.E.; Shene, C. Macrocystis pyrifera source of nutrients for the production of carotenoids by a marine yeast Rhodotorula mucilaginosa. J. Appl. Microbiol. 2019, 127, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Lario, L.D.; Chaud, L.; Almeida Mdas, G.; Converti, A.; Durães Sette, L.; Pessoa, A. Production, purification, and characterization of an extracellular acid protease from the marine Antarctic yeast Rhodotorula mucilaginosa L7. Fungal Biol. 2015, 119, 1129–1136. [Google Scholar] [CrossRef]
- Meng, X.; Yang, J.; Xu, X.; Zhang, L.; Nie, Q.; Xian, M. Biodiesel production from oleaginous microorganisms. Renew. Energ. 2009, 34, 1–5. [Google Scholar] [CrossRef]
- Sarlin, P.J.; Rosamma, P. Marine yeasts as feed supplement for Indian white prawn Fenneropenaeus indicus: Screening and Testing the Efficacy. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 55–70. [Google Scholar] [CrossRef]
- Sarlin, P.J.; Rosamma, P. Efficacy of marine yeasts as feed supplement for Fenneropenaeus indicus in culture systems. Int. J. Curr. Life Sci. 2018, 7, 927–933. [Google Scholar]
- Andlid, T.; Larsson, C.; Liljenberg, C.; Marison, I.; Gustafsson, L. Enthalpy content as a function of lipid accumulation in Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 1995, 42, 818–825. [Google Scholar] [CrossRef]
- Brown, M.R.; Barrett, S.M.; Volkman, J.K.; Nearhos, S.P.; Nell, J.A.; Allan, G.L. Biochemical composition of new yeasts and bacteria evaluated as food for bivalve aquaculture. Aquaculture 1996, 143, 341–360. [Google Scholar] [CrossRef]
- Jones, R.P.; Greenfield, P.F. A review of yeast ionic nutrition, I.: Growth and fermentative requirements. Process. Biochem. 1994, 4, 48–59. [Google Scholar]
- Burke, R.M.; Jennings, D.H. Effect of sodium chloride on growth characteristics of the marine yeast Debaryomyces hansenii in batch and continuous culture under carbon and potassium limitation. Mycol. Res. 1990, 94, 378–388. [Google Scholar] [CrossRef]
- Cong, L.; Chi, Z.; Li, J.; Wang, X. Enhanced carotenoid production by a mutant of the marine yeast Rhodotorula sp. Hidai. J. Ocean. Univ. China 2006, 6, 66–71. [Google Scholar] [CrossRef]
- Zinjarde, S.S.; Bhagyashree, V.K.; Vishwasrao, P.V.; Kumar, A.R. Morphogenetic Behavior to tropical marine yeast Yarrowia lipolytica in response to hydrophobic substrates. J. Microbio. Biotechnol. 2008, 18, 1522–1528. [Google Scholar]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Effect of addition of Tween 80 and potassium dihydrogenphosphate to basal medium on the isolation of marine eukaryotes, thraustochytrids. J. Biosci. Bioeng. 2008, 105, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Sarlin, P.J.; Philip, R. A molasses-based fermentation medium for marine yeast biomass production. Int. J. Res. Mar. Sci. 2013, 2, 39–44. [Google Scholar]
- Sarlin, P.J. Marine Yeasts as Source of Single Cell Protein and Immunostimulant for Application in Penaeid Prawn Culture Systems. Chapiter 3. Ph.D. Thesis, Cochin University of Science and Technology, Cochin, India, 2005; pp. 1–258. [Google Scholar]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Ageitos, J.J.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef]
- Banat, I.M.; Franzetti, A.; Gandolfi, I.; Bestetti, G.; Martinotti, M.G.; Fracchia, L.; Smyth, T.J.; Marchant, R. Microbial biosurfactants production, applications and future potential. Appl. Microbiol. Biotechnol. 2010, 87, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Dalmas Neto, C.J.; Sydney, E.B.; Candeo, E.S.; de Souza, E.B.S.; Camargo, D.; Sydney, A.C.N.; Soccol, C.R. New Method for the Extraction of Single-Cell Oils from Wet Oleaginous Microbial Biomass: Efficiency, Oil Characterization and Energy Assessment. Waste Biomass Valorization 2020, 11, 3443–3452. [Google Scholar] [CrossRef]
- Bommareddy, R.R.; Sabra, W.; Maheshwari, G.; Zeng, A.P. Metabolic network analysis and experimental study of lipid production in Rhodosporidium toruloides grown on single and mixed substrates. Microb. Cell Fact. 2015, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Z.; Zanotti, M.; Wang, X.; Ducey, C.; Liu, Y. Evaluation of lipid accumulation from lignocellulosic sugars by Mortierellaisabellina for biodiesel production. Bioresour. Technol. 2012, 110, 198–205. [Google Scholar] [CrossRef]
- Enshaeieh, M.; Abdoli, A.; Madani, M. Single Cell Oil (SCO) Production by Rhodotorula mucilaginosa and Its Environmental Benefits. J. Agr. Sci. Tech. 2015, 17, 387–400. [Google Scholar]
- Enshaeieh, M.; Abdoli, A.; Nahvi, I.; Madani, M. Medium optimization for biotechnological production of single cell oil using Candida gali and Yarrowia lipolytica M7. J. Cell Mol. Res. 2013, 5, 17–23. [Google Scholar]
- Enshaeieh, M.; Nahvi, I.; Madani, M. Improving microbial oil production with standard and native oleaginous yeasts by using Taguchi design. Int. J. Environ. Sci. Technol. 2014, 11, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for single cell oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef]
- Angerbauer, C.; Siebenhofer, M.; Mittelbach, M.; Guebitz, G.M. Conversion of sewage sludge into lipids by Lipomycesstarkeyi for biodiesel production. Bioresour. Technol. 2008, 99, 3051–3056. [Google Scholar] [CrossRef]
- Li, Q.; Du, W.; Liu, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef]
- Gientka, I.; Kieliszek, M.; Jermacz, K.; Błażejak, S. Identification and Characterization of Oleaginous Yeast Isolated from Kefir and Its Ability to Accumulate Intracellular Fats in Deproteinated Potato Wastewater with Different Carbon Sources. Biomed. Res. Int. 2017, 2017, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellou, S.; Baeshen, M.N.; Elazzazy, A.M.; Aggeli, D.; Sayegh, F.; Aggelis, G. Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 2014, 32, 1476–1493. [Google Scholar] [CrossRef]
- Kang, C.K.; Park, H.Y.; Kim, M.C.; Lee, W.J. Use of marine yeasts as an available diet for mass cultures of Moinamacrocopa. Aquac. Res. 2006, 37, 1227–1237. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, D.; Yang, Q.; Wang, P. Enhancing carotenoid production in Rhodotorula mucilaginosa KC8 by combining mutation and metabolic engineering. Ann. Microbiol. 2017, 67, 425–431. [Google Scholar] [CrossRef]
- Ghanavati, H.; Nahvi, I.; Karimi, K. Organic fraction of municipal solid waste as a suitable feedstock for the production of lipid by oleaginous yeast Cryptococcus aerius. Waste Manag. 2015, 38, 141–148. [Google Scholar] [CrossRef]
- Spier, F.; Buffon, J.G.; Burkert, C.A. Bioconversion of raw glycerol generated from the synthesis of biodiesel by different oleaginous yeasts: Lipid content and fatty acid profile of biomass. Indian J. Microbiol. 2015, 55, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimura, A.; Takashima, M.; Sugita, T.; Endoh, R.; Kikukawa, M.; Yamaguchi, S.; Sakuradani, E.; Ogawa, J.; Ohkuma, M.; Shima, J. Cryptococcus terricola is a promising oleaginous yeast for biodiesel production from starch through consolidated bioprocessing. Sci. Rep. 2014, 4, 4776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ren, H. Microbial oil production by Rhodotorula glutinis CICC 31643 using sugar cane molasses. J. Renew. Sustain. Energy 2014, 6, 104–108. [Google Scholar] [CrossRef]
- Chi, Z.M.; Liu, Z.Q.; Gao, L.M.; Gong, F.; Ma, C.L.; Wang, X.H. Marine yeasts and their applications in mariculture. J. Ocean. Univer China 2006, 5, 251–257. [Google Scholar]
- Cui, W.; Wang, Q.; Zhang, F.; Zhang, S.C.; Chi, Z.M.; Madzak, C. Direct conversion of inulin into single cell protein by the engineered Yarrowia lipolytica carrying inulinase gene. Proc. Biochem. 2011, 46, 1442–1448. [Google Scholar] [CrossRef]
- Zhang, T.; Chi, Z.M.; Sheng, J. A highly thermosensitive and permeable mutant of the marine yeast Cryptococcus aureus G7a potentially useful for single-cell protein production and its nutritive components. Mar. Biotechnol. 2009, 12, 280–286. [Google Scholar] [CrossRef]
- Jach, M.E.; Serefko, A.; Ziaja, M.; Kieliszek, M. Yeast Protein as an Easily Accessible Food Source. Metabolites 2022, 12, 63. [Google Scholar] [CrossRef]
- TenBrink, B.; Damink, C.; Joosten, H.M.L.J.; Huis Veld, J.H.J. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef]
- Islam, M.A.; Pang, J.; Meng, F.; Li, Y.; Xu, N.; Yang, C.; Liu, J. Putrescine, spermidine, and spermine play distinct roles in rice salt tolerance. J. Integr. Agric. 2020, 19, 643–655. [Google Scholar] [CrossRef]
- Goes, E.C.; Cardoso Dal Pont, G.; Oliveira, P.R.; Rocha, C.; Maiorka, A. Effects of putrescine injection in broiler breeder eggs. J. Anim. Physiol. Anim. 2020, 105, 294–304. [Google Scholar] [CrossRef]
- Schwarz, C.; Horn, N.; Benson, G.; Calzado, I.W.; Wurdack, K.; Pechlaner, R.; Grittner, U.; Wirth, M.; Flöel, A. Spermidine Intake is Associated with Cortical Thickness and Hippocampal Volume in Older Adults. NeuroImage 2020, 221, 117132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Chen, Y.; Jin, L.W.; Ye, H.Y.; Liu, G. Cytotoxicity and Genotoxicity in Human Embryonic Kidney Cells Exposed to Surface Modify Chitosan Nanoparticles Loaded with Curcumin. AAPS PharmSciTech 2016, 17, 1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, G.; Wang, X.; Jiang, J.; Zhang, C.; Chi, M.; Ju, Y.; Shi, C. Copper stress response in yeast Rhodotorula mucilaginosa AN5 isolated from sea ice, Antarctic. Microbiologyopen 2018, 8, e00657. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Zong, M.H.; Wu, H. Efficient lipid production with Trichosporon fermantans and its use for biodiesel preparation. Bioresour. Technol. 2008, 99, 7881–7885. [Google Scholar] [CrossRef] [PubMed]
- Easterling, E.R.; French, W.T.; Hernandez, R.; Licha, M. The effect of glycerol as a sole and secondary substrate on the growth and fatty acid composition of Rhodotorula glutinis. Bioresour. Technol. 2009, 100, 356–361. [Google Scholar] [CrossRef]
- Li, M.; Liu, G.L.; Chi, Z.; Chi, Z.M. Single cell oil production from hydrolysate of cassava starch by marine-derived yeast Rhodotorula mucilaginosa TJY15a. Biomass Bioenergy 2010, 34, 101–107. [Google Scholar] [CrossRef]
- Galafassi, S.; Cucchetti, D.; Pizza, F.; Franzosi, G.; Bianchi, D.; Compagno, C. Lipid production for second-generation biodiesel by the oleaginous yeast Rhodotorula graminis. Bioresour. Technol. 2012, 111, 398–403. [Google Scholar] [CrossRef]
- Tanimura, A.; Takashima, M.; Sugita, T.; Endoh, R.; Kikukawa, M.; Yamaguchi, S.; Sakuradani, E.; Ogawa, J.; Shima, J. Selection of oleaginous yeasts with high lipid productivity for practical biodiesel production. Bioresour. Technol. 2014, 153, 230–253. [Google Scholar] [CrossRef]
- Munch, G.; Sestric, R.; Sparling, R.; Levin, D.B.; Cicek, N. Lipid production in the under-characterized oleaginous yeasts, Rhodosporidium babjevae and Rhodosporidium diobovatum, from biodiesel-derived waste glycerol. Bioresour. Technol. 2015, 185, 49–55. [Google Scholar] [CrossRef]
- Arous, F.; Frikha, F.; Triantaphyllidou, I.E.; Aggelis, G.; Nasri, M.; Mechichi, T. Potential utilization of agro-industrial wastewaters for lipid production by the oleaginous yeast Debaryomyces etchellsii. J. Clean. Prod. 2016, 133, 899–909. [Google Scholar] [CrossRef]
- Brar, K.K.; Sarma, A.K.; Aslam, M.; Polikarpov, I.; Chadha, B.S. Potential of oleaginous yeast Trichosporon sp., for conversion of sugarcane bagasse hydrolysate into biodiesel. Bioresou. Technol. 2017, 242, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Guerfali, M.; Ayadi, I.; Belhassen, A.; Gargouri, A.; Belghith, H. Single cell oil production by Trichosporon cutaneum and lignocellulosic residues bioconversion for biodiesel synthesis. Process Saf. Environ. Prot. 2018, 113, 292–304. [Google Scholar] [CrossRef]
- Katre, G.; Joshi, C.; Khot, M.; Zinjarde, S.; RaviKumar, A. Evaluation of single cell oil (SCO) from a tropical marine yeast Yarrowia lipolytica NCIM 3589 as a potential feedstock for biodiesel. AMB Exp. 2012, 2, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katre, G.; Raskar, S.; Zinjarde, S.; Kumar, R.; Kulkarni, B.D.; Kumar, A.R. Optimization of the in-situ transesterification step for biodiesel production using biomass of Yarrowia lipolytica NCIM 3589 grown on waste cooking oil. Energy 2018, 142, 944–952. [Google Scholar] [CrossRef]
- Jiru, T.M.; Steyn, L.; Pohl, C.; Abate, D. Production of single cell oil from cane molasses by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 as a biodiesel feedstock. Chem. Cent. J. 2018, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.W.; Hu, C.Y.; Liang, W.S. A cost-efficient way to obtain lipid accumulation in the oleaginous yeast Rhodotorula glutinis using supplemental waste cooking oils (WCO). J. Taiwan Inst. Chem Eng. 2019, 97, 80–87. [Google Scholar] [CrossRef]
- Dobrowolski, A.; Drzymała, K.; Rzechonek, D.A.; Mituła, P.; Mirończuk, A.M. Lipid production from waste materials in seawater-based medium by the yeast Yarrowia lipolytica. Front. Microbiol. 2019, 10, 547. [Google Scholar] [CrossRef] [Green Version]
- Senthilraja, P.; Kathiresan, K.; Saravanakumar, K. Comparative analysis of bioethanol production by different strains of immobilized marine yeast. J. Yeast Fungal Res. 2011, 8, 113–116. [Google Scholar]
- Tsigie, Y.A.; Wu, C.H.; Huynh, L.H.; Ismadji, S.; Ju, Y.H. Bioethanol production from Yarrowia lipolytica Po1g biomass. Bioresour. Technol. 2013, 145, 210–216. [Google Scholar] [CrossRef]
- Gonzalez-lopez, C.I.; Szabo, R.; Blanchin-Roland, S.; Gaillardin, C. Genetic control of extracellular protease synthesis in the yeast Yarrowia lipolytica. Genetics 2001, 160, 417–427. [Google Scholar]
- Li, J.; Chi, Z.; Wang, X.; Peng, Y.; Chi, Z. The selection of alkaline protease-producing yeast from marine environments and evaluation of their bioactive peptide production. Chin. J. Oceanol. Limnol. 2009, 27, 753. [Google Scholar] [CrossRef]
- Akpinar, O.; Uçar, F.B. Molecular characterization of Yarrowia lipolytica strains isolated from different environments and lipase profiling. Turk. J. Biol. 2013, 37, 249–258. [Google Scholar] [CrossRef]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Jiao, L. Engineering Yarrowia lipolytica to Simultaneously Produce Lipase and Single Cell Protein from Agro-industrial Wastes for Feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papouskova, K.; Sychrova, H. The co-action of osmotic and high temperature stresses results in a growth improvement of Debaryomyces hansenii cells. Int. J. Food Microbiol. 2007, 118, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Angulo, C.; Maldonado, M.; Delgado, K.; Reyes-Becerril, M. Debaryomyces hansenii up regulates superoxide dismutase gene expression and enhances the immune response and survival in Pacific red snapper (Lutjanus peru) leukocytes after Vibrio parahaemolyticus infection. Dev. Comp. Immunol. 2017, 71, 18–27. [Google Scholar] [CrossRef]
- Shanmugam, V.K.; Gopalakrishnan, D. Screening and partial purification for the production of lignianse enzyme from the fungal isolate Trichosporon asahii. Braz. Arch. Biol. Technol. 2016, 59, e160220. [Google Scholar] [CrossRef]
- Duarte, S.H.; de Andrade, C.C.P.; Ghiselli, G.; Maugeri, F. Exploration of Brazilian biodiversity and selection of a new oleaginous yeast strain cultivated in raw glycerol. Bioresour. Technol. 2013, 138, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Martuscelli, M.; Arfelli, G.; Manetta, A.C.; Suzzi, G. Biogenic amines content as a measure of the quality of wines of Abruzzo (Italy). Food Chem. 2013, 140, 590–597. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Xia, Y.; Zhuang, X.; Chu, W. Isolation, Identification of Carotenoid-Producing Rhodotorula sp. from Marine Environment and Optimization for Carotenoid Production. Mar. Drugs. 2019, 17, 161. [Google Scholar] [CrossRef] [Green Version]
- Machado, W.R.C.; Silva, L.G.; da-Vanzela, E.S.L.; Del Bianchi, V.L. Evaluation of the process conditions for the production of microbial carotenoids by the recently isolated Rhodotorula mucilaginosa URM 7409. Braz. J. Food Technol. 2019, 22, e2018267. [Google Scholar] [CrossRef]
- Zhang, C.; Shen, H.; Zhang, X.; Yu, X.; Wang, H.; Xiao, S.; Wang, J.; Zhao, Z.K. Combined mutagenesis of Rhodosporidium toruloides for improved production of carotenoids and lipids. Biotechnol. Lett. 2019, 38, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Bryś, J.; Reczek, L.; Pobiega, K. Effect of exogenous stress factors on the biosynthesis of carotenoids and lipids by Rhodotorula yeast strains in media containing agro-industrial waste. World J. Microbiol. Biotechnol. 2019, 35, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landolfo, S.; Chessa, R.; Zara, G.; Zara, S.; Budroni, M.; Mannazzu, I. Rhodotorula mucilaginosa C2.5t1 Modulates Carotenoid Content and CAR Genes Transcript Levels to Counteract the Pro-Oxidant Effect of Hydrogen Peroxide. Microorganisms 2019, 7, 316. [Google Scholar] [CrossRef] [Green Version]
- Selvi, P.S.; Iyer, P. Isolation and characterization of pigments from marine soil microorganisms. Inter. J. Life. Sci. Scienti. Res. 2019, 2003–2011. [Google Scholar]
- Tkáčová, J.; Klempová, T.; Čertík, M. Kinetic study of growth, lipid and carotenoid formation in β-carotene producing Rhodotorula glutinis. Chem. Pap. 2018, 72, 1193–1203. [Google Scholar] [CrossRef]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of β-carotene. Biotechnol. Bioeng. 2017, 115, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis—potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [Green Version]
- Yoo, K.M.; Moon, B. Comparative carotenoid compositions during maturation and their antioxidative capacities of three citrus varieties. Food Chem. 2016, 196, 544–549. [Google Scholar] [CrossRef]
- Grenfell-Lee, D.; Zeller, S.; Cardoso, R.; Pucaj, K. The safety of beta-carotene from Yarrowia lipolytica. Food Chem. Toxicol. 2014, 65, 1–11. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Certik, M.; Dvorakova, T.; Haronikova, A. Use of several waste substrates for carotenoid-rich yeast biomass production. J. Env. Manag. 2011, 95, 338–342. [Google Scholar] [CrossRef]
- Moliné, M.; Flores, M.R.; Libkind, D.; del Carmen Diéguez, M.; Farías, M.E.; van Broock, M. Photoprotection by carotenoid pigments in the yeast Rhodotorula mucilaginosa: The role of torularhodin. Photochem. Photobiol. Sci. 2010, 9, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Malisorn, C.; Suntornsuk, W. Optimization of β-carotene production by Rhodotorula glutinis DM28 in fermented radish brine. Bioresour. Technol. 2008, 99, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, B.P.; Gadre, R.V. Production of β-carotene by a mutant of Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2001, 55, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Saavedra, N.Y.; Ochoa, J.L. Copper-zinc superoxide dismutase from the marine yeast Debaryomyces hansenii. Yeast 1999, 15, 657–668. [Google Scholar] [CrossRef]
- Liu, G.; Yue, L.; Chi, Z.; Yu, W.; Chi, Z.; Madzak, C. The Surface Display of the Alginate Lyase on the Cells of Yarrowia lipolytica for Hydrolysis of Alginate. Mar. Biotechnol. 2009, 11, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Apte, M.; Sambre, D.; Gaikawad, S.; Joshi, S.; Bankar, A.; Kumar, A.R.; Zinjarde, S. Psychrotrophic yeast Yarrowia lipolytica NCYC 789 mediates the synthesis of antimicrobial silver nanoparticles via cell-associated melanin. AMB Express 2013, 3, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amornrattanapan, P. Riboflavin production by Candida tropicalis isolated from seawater. Sci. Res. Essays. 2013, 8, 43–47. [Google Scholar]
- Subramanian, M.; Alikunhi, N.M.; Kandasamy, K. In vitro synthesis of silver nanoparticles by marine yeasts from coastal mangrove sediment. Adv. Sci. Lett. 2010, 3, 428–433. [Google Scholar] [CrossRef]
- Soliman, H.; Elsayed, A.; Dyaa, A. Antimicrobial activity of silver nanoparticles biosynthesized by Rhodotorula sp. strain ATL72. Egypt. J. Basic Appl. Sci. 2018, 5, 228–233. [Google Scholar]
- Carvalho, T.; Pereira, A.; Finotelli, P.V.; Amaral, P.F.F. Palm oil fatty acids and carotenoids extraction with lipase immobililized in magnetic nanoparticles. Adv. Mater. Lett. 2018, 9, 643–646. [Google Scholar] [CrossRef]
- Libkind, D.; Brizzio, S.; van Broock, M. Rhodotorula mucilaginosa, a carotenoid producing yeast strain from a Patagonian high-altitude lake. Folia Microbiol. 2004, 49, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.S.; Vinusha, B.; Vijaya, C. Efficacy of the marine yeast Debaryomyces hansenii on growth of the shrimp Litopenaeusvannamei. Int. J. Res. Appl. Sci. Eng. Technol. 2017, 5, 1545–1547. [Google Scholar] [CrossRef]
- Opazo, R.; Fuenzalida, K.; Plaza-Parrochia, F.; Romero, J. Performance of Debaryomyces hansenii as a Diet for Rotifers for Feeding Zebrafish Larvae. Zebrafish 2017, 14, 187–194. [Google Scholar] [CrossRef]
- Medina-Córdova, N.; Rosales-Mendoza, S.; Hernández-Montiel, L.G.; Angulo, C. The potential use of Debaryomyces hansenii for the biological control of pathogenic fungi in food. Biol. Control 2018, 121, 216–222. [Google Scholar] [CrossRef]
- Alvarez-Sanchez, A.R.; Nolasco-Soria, H.; Pena-Rodriguez, A.; Mejia-Ruiz, H. In vitro digestibility of Yarrowia lipolytica Yeast and growth performance in White leg shrimp Litopenaeusvannamei. Turkish J. Fish. Aquat. Sci. 2018, 18, 395–404. [Google Scholar]
- Ma, Y.; Li, L.; Li, M.; Chen, W.; Bao, P.; Yu, Z.; Chang, Y. Effects of dietary probiotic yeast on growth parameters in juvenile sea cucumber, Apostichopus japonicus. Aquaculture 2019, 499, 203–211. [Google Scholar] [CrossRef]
- Zinjarde, S.S.; Pant, A. Emulsifier from a tropical marine yeast Yarrowia lipolytica NCIM 3589. J. Basic Microbial. 2002, 42, 67–73. [Google Scholar] [CrossRef]
- Amaral, P.F.F.; Lehocky, M.; Barros-Timmons, A.M.V.; Rocha-Leão, M.H.M.; Coelho, M.A.Z.; Coutinho, J.A.P. Cell surface characterization of Yarrowia lipolytica IMUFRJ 50682. Yeast 2006, 23, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Gusmão, C.A.B.; Rufino, R.D.; Sarubbo, L.A. Laboratory production and characterization of a new biosurfactant from Candida glabrata UCP 1002 cultivated in vegetable fat waste applied to the removal of hydrophobic contaminant. World J. Microbiol. Biotechnol. 2010, 26, 1683–1692. [Google Scholar] [CrossRef]
- Batista, R.M.; Rufino, R.D.; Luna, J.M.; Souza, J.E.G.; Sarubbo, L.A. Effect of medium components on the production of a biosurfactant from Candida tropicalis applied to the removal of hydrophobic contaminants in soil. Water Environ. Res. 2010, 82, 418–425. [Google Scholar] [CrossRef]
- Priji, P.; Unni, K.N.; Sajith, S.; Benjamin, S. Candida tropicalis BPU1, a novel isolate from the rumen of the Malabari goat, is a dual producer of biosurfactant and polyhydroxybutyrate. Yeast 2013, 30, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, H.; Hirai, A.; Minabe, T.; Obata, H. Stabilization of astaxanthin by a novel biosurfactant produced by Rhodotorula mucilaginosa KUGPP-1. Biocontrol. Sci. 2013, 18, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufino, R.D.; Luna, J.M.; Campos–Takaki, G.M.; Sarubbo, L.A. Characterization and properties of the biosurfactant produced by Candida lipolytica UCP 0988. Electron. J. Biotech. 2014, 17, 34–38. [Google Scholar] [CrossRef]
- Luna, J.M.; Santos, F.A.; Rufino, R.D.; Sarubbo, L.A. Production of a biosurfactant from Candida bombicola URM 3718 for environmental applications. Chem. Eng. Trans. 2016, 49, 583–588. [Google Scholar]
- Rubio-Ribeaux, D.; da Silva Andrade, R.F.; da Silva, G.S.; de Holanda, R.A.; Pele, M.A.; Nunes, P.; Vilar-Junior, J.C.; de Resende-Stoianoff, M.A.; Campos-Takaki, G.M. Promising biosurfactant produced by a new Candida tropicolis UCP 1613 strain using substrates from renewable resources. Afr. J. Microbiol. Res. 2017, 11, 981–991. [Google Scholar]



{kind=link}
{kind=link}
| Sample | Isolation Technique | Quantity (g; mL; cm) | Isolation Media |
|---|---|---|---|
| Sea water | Enrichment | 3 mL | MI; YNB; YPD |
| Filtration | 500 mL | MI-Agar | |
| Sediment | Enrichment | 2 g | MI; YPD |
| Direct culture | 0.1 g | MI-Agar | |
| Gills (S. aurata/D. labrax) | Enrichment | 0.450 g | YPD |
| Scales (S. aurata/D. labrax) | Enrichment | 0.120 g | YPD |
| Skin (S. aurata/D. labrax) | Enrichment | 0.780 g | YPD |
| Byproducts (P. longirostris) | Enrichment | 2 mL | YPD; YNB; MI |
| Direct culture | 1 g | YPD-Agar | |
| P. longirostris hydrolyses | Enrichment | 1 mL | YPD |
| Sea-grass and Algae | Enrichment | 1 g | YPD |
| Direct culture | 4 cm | YPD-Agar |
| Sample | YPD | Min-YPD | YNB |
|---|---|---|---|
| A1 | ++ | + | + |
| A2 | +++ | + | + |
| A3 | +++ | + | + |
| A4 | ++ | + | + |
| Sediment | + | + | − |
| Sea water La Goulette | ++++ | + | + |
| P. oceanica | ++ | + | + |
| Z. marina | ++ | + | + |
| Red Algae | +++ | − | + |
| P. longirostris coproduct | ++ | + | + |
| * Raw Spring | ++ | + | − |
| * Raw Summer | ++ | + | − |
| * Raw Autumn | ++ | + | − |
| * Raw Winter | ++ | + | − |
| Chitin | ++ | + | − |
| Chitosan | − | − | − |
| Hydrolysis of P. longirostris | ++ | − | − |
| Skin (Sparus aurata) | +++ | ++ | + |
| Skin (Dicentrarchus labrax) | +++ | ++ | + |
| Scales (Sparus aurata) | ++++ | ++ | + |
| Scales (Dicentrarchus labrax) | ++++ | ++ | + |
| Gills (Sparus aurata) | ++++ | ++ | + |
| Gills (Dicentrarchus labrax) | ++++ | ++ | + |
| Species | Substrates |
|---|---|
| Cryptococcus curvatus | Seawater (−34 m) |
| Meira nashicola | Seawater (−64 m) |
| Meira sp. | Seawater (−64 m) |
| Rhodotorula mucilaginosa | Seawater (−64 m), Seawater (−0.4 m) |
| Sporobolomyces roseus | Chitin P. longirostris |
| Sporobolomyces aff. Ruberrimus | Chitin P. longirostris |
| Sporobolomyces ruberrimus | Chitin P. longirostris |
| Trichosporon asahii | P.longirostris coproduct |
| Debaryomyces hansenii | Scale of D. labrax |
| Candida parapsiolis | Posidonia, red algae, Zostera, skin of D. labrax |
| Yarrowia lipolytica | Sediment, scale of S. aurata, gills of D. labrax, P. longirostris coproduct |
| Candida tenuis | S. aurata gills and skin |
| Source | Specie | Reference | |
|---|---|---|---|
| Sediment | Pacific Ocean | R. minuta; R. mucilaginosa | [68] |
| South China Sea | Cryptococcus aureus | [57] | |
| Pit of Japan | Dipodascus tetrasporeus | [69] | |
| Veraval, India | Candida sp. | [70] | |
| South East coast of India | Candida albicans, C. tropicalis, D. hansenii, Geotrichum sp., Pichia capsulata, Pichia fermentans, Pichia salicaria, | [71] | |
| South East coast of India | R. minuta C. dimennae Y. lipolytica | [72] | |
| Tamil Nadu, India | Cyberlindnera saturnus | [59] | |
| Mangrove of Makran, Gwadar, Pakistan | Candida parapsilosis, Debaryomyces hansenii, Debaryomyces fabryii, Saccharomyces cerevisiae, Saccharomyces bayanus Shizosaccharomyces pombe | [8] | |
| Seawater | Northwest Pacific Ocean | R. pacifica | [16] |
| Mid-Atlantic ridge (−2300 m) | Candida sp | [72] | |
| Coastal waters of northeast Taiwan | Candida tropicalis Pichia anomala Issatchenkia orientalis C. glabrata Saccharomyces vakushimaensis Kodamaea ohmeri Hanseniaspora uvarum Kazachstania jiainicus Torulaspora delbrueckii | [34] | |
| Veraval, India | Candida sp. | [70] | |
| Queens Cliff, Victoria region, Australia | Rhodotorula sp | [26] | |
| Medit. Sea. Alexandria, Egypt | Candida viswanathii | [73] | |
| Red Sea. Ismailia, Egypt | Candida tropicalis | ||
| Irish Sea. | Candida tropicalis | ||
| Wales, U.K. | Candida tropicalis | ||
| English Channel, Plymouth, U.K. | Saccharomyces cerevisiae Wickerhamomyces anomalus Pichia kudriavzevii Candida glabrata | ||
| Fisheries coproduct | Gastropod gills Ifremeria nautilei | Debaryomyces hansenii | [38] |
| Shrimp and Mussel | R. mucilaginosa | [38] | |
| Fish intestine, Chili | R. mucilaginosa Debaryomyces hansenii | [74] | |
| Abramis brama, Rutilus, Perca fluviatilis | Cr. uniguttulatus; R. mucilaginosa; R. glutinis | [61] | |
| Marine plant | Marine Plant, Chili | Rhodotorula mucilaginosa | [75] |
| Antarctic Algae | R. mucilaginosa | [76] | |
| RmTun15 | YlTun15 | TaTun15 | CtTun15 | DhTun2015 | ||
|---|---|---|---|---|---|---|
| mg/100 g (dry biomass) | ||||||
| Mineral | Calcium | 1.15 | 1.9 | 1.75 | 2.62 | 1.82 |
| Magnesium | 0.823 | 0.896 | 0.637 | 1.92 | 0.573 | |
| Potassium | 8.68 | 11.7 | 7.03 | 18.9 | 5.8 | |
| Sodium | 0.12 | 0.05 | 0.116 | 0.15 | 0.0948 | |
| Iron | 0.0884 | 0.0713 | 0.0523 | 0.111 | 0.0656 | |
| Manganese | 0.0061 | 0.00547 | 0.00259 | 0.00668 | 0.00245 | |
| Nickel | 0.00164 | 0.00328 | 0.00243 | |||
| Selenium | 0.00446 | 0.00453 | 0.00113 | 0.00065 | 0.00046 | |
| Biogenic Amine | Putrescine | 12 | 15 | - | 1 | 2 |
| Cadaverine | 37 | 94 | 6 | 23 | 18 | |
| Histamine | 14 | 13 | 9 | 12 | 8 | |
| Tyramine | 56 | 20 | 20 | 30 | 13 | |
| Spermidine | 84 | 61 | 12 | 44 | 22 | |
| BAI | 0.74 | 1.97 | 1.15 | 0.80 | 1.21 | |
| g/100 g (dry mass) | ||||||
| Ash | 2.43 ± 0.01 | 2.56 ± 0.06 | 1.82 ± 0.10 | 2.36 ± 0.35 | 3.01 ± 0.14 | |
| Carbohydrate | 25.19 ± 0.27 m | 23.21 ± 0.74 n | 33.12 ± 0.35 | 23.21 ± 0.51 n | 26.19 ± 0.58 m | |
| Lipid | 26.63 ±0.19 1 | 37.57 ± 0.41 2 | 11.15 ± 0.42 3 | 15.25 ± 0.244 | 14.33 ± 0.35 5 | |
| Protein | 28.67 ± 0.11 S (33%) | 24.37 ± 0.20 T(26%) | 30.76 ± 0.03 (35%) | 30.50 ± 0.09 (40%) | 32.12 ± 0.28 V (40%) | |
| Fatty Acid | C14:0 | - | 0.50 ± 0.04 | 0.48 ± 0.05 | 0.03 | 0.04 |
| C15:0 | 7.34 ± 1.44 | 6.88 ± 0.45 | 2.16 | 4.14 ± 0.26 | 6.78 ± 1.04 | |
| C16:0 | 1.21 ± 0.41 | 12.11 ± 0.43 | 3.56 ± 0.52 | 0.26 | 0.68 ± 0.06 | |
| C16:1 W7 | 0.83 ± 0.03 | 0.05 ± 0.006 | - | 0.26 | 0.07 | |
| C16:2 W4 | 7.94 ± 1.03 | 2.21 ± 0.44 | 0.34 ± 0.06 | 2.37 ± 0.20 | 1.31 ± 0.07 | |
| C16:3 W4 | - | - | - | 0.04 | 0.15 | |
| C18:0 | 1.75 ±0.18 | 7.56 ± 0.21 | 1.42 ± 0.07 | 0.57 ± 0.11 | 1.97 ± 0.67 | |
| C18:1 W9 | 0.24 ± 0.07 | 0.21 ± 0.006 | 0.29 ± 0.02 | 0.47 ± 0.21 | 2.13 ± 0.03 | |
| C18:1 W7 | - | 0.15 ± 0.004 | - | - | - | |
| C18:2 W6 | 2.18 ± 0.13 | 1.41 ± 0.41 | 2.12 ± 0.61 | 2.33 ± 0.95 | 0.84 ± 0.09 | |
| C18:3 W4 | 0.75 | 1.41 ± 0.03 | 0.25 ± 0.04 | - | - | |
| C18:3 W3 | - | - | - | 0.27 ± 0.11 | 0.24 ± 0. 29 | |
| C18:4 W3 | - | - | - | - | 0.07 ±0.017 | |
| C20:1 W9 | - | 0.18 ± 0.02 | - | - | - | |
| C20:4 W6 | - | - | - | 0.13 ± 0.02 | - | |
| C20:4 W3 | - | - | - | - | - | |
| C20:5 W3 | - | - | - | - | - | |
| C22:5 W3 | 0.47± 0.04 | 0.50 ± 0.04 | - | - | ||
| C22:6 W3 | 0.76 ± 0.07 | 1.09 ± 0.14 | 0.34 ± 0.01 | - | 0.03 ± 0.003 | |
| SFA | 10.30 ± 0.52 b | 27.04± 0.28 a | 7.61 ± 1.35 | 5 ± 0.62 | 9.47 ± 1.46 b | |
| MUFA | 1.07 α | 0.58 ± 0.05 β | 0.29 ± 0.03 γ | 0.73 ± 0.08 λ | 2.20 ± 0.78 Ω | |
| PUFA | 12.10 ± 0.07 ** | 6.61 ± 0.10 * | 3.06 ± 0.58 | 5.14 ± 1.41 * | 2.64 ± 0.15 | |
| ∑ FA | 23.47 ± 0.02 A | 34.24 ± 0.13 B | 10.95 ±0.78 | 10.86 ± 1.37 | 14.32 ± 0.77 C | |
| Amino Acid | Aspartate | 1.41 ± 0.11 | 1.90 ± 0.36 | 2.16 ± 0.12 | 8.67 ± 0.02 | 1.91 ± 0.04 |
| Glutamate | 1.56 ± 0.41 | 0.92 ± 0.01 | 0.98 ± 0.02 | 1.63 ± 0.69 | 1.86 ± 0.37 | |
| Serine | 2.05 ± 0.08 | 1.73 ± 0.03 | 1.32 ± 0.03 | 1.23 ± 0 | 1.88 ± 0.02 | |
| Asparagine | 0.14 ± 0.00 | - | 0.14 | - | 0.14 | |
| Glutamine | 0.20 ± 0.00 | 0.2 | 0.2 | - | 0.2 | |
| Histidine | 0.58 ± 0.01 | 0.72 ± 0.1 | 0.48 ± 0.05 | 0.81 | 0.50 ± 0.01 | |
| Glycine | 1.48 ± 0.02 | 1.73 | 1.22 ± 0.01 | 1.04 | 1.23 ± 0.03 | |
| Threonine | 1.37 ± 0.02 | 1.72 ± 0.13 | 1.24 ± 0.16 | 1.11 | 1.49 ± 0.13 | |
| Arginine | 1.84 ± 0.09 | 0.94 | 1.15 ± 0.05 | 0.68 | 1.67 ± 0.10 | |
| Alanine | 1.40 ± 0.27 | 1.09 ± 0.25 | 2.87 ± 0.14 | 1.67 ± 0.07 | 1.82 ± 0.04 | |
| Tyrosine | 0.87 ± 0.05 | 0.76 ± 0.03 | 0.62 ± 0.02 | 0.51 | 0.73 | |
| Valine | 0.45 ± 0.02 | 2.31 ± 0.16 | 0.26 ± 0.16 | 0.18 | 0.23 ± 0.17 | |
| Methionine | 2.17 ± 0.04 | 0.53 ± 0.05 | 1.08 ± 0.05 | 0.61 ± 0.05 | 0.28 ± 0.02 | |
| Tryptophan | 0.10 ± 0.02 | 0.10 ± 0.04 | 0.08 ± 0.03 | 0.8 | 3.48 ± 0.14 | |
| Phenylalanine | 0.97 ± 0.06 | 0.98 ± 0.09 | 0.78 ± 0.06 | 1.35 | 1.07 ± 0.03 | |
| Isoleucine | 1.01 ± 0.01 | 1.06 ± 0.08 | 0.86 ± 0.03 | 0.14 | 1.12 ± 0.02 | |
| Leucine | 2.21 ± 0.14 | 1.64 ± 0.13 | 1.74 ± 0.10 | 1.29 | 1.82 ± 0.02 | |
| Lysine | 2.55 ± 0.35 | 2.54 ± 0.04 | 1.94 ± 0.03 | 2.54 ± 0.03 | 2.59 ± 0.01 | |
| Hydroxyproline | 0.71 ± 0.35 | 2.05 ± 0.49 | 2.42 ± 0.09 | 1.29 ± 0.02 | 2.85 ± 0.59 | |
| Proline | 2.11 ± 0.02 | 2.94 ± 0.13 | 1.71 ± 0.14 | 1.69 ± 0.44 | 2.57 ± 0.27 | |
| ∑ AA | 25.18 ± 0.10 R | 23.86 ± 0.11 | 23.25 ± 0.06 | 27.24 ± 0.06 S | 29.44 ± 0.10 T | |
| EAA | 15.26 ± 0.07 (61%) | 15.38 ± 0.09 (64%) | 11.24 ± 0.08 X (48%) | 10.4 ± 0.05 Y (38%) | 13.34 ± 0.07 Z (45%) | |
| NEAA | 9.92 ± 0.13 (39%) | 8.48 ± 0.12 (36%) | 12.01 ± 0.04 (52%) | 16.84 ± 0.08 (62%) | 16.1 ± 0.12 (55%) | |
| Application | Species | Reference |
|---|---|---|
| Biodiesel (SCO) | Trichosporon fermentans | [119] |
| Rhodotorula glutines | [120] | |
| Rhodotorula mucilaginosa | [121] | |
| Rhodotorula graminis | [122] | |
| Rhodotorula toruloides NBRC 0559 | [123] | |
| Yarrowia lipolytica | [124] | |
| Debaryomyces etchellssi | [125] | |
| Trichosporon sp. | [126] | |
| Trichosporon cutaneum | [127] | |
| Yarrowia lipolytica NCIM3589 | [128,129] | |
| Rhodotorula kartochvitovae SY89 | [130] | |
| Rhodotorula glutinis | [131] | |
| Yarrowia lipolytica | [132] | |
| Y. lipolytica YlTun15; MY-2 et MY-3 | [31] | |
| R. mucilaginosa RmTun15; MR-2 et MR-3 | ||
| T. asahii TaTun15; MT-2 et MT-2 | ||
| Bioethanol | Rhodotorula minuta Yarrowia lipolytica | [133] |
| Candida albicans Candida tropicalis Debaryomyces hansenii | [71] | |
| Candida sp. | [70] | |
| Yarrowia lipolytica Po1g | [134] | |
| Yarrowia lipolytica | [135] | |
| Yarrowia lipolytica | [136] | |
| Yarrowia lipolytica | [12] | |
| Yarrowia lipolytica | [137] | |
| Yarrowia lipolytica | [138] | |
| Debaryomyces hansenii | [139] | |
| Debaryomyces hansenii CBS004 | [140] | |
| Trichosporon asahii | [141] | |
| Y. lipolytica RmTun15 | [28] | |
| Rhodotorula mucilaginosa | [142] | |
| Carotenoids (Torulene, torularhodin,β-carotene) | Rhodotorula mucilaginosa | [143] |
| Rhodotorula RY1801 | [144] | |
| Rhodotorula mucilaginosa URM7409 | [145] | |
| Rhodotorula glutinis | [146] | |
| R. mucilaginosa ATCC 66034 R. gracilis ATCC R. glutinis LOCKR13 | [147] | |
| Rhodotorula mucilaginosa C2.5t1 | [148] | |
| Rhodotorula sp | [149] | |
| Rhodotorula glutinis CCY 20-2-26 | [150] | |
| Yarrowia lipolytica W29 | [151] | |
| Rhodotorula glutinis | [152] | |
| Rhodotorula mucilaginosa AY-01 | [153] | |
| Yarrowia lipolytica | [154] | |
| Rhodotorula mucilaginosa | [155] | |
| Rhodotorula mucilaginosa | [156] | |
| Rhodotorula glutinis DM28 | [157] | |
| Rhodotorula glutinis | [158] | |
| Pharmaceutical | Debaryomyces hansenii | [159] |
| Yarrowia lipolytica | [160] | |
| Yarrowia lipolytica NCYC 789 | [161] | |
| Candida tropicalis | [162] | |
| Nanoparticles | Candida albicans, C. tropicalis, Debaryomyces hansenii, Rhodotorula minuta, Yarrowia lipolytica | [163] |
| Rhodotorula sp ATL72 | [164] | |
| Yarrowia lipolytica IMUFRJ 50682 | [165] | |
| Alimentation | Candida sp. | [35] |
| Rhodotorula mucilaginosa | [166] | |
| Rhodotorula sp. | [80] | |
| Debaryomyces hansenii S8 Debaryomyces hansenii S100 Candida tropicalis S186 | [79,87] | |
| Debaryomyces hansenii | [167] | |
| Debaryomyces hansenii | [168] | |
| Debaryomyces hansenii | [169] | |
| Yarrowia lipolytica | [170] | |
| Rhodotorula sp. H26 | [171] | |
| Bio-surfactant | Yarrowia lipolytica NCIM 3589 | [172] |
| Yarrowia lipolytica IMUFRJ50682 | [173] | |
| Candida glabrata | [174] | |
| Candida tropicalis | [175] | |
| Candida tropicalis | [176] | |
| Rhodotorula mucilaginosa KUGPP-1 | [177] | |
| Candida lipolytica | [178] | |
| Candida bombicola | [179] | |
| Candida tropicalis UCP 1613 | [180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessadok, B.; Jaouadi, B.; Brück, T.; Santulli, A.; Messina, C.M.; Sadok, S. Molecular Identification and Biochemical Characterization of Novel Marine Yeast Strains with Potential Application in Industrial Biotechnology. Fermentation 2022, 8, 538. https://doi.org/10.3390/fermentation8100538
Bessadok B, Jaouadi B, Brück T, Santulli A, Messina CM, Sadok S. Molecular Identification and Biochemical Characterization of Novel Marine Yeast Strains with Potential Application in Industrial Biotechnology. Fermentation. 2022; 8(10):538. https://doi.org/10.3390/fermentation8100538
Chicago/Turabian StyleBessadok, Boutheina, Bassem Jaouadi, Thomas Brück, Andrea Santulli, Concetta Maria Messina, and Saloua Sadok. 2022. "Molecular Identification and Biochemical Characterization of Novel Marine Yeast Strains with Potential Application in Industrial Biotechnology" Fermentation 8, no. 10: 538. https://doi.org/10.3390/fermentation8100538