
Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus Co-Inoculation on Alcoholic Fermentation Behavior and Aromatic Profile of Sauvignon Blanc Wine
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Conditions
2.2. Must Preparation and Fermentation Kinetics
2.3. Microbiological Analyses and Yeast Molecular Identification
2.4. Chemical Analyses
2.4.1. Analysis of Classical Oenological Parameters
2.4.2. Higher Alcohols, Acetates and Esters Quantification
2.4.3. Varietal Thiols Quantification
2.5. Sensory Analyses
2.6. Statistical Analyses
3. Results
3.1. Yeast Growth and Ooenological Parameters during Fermentation
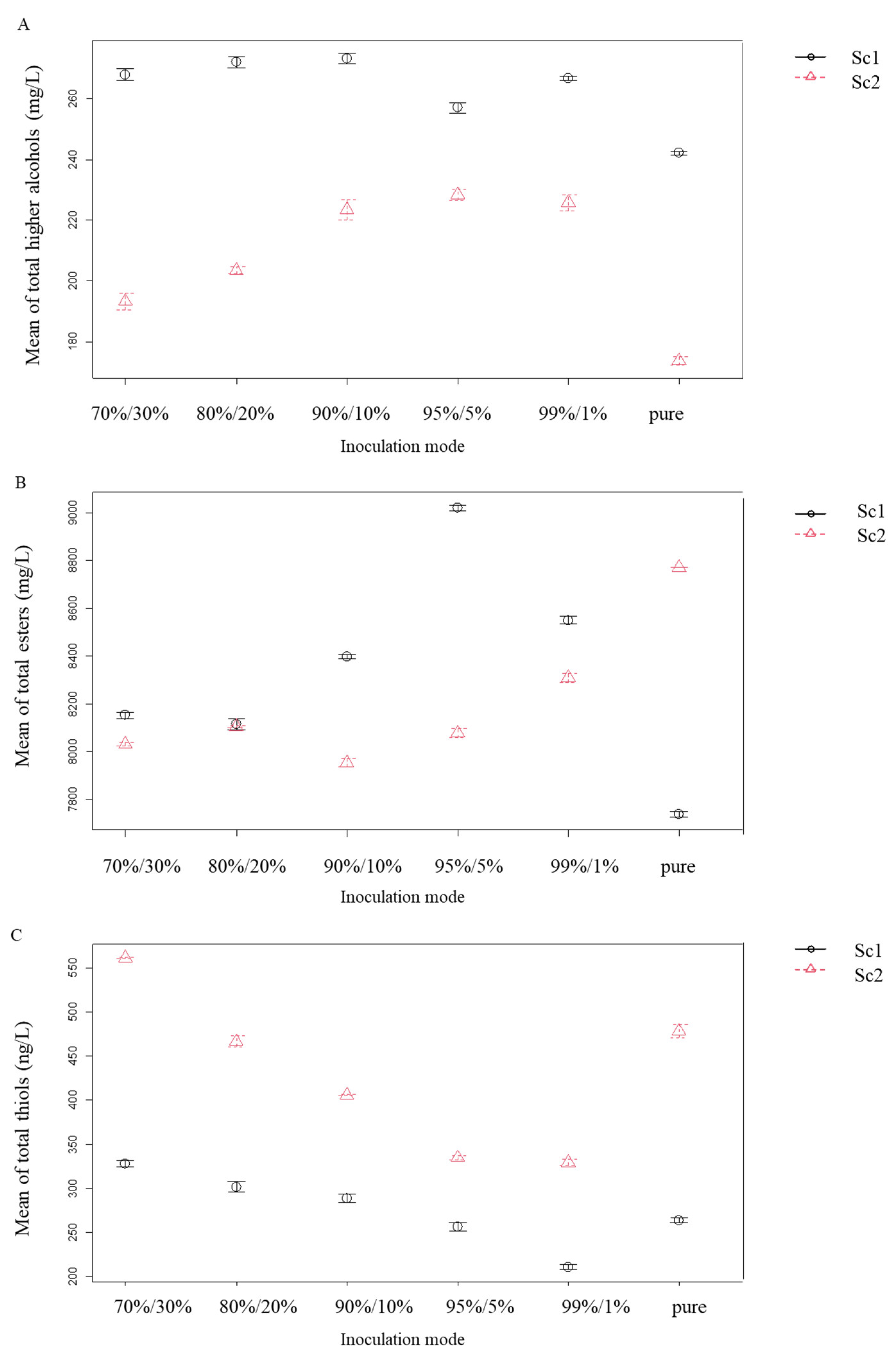
3.2. Volatile Compounds Analyses
3.3. Sensory Impact
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the enzymatic basis of wine “flavorome”: A phylo-functional study of wine related yeast species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, M.; Mannazzu, I.; Marinangeli, P.; Clementi, F.; Martini, A. Contribution of winery-resident Saccharomyces cerevisiae strains to spontaneous grape must fermentation. Antonie Van Leeuwenhoek 2004, 85, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast interactions and molecular mechanisms in wine fermentation: A comprehensive review. Int. J. Mol. Sci. 2021, 22, 7754. [Google Scholar] [CrossRef] [PubMed]
- Brice, C.; Cubillos, F.A.; Dequin, S.; Camarasa, C.; Martínez, C. Adaptability of the Saccharomyces cerevisiae yeasts to wine fermentation conditions relies on their strong ability to consume nitrogen. PLoS ONE 2018, 13, e0192383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertin, W.; Masneuf-Pomarede, I.; Galeote, V.; Legras, J.-L. New insights into wine yeast diversities. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 117–163. [Google Scholar]
- Dimopoulou, M.; Troianou, V.; Toumpeki, C.; Gosselin, Y.; Dorignac, É.; Kotseridis, Y. Effect of strains from different Saccharomyces species used in different inoculation schemes on chemical composition and sensory characteristics of Sauvignon blanc wine. OENO One 2020, 54, 745–759. [Google Scholar] [CrossRef]
- Darriet, P.; Tominaga, T.; Lavigne, V.; Boidron, J.-N.; Dubourdieu, D. Identification of a powerful aromatic component of Vitis vinifera L. var. Sauvignon wines: 4-mercapto-4-methylpentan-2-one. Flavour Fragr. J. 1995, 10, 385–392. [Google Scholar] [CrossRef]
- Tominaga, T.; Furrer, A.; Henry, R.; Dubourdieu, D. Identification of new volatile thiols in the aroma of Vitis vinifera L. var. Sauvignon blanc wines. Flavour Fragr. J. 1998, 13, 159–162. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A.; Francis, I.L.; Pretorius, I.S. Engineering volatile thiol release in Saccharomyces cerevisiae for improved wine aroma. Yeast 2007, 24, 561–574. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Pollon, M.; Fracassetti, D.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Volatile profile of white wines fermented with sequential inoculation of Starmerella bacillaris and Saccharomyces cerevisiae. Food Chem. 2018, 257, 350–360. [Google Scholar] [CrossRef]
- Masneuf-Pomarède, I.; Mansour, C.; Murat, M.-L.; Tominaga, T.; Dubourdieu, D. Influence of fermentation temperature on volatile thiols concentrations in Sauvignon blanc wines. Int. J. Food Microbiol. 2006, 108, 385–390. [Google Scholar] [PubMed]
- Legras, J.-L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [Green Version]
- International Organisation of Vine and Wine. Compendium of International Methods of Wine and Must Analysis; OIV: Paris, France, 2020. [Google Scholar]
- Cozzolino, D.; Parker, M.; Dambergs, R.G.; Herderich, M.; Gishen, M. Chemometrics and visible-near infrared spectroscopic monitoring of red wine fermentation in a pilot scale. Biotechnol. Bioeng. 2006, 95, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Heymann, H.; Ebeler, S.E. Sensory and Instrumental Evaluation of Alcoholic Beverages; Elsevier Inc.: London, UK, 2017. [Google Scholar]
- Nanou, E.; Mavridou, E.; Milienos, F.S.; Papadopoulos, G.; Tempère, S.; Kotseridis, Y. Odor characterization of white wines produced from indigenous Greek grape varieties using the frequency of attribute citation method with trained assessors. Foods 2020, 9, 1396. [Google Scholar] [CrossRef] [PubMed]
- Dubourdieu, D.; Tominaga, T. Polyfunctional thiol compounds. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 275–293. [Google Scholar]
- Goméz, M.J.; Cacho, J.F.; Ferreira, V.; Vicario, I.M.; Heredia, F.J. Volatile components of Zalema white wines. Food Chem. 2007, 100, 1464–1473. [Google Scholar] [CrossRef]
- Li, H. Wine Tasting; Science Press: Beijing, China, 2006; pp. 29–106. [Google Scholar]
- Goddard, M.R. Quantifying the complexities of Saccharomyces cerevisiae’s ecosystem engineering via fermentation. Ecology 2008, 89, 2077–2082. [Google Scholar] [CrossRef] [Green Version]
- Tempère, S.; Marchal, A.; Barbe, J.-C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma—A review. South Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces commercial starter cultures: Scientific trends, recent patents and innovation in the wine sector. Recent Pat. Food Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Wang, C.; Mas, A.; Esteve-Zarzoso, B. The interaction between Saccharomyces cerevisiae and non-Saccharomyces yeast during alcoholic fermentation is species and strain specific. Front. Microbiol. 2016, 7, 502. [Google Scholar] [CrossRef]
- Franco, W.; Benavides, S.; Valencia, P.; Ramírez, C.; Urtubia, A. Native yeasts and lactic acid bacteria isolated from spontaneous fermentation of seven grape cultivars from the Maule region (Chile). Foods 2021, 10, 1737. [Google Scholar] [CrossRef] [PubMed]
- Bordet, F.; Joran, A.; Klein, G.; Roullier-Gall, C.; Alexandre, H. Yeast–yeast interactions: Mechanisms, methodologies and impact on composition. Microorganisms 2020, 8, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Ždralević, M.; Marra, E. Molecular mechanisms of Saccharomyces cerevisiae stress adaptation and programmed cell death in response to acetic acid. Front. Microbiol. 2013, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Reduction of volatile acidity of wines by selected yeast strains. Appl. Microbiol. Biotechnol. 2008, 80, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The influence of non-Saccharomyces species on wine fermentation quality parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Renault, P.E.; Albertin, W.; Bely, M. An innovative tool reveals interaction mechanisms among yeast populations under oenological conditions. Appl. Microbiol. Biotechnol. 2013, 97, 4105–4119. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Fragasso, M.; Pico, J.; Panighel, A.; Castellarin, S.D.; Flamini, R.; Grieco, F. Influence of non-Saccharomyces on wine chemistry: A focus on aroma-related compounds. Molecules 2021, 26, 644. [Google Scholar] [CrossRef]
- Bordet, F.; Roullier-Gall, C.; Ballester, J.; Vichi, S.; Quintanilla Casas, B.; Gougeon, R.; Julien-Ortiz, A.; Schmitt-Kopplin, P.; Alexandre, H. Different wines from different yeasts? Saccharomyces cerevisiae intraspecies differentiation by metabolomic signature and sensory patterns in wine. Microorganisms 2021, 9, 2327. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41 (Suppl. S1), S95–S128. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.-J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast–yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; Bordet, F.; David, V.; Schmitt-Kopplin, P.; Alexandre, H. Yeast interaction on Chardonnay wine composition: Impact of strain and inoculation time. Food Chem. 2022, 374, 131732. [Google Scholar] [CrossRef] [PubMed]
- Cameleyre, M.; Lytra, G.; Barbe, J.-C. Static headspace analysis using low-pressure gas chromatography and mass spectrometry, application to determining multiple partition coefficients: A practical tool for understanding red wine fruity volatile perception and the sensory impact of higher alcohols. Anal. Chem. 2018, 90, 10812–10818. [Google Scholar]
- Dimopoulou, M.; Raffenne, J.; Claisse, O.; Miot-Sertier, C.; Iturmendi, N.; Moine, V.; Coulon, J.; Dols-Lafargue, M. Oenococcus oeni exopolysaccharide biosynthesis, a tool to improve malolactic starter performance. Front. Microbiol. 2018, 9, 1276. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Gardner, R.C. Yeast genes required for conversion of grape precursors to varietal thiols in wine. FEMS Yeast Res. 2015, 15, fov034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiegers, J.H.; Willmott, R.; Hill-Ling, A.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Howell, K.S.; de Barros Lopes, M.A.; Sefton, M.A.; Lilly, M.; et al. Modulation of volatile thiol and ester aromas by modified wine yeast. Dev. Food Sci. 2006, 43, 113–116. [Google Scholar]
- Sarrazin, E.; Shinkaruk, S.; Pons, M.; Thibon, C.; Bennetau, B.; Darriet, P. Elucidation of the 1,3-sulfanylalcohol oxidation mechanism: An unusual identification of the disulfide of 3-sulfanylhexanol in sauternes botrytized wines. J. Agric. Food Chem. 2010, 58, 10606–10613. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strains in Pure Cultures | |
|---|---|
| Code | Yeast strains |
| Sp2 | S. pastorianus Sp2 |
| Sc1 | S. cerevisiae CK S102 |
| Sc2 | S. cerevisiae SH 12 |
| Yeast strains in mixed cultures | |
| Code | Yeast strains |
| Sp2 70%–Sc1 30% | S. pastorianus Sp2 70%–S. cerevisiae CK S102 30% (w/w) |
| Sp2 80%–Sc1 20% | S. pastorianus Sp2 80%–S. cerevisiae CK S102 20% (w/w) |
| Sp2 90%–Sc1 10% | S. pastorianus Sp2 90%–S. cerevisiae CK S102 10% (w/w) |
| Sp2 95%–Sc1 5% | S. pastorianus Sp2 95%–S. cerevisiae CK S102 5% (w/w) |
| Sp2 99%–Sc1 1% | S. pastorianus Sp2 99%–S. cerevisiae CK S102 1% (w/w) |
| Sp2 70%–Sc2 30% | S. pastorianus Sp2 70%–S. cerevisiae SH 12 30% (w/w) |
| Sp2 80%–Sc2 20% | S. pastorianus Sp2 80%–S. cerevisiae SH 12 20% (w/w) |
| Sp2 90%–Sc2 10% | S. pastorianus Sp2 90%–S. cerevisiae SH 12 10% (w/w) |
| Sp2 95%–Sc2 5% | S. pastorianus Sp2 95%–S. cerevisiae SH 12 5% (w/w) |
| Sp2 99%–Sc2 1% | S. pastorianus Sp2 99%–S. cerevisiae SH 12 1% (w/w) |
| Aromatic Compounds | Pure Cultures | Sp2/Sc1 Mixed Cultures | Sp2/Sc2 Mixed Cultures | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sp2 | Sc1 | Sc2 | Sp2 70% Sc1 30% | Sp2 80% Sc1 20% | Sp2 90% Sc1 10% | Sp2 95% Sc1 5% | Sp2 99% Sc1 1% | Sp2 70% Sc2 30% | Sp2 80% Sc2 20% | Sp2 90% Sc2 10% | Sp2 95% Sc2 5% | Sp2 99% Sc2 1% | |
| Thiols (ng/L) | |||||||||||||
| 3-mercaptohexan-1-ol | 282.5 ± 15 f | 249.7 ± 3 g | 385.7 ± 15 b | 315.3 ± 7 c,d | 290.7 ± 9 d,e,f | 283.3 ± 7 e,f | 251.0 ± 6 g | 205.7 ± 3 h | 424.0 ± 5 a | 373.7 ± 7 b | 317.7 ± 5 c | 308.7 ± 2 c,d,e | 317.3 ± 4 c,d |
| 3-mercaptohexan-1-ol Acetate | 5.8 ± 0.6 f | 10.0 ± 1.0 b,c | 19.3 ± 1.4 a | 8.0 ± 1.0 c,d,e | 6.0 ± 1.0 e,f | ND g | ND g | ND g | 11.0 ± 1.0 b | 8.7 ± 1.5 b,c,d | 6.7 ± 0.6 d,e,f | 5.0 ± 0.0 f | ND g |
| 4-methyl-4-mercaptopentan-2-one | 5.2 ± 1.2 f | 4.3 ± 0.6 f | 73.3 ± 3.5 c | 4.6 ± 0.6 f | 5.0 ± 1.0 f | 5.3 ± 0.6 f | 5.3 ± 1.5 f | 5.3 ± 1.5 f | 126.0 ± 3 a | 84.0 ± 3.0 b | 81.3 ± 3.5 b | 21.0 ± 2.0 d | 12.3 ± 1.5 e |
| Total thiols | 293.5 ± 16 f | 264.0 ± 4 g,h | 478.3 ± 13 b | 328.0 ± 6 d,e | 301.6 ± 11 e,f | 288.6 ± 8 f,g | 256.3 ± 8 h | 211.0 ± 4 i | 561.0 ± 2 a | 466.3 ± 10 b | 405.7 ± 1 c | 334.7 ± 4 d | 329.7 ± 5 d,e |
| Higher alcohols (mg/L) | |||||||||||||
| 2- phenyl-ethanol | 18.8 ± 0.7 d,e,f | 28.4 ± 0.9 a,b | 13.9 ± 1.1 g | 28.6 ± 0.6 a,b | 30.1 ± 0.9 a | 24.9 ± 4.0 b,c | 20.9 ± 0.9 c,d | 19.8 ± 0.8 d,e | 14.5 ± 0.9 f,g | 13.9 ± 1.1 g | 15.5 ± 1.1 e,f,g | 17.0 ± 1 d,e,f,g | 16.5 ± 1.0 d,e,f,g |
| propanol | 28.3 ± 1.8 a,b,c | 20.3 ± 0.7 h | 23.9 ± 1.8 e,f,g | 20.6 ± 0.6 g,h | 25.0 ± 1.0 d,e,f | 26.7 ± 1.3 c,d,e | 22.2 ± 0.7 f,g,h | 28.0 ± 0.8 a,b,c,d | 25.5 ± 1.1 d,e | 30.4 ± 0.6 a | 30.4 ± 1.2 a | 27.0 ± 1.0 b,c,d,e | 30.1 ± 0.9 a,b |
| isobutanol | 31.3 ± 1.2 a | 19.0 ± 0.0 g,h | 12.3 ± 0.8 i | 22.7 ± 0.5 e,f | 26.1 ± 0.8 b,c,d | 27.7 ± 1.5 b,c | 22.7 ± 0.6 e,f | 28.2 ± 0.3 b | 17.2 ± 0.9 h | 18.1 ± 0.4 h | 21.3 ± 1.4 f,g | 24.1 ± 0.9 d,e | 25.1 ± 0.9 c,d,e |
| butanol | 1.1 ± 0.1 a | 0.7 ± 0.1 d | 0.9 ± 0.0 b,c,d | 0.8 ± 0.1 c,d | 0.9 ± 0.1 a,b,c | 1.0 ± 0.1 a,b,c | 1.0 ± 0.0 a,b,c | 1.1 ± 0.1 a,b | 0.9 ± 0.2 a,b,c | 1.0 ± 0.1 a,b,c | 1.0 ± 0.0 a,b,c | 1.0 ± 0.1 a,b,c | 1.1 ± 0.1 a,b |
| isoamyl alcohol | 174.4 ± 5 b | 173.8 ± 3 b | 122.7 ± 2 e | 195.2 ± 2 a | 190.0 ± 3 a | 193.1 ± 3 a | 190.3 ± 2 a | 189.7 ± 1.5 a | 135.3 ± 3 d | 140.0 ± 1 d | 157.2 ± 4 c | 159.3 ± 1 c | 153.0 ± 2 c |
| Total alcohols | 254.0 ± 7 c | 242.3 ± 1 d | 173.7 ± 2 f | 239.4 ± 3 a,b,c | 242.0 ± 2 a,b,c | 247.1 ± 2 a | 248.6 ± 3 b,c | 236.2 ± 2 a,b | 247.0 ± 1 e | 178.9 ± 4 e | 189.6 ± 1 d | 207.9 ± 5 d | 211.7 ± 4 d |
| Esters (mg/L) | |||||||||||||
| Isoamyl acetate | 3.1 ± 0.1 g | 3.5 ± 0.2 e | 3.7 ± 0.0 d | 3.9 ± 0.0 c | 4.0 ± 0.0 c | 4.2 ± 0.0 b | 4.4 ± 0.0 a | 3.6 ± 0.0 d,e | 2.9 ± 0.0 h | 3.0 ± 0.0 g,h | 3.1 ± 0.0 f,g | 3.2 ± 0.0 f | 3.2 ± 0.0 f |
| Hexyl acetate | 0.2 ± 0.1 c | 0.2 ± 0.0 b | 0.3 ± 0.0 a | 0.2 ± 0.0 c,d | 0.2 ± 0.0 d,e | 0.2 ± 0.0 c,d | 0.2 ± 0.0 d,e | 0.2 ± 0.0 c,d | 0.2 ± 0.0 b,c | 0.2 ± 0.0 f | 0.2 ± 0.0 c | 0.2 ± 0.0 d,e | 0.2 ± 0.0 e,f |
| 2-phenylethyl acetate | 0.3 ± 0.2 e | 0.4 ± 0.0 a | 0.3 ± 0.0 f | 0.3 ± 0.0 b,c | 0.3 ± 0.0 c,d | 0.3 ± 0.0 b | 0.3 ± 0.0 d,e | 0.3 ± 0.0 d,e | 0.2 ± 0.0 g | 0.2 ± 0.0 g | 0.2 ± 0.0 g | 0.2 ± 0.0 f | 0.2 ± 0.0 f |
| Ethyl isobutyrate | 0.1 ± 0.0 b | 0.1 ± 0.0 a | 0.1 ± 0.0 d | 0.1 ± 0.0 b | 0.1 ± 0.0 b,c | 0.1 ± 0.0 b,c | 0.1 ± 0.0 c | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 0.1 ± 0.0 b,c | 0.1 ± 0.0 b,c | 0.1 ± 0.0 b,c | 0.1 ± 0.0 b |
| Ethyl butyrate | 0.6 ± 0.0 a | 0.4 ± 0.0 f | 0.5 ± 0.0 c,d | 0.4 ± 0.1 e | 0.4 ± 0.0 e | 0.4 ± 0.0 e | 0.5 ± 0.0 c,d | 0.5 ± 0.0 c | 0.5 ± 0.1 b,c | 0.6 ± 0.0 a,b | 0.5 ± 0.0 c,d | 0.5 ± 0.0 d | 0.5 ± 0.0 c,d |
| Ethyl octanoate | 1.6 ± 0.1 c | 1.2 ± 0.1 h | 1.7 ± 0.0 a | 1.3 ± 0.1 g,h | 1.3 ± 0.0 g | 1.2 ± 0.0 h | 1.3 ± 0.0 e,f | 1.4 ± 0.0 d,e | 1.6 ± 0.0 b,c | 1.6 ± 0.0 a,b | 1.5 ± 0.0 d | 1,4 ± 0.0 f,g | 1.5 ± 0.0 d,e |
| Ethyl hexanoate | 2.2 ± 0.2 a | 1.8 ± 0.0 e | 2.1 ± 0.0 a.b,c | 1.7 ± 0.1 e,f | 1.6 ± 0.0 f | 1.6 ± 0.0 e,f | 1.9 ± 0.0 d | 2.1 ± 0.0 a,b | 2.2 ± 0.0 a | 2.1 ± 0.0 a,b | 2.0 ± 0.0 c,d | 2.0 ± 0.0 b,c,d | 2.2 ± 0.0 a |
| Ethyl 3-hydroxy butanoate | 0.4 ± 0.0 b | 0.2 ± 0.0 g | 0.2 ± 0.0 e | 0.2 ± 0.0 g | 0.3 ± 0.0 e,f | 0.3 ± 0.0 e | 0.3 ± 0.0 d | 0.3 ± 0.0 c,d | 0.3 ± 0.0 c | 0.4 ± 0.0 c | 0.4 ± 0.0 b | 0.4 ± 0.0 a | 0.4 ± 0.0 a |
| Total esters | 8.4 ± 0.2 d | 7.7 ± 0.0 h | 8.8 ± 0.0 b | 8.2 ± 0.0 e,f | 8.1 ± 0.1 e,f,g | 8.4 ± 0.0 c,d | 9.0 ± 0.0 a | 8.6 ± 0.0 c | 8.0 ± 0.0 f,g | 8.1 ± 0.0 f,g | 8.0 ± 0.0 g | 8.1 ± 0.0 f,g | 8.3 ± 0.0 d,e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimopoulou, M.; Goulioti, E.; Troianou, V.; Toumpeki, C.; Paramithiotis, S.; Gosselin, Y.; Dorignac, E.; Papadopoulos, G.; Kotseridis, Y. Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus Co-Inoculation on Alcoholic Fermentation Behavior and Aromatic Profile of Sauvignon Blanc Wine. Fermentation 2022, 8, 539. https://doi.org/10.3390/fermentation8100539
Dimopoulou M, Goulioti E, Troianou V, Toumpeki C, Paramithiotis S, Gosselin Y, Dorignac E, Papadopoulos G, Kotseridis Y. Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus Co-Inoculation on Alcoholic Fermentation Behavior and Aromatic Profile of Sauvignon Blanc Wine. Fermentation. 2022; 8(10):539. https://doi.org/10.3390/fermentation8100539
Chicago/Turabian StyleDimopoulou, Maria, Elli Goulioti, Vicky Troianou, Chrisavgi Toumpeki, Spiros Paramithiotis, Yves Gosselin, Etienne Dorignac, Georgios Papadopoulos, and Yorgos Kotseridis. 2022. "Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus Co-Inoculation on Alcoholic Fermentation Behavior and Aromatic Profile of Sauvignon Blanc Wine" Fermentation 8, no. 10: 539. https://doi.org/10.3390/fermentation8100539