
q-PCR Methodology for Monitoring the Thermophilic Hydrogen Producers Enriched from Elephant Dung

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate
2.2. Inoculum and Culture Enrichment
2.3. Bio-Hydrogen Fermentation from Sawdust
2.4. Analytical Methods
2.5. PCR-DGGE Analysis
2.6. q-PCR Analysis
3. Results and Discussion
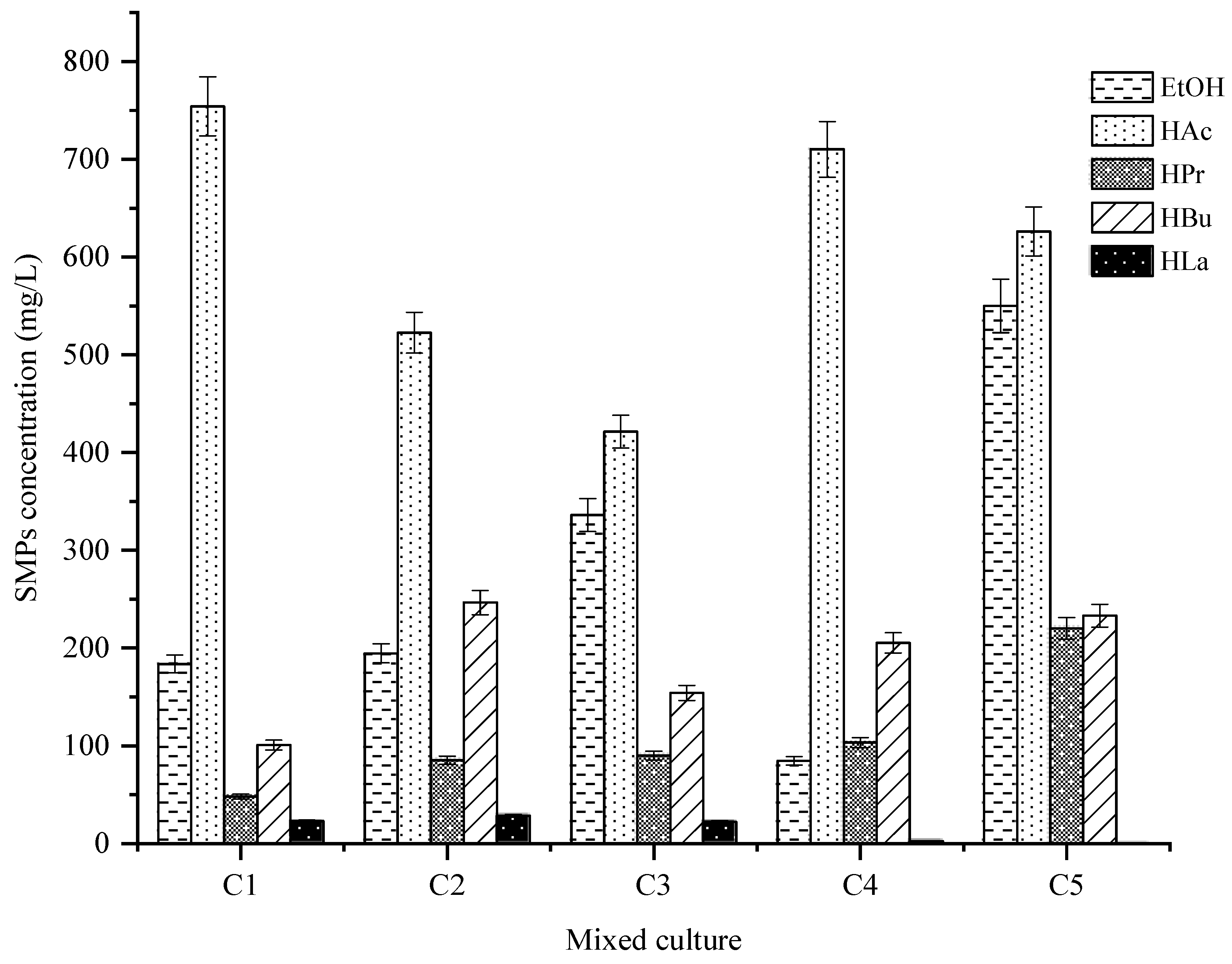
3.1. Enrichment of Thermophilic Hydrogen-Producing Bacteria
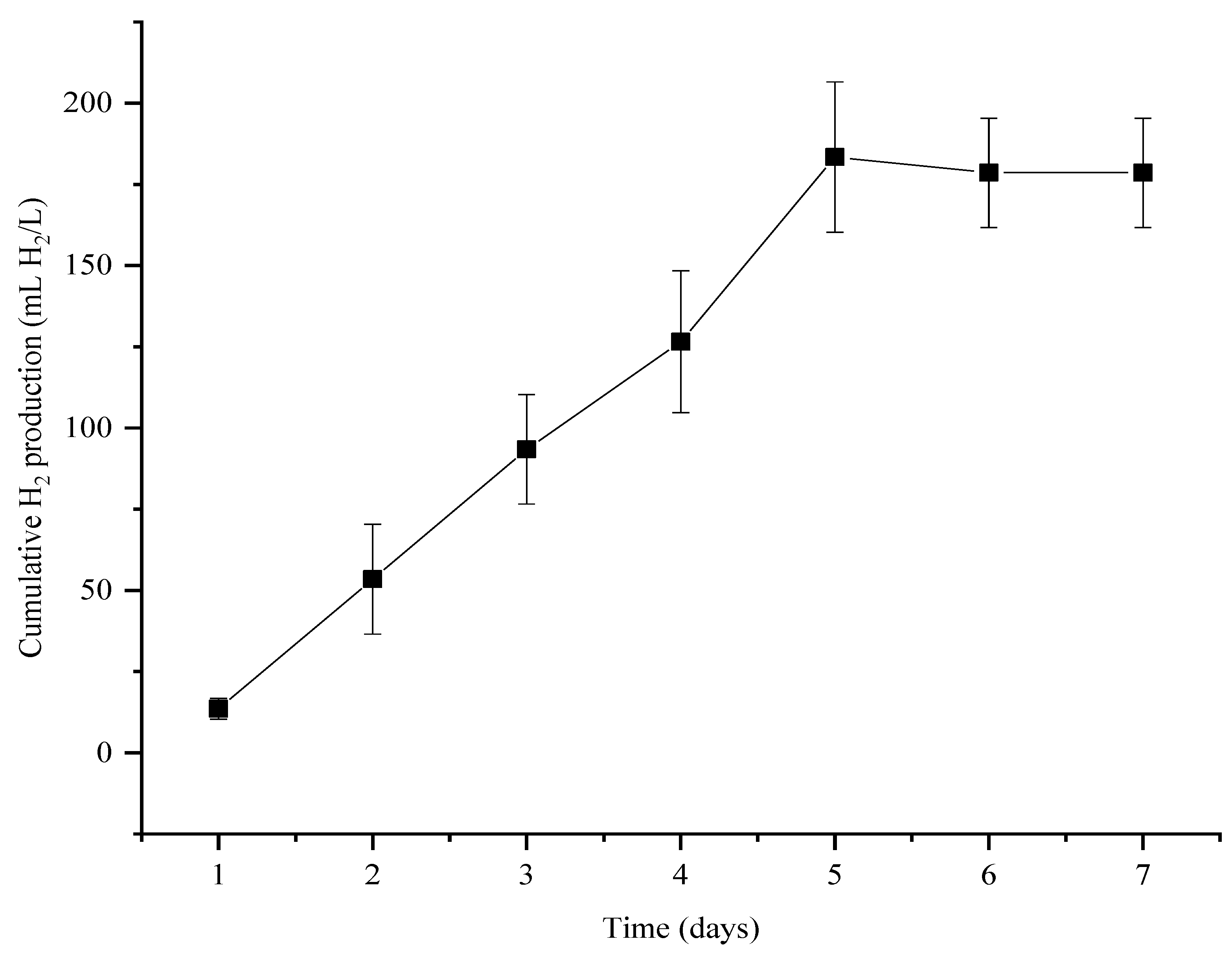
3.2. Thermophilic Bio-Hydrogen Production from Sawdust
3.3. Microbial Community Structure
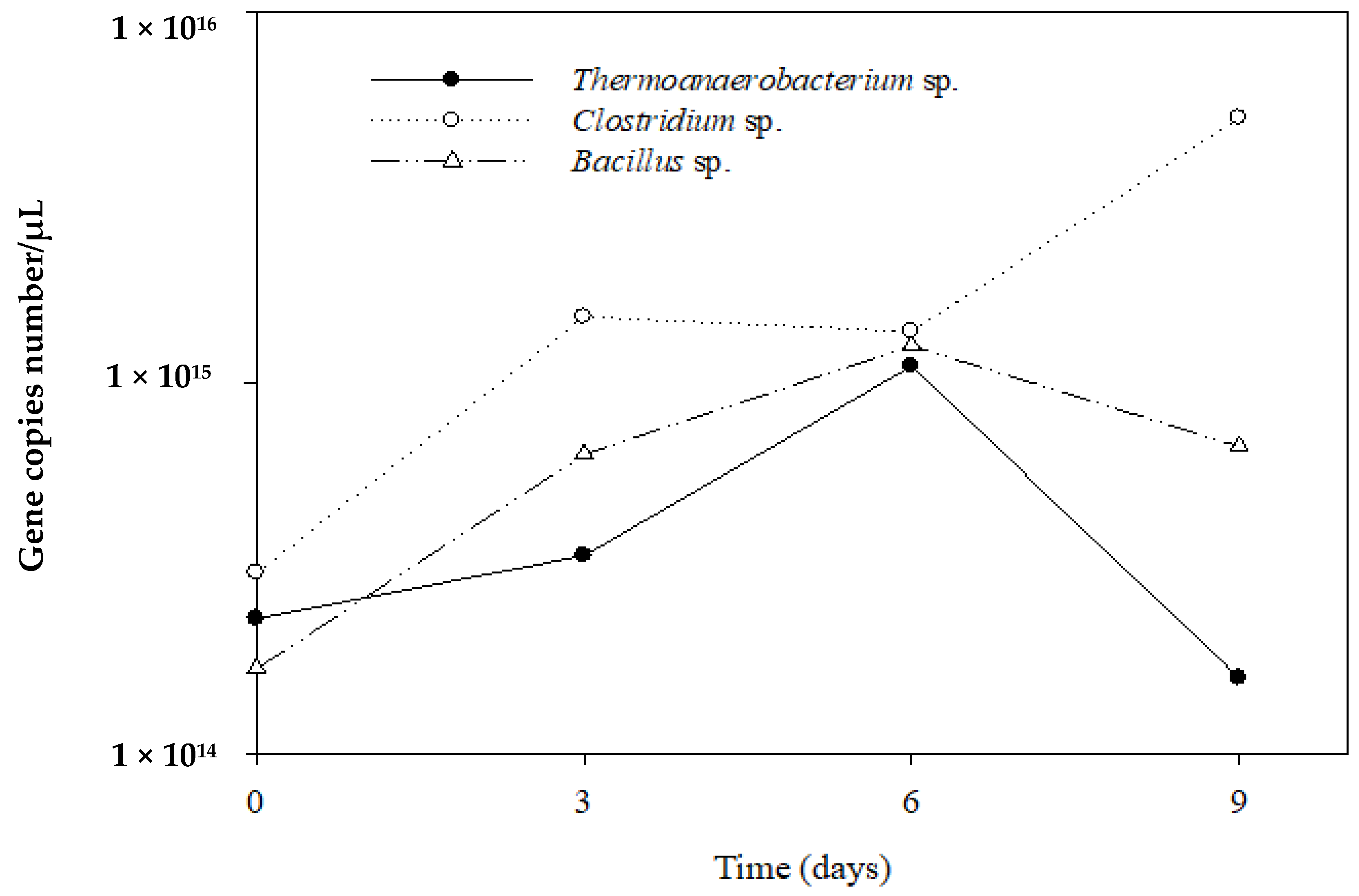
3.4. q-PCR
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, M.; Elreedy, A.; Ibrahim, M.G.; Fujii, M.; Tawfik, A. Hydrogen and methane bio-production and microbial community dynamics in a multi-phase anaerobic reactor treating saline industrial wastewater. Energy Convers. Manag. 2019, 186, 1–14. [Google Scholar] [CrossRef]
- Gomez-Flores, M.; Nakhla, G.; Hafez, H. Hydrogen production and microbial kinetics of Clostridium termitidis in mono-culture and co-culture with Clostridium beijerinckii on cellulose. AMB Express 2017, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Rosa, D.; Medeiros, A.B.P.; Martinez-Burgos, W.J.; do Nascimento, J.R., Jr.; de Carvalho, J.C.; Sydney, E.B.; Soccol, C.R. Biological hydrogen production from palm oil mill effluent (POME) by anaerobic consortia and Clostridium beijerinckii. J. Biotechnol. 2020, 323, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Cao, G.L.; Ren, N.Q. Bioaugmentation with Thermoanaerobacterium thermosaccharolyticum W16 to enhance thermophilic hydrogen production using corn stover hydrolysate. Int. J. Hydrogen Energy 2019, 44, 5821–5829. [Google Scholar] [CrossRef]
- Magnusson, L.; Islam, R.; Sparling, R.; Levin, D.; Cicek, N. Direct hydrogen production from cellulosic waste materials with a single-step dark fermentation process. Int. J. Hydrogen Energy 2008, 33, 5398–5403. [Google Scholar] [CrossRef]
- Chen, C.C.; Chuang, Y.S.; Lin, C.Y.; Lay, C.H.; Sen, B. Thermophilic dark fermentation of untreated rice straw using mixed cultures for hydrogen production. Int. J. Hydrogen Energy 2012, 37, 15540–15546. [Google Scholar] [CrossRef]
- Sheng, T.; Gao, L.; Zhao, L.; Liu, W.; Wang, A. Direct hydrogen production from lignocellulose by the newly isolated Thermoanaerobacterium thermosaccharolyticum strain DD32. RSC Adv. 2015, 5, 99781–99788. [Google Scholar] [CrossRef]
- Mamimin, C.; Kongjan, P.; O-Thong, S.; Prasertsan, P. Enhancement of biohythane production from solid waste by co-digestion with palm oil mill effluent in two-stage thermophilic fermentation. Int. J. Hydrogen Energy 2019, 44, 17224–17237. [Google Scholar] [CrossRef]
- Mamimin, C.; Jehlee, A.; Seengenyoung, J.; Saelor, S.; Prasertsan, P.; O-Thong, S. Thermophilic hydrogen production from co-fermentation of palm oil mill effluent and decanter cake by Thermoanaerobacterium thermosaccharolitycum PSU-2. Int. J. Hydrogen. Energy 2016, 41, 21692–21701. [Google Scholar] [CrossRef]
- O-Thong, S.; Mamimin, C.; Prasertsan, P. Effect of temperature and initial pH on biohydrogen production from palm oil mill effluent: Long-term evaluation and microbial community analysis. Electron. J. Biotechnol. 2011, 14, 9. [Google Scholar] [CrossRef]
- O-Thong, S.; Hniman, A.; Prasertsan, P.; Imai, T. Biohydrogen production from cassava starch processing wastewater by thermophilic mixed cultures. Int. J. Hydrogen Energy 2011, 36, 3409–3416. [Google Scholar] [CrossRef]
- Lay, C.H.; Wu, J.H.; Hsiao, C.L.; Chang, J.J.; Chen, C.C.; Lin, C.Y. Biohydrogen production from soluble condensed molasses fermentation using anaerobic fermentation. Int. J. Hydrogen Energy 2010, 35, 13445–13451. [Google Scholar] [CrossRef]
- Kan, E. Effects of pretreatments of anaerobic sludge and culture conditions on hydrogen productivity in dark anaerobic fermentation. Renew. Energy 2013, 49, 227–231. [Google Scholar] [CrossRef]
- Pachapur, V.L.; Kutty, P.; Pachapur, P.; Brar, S.K.; Le Bihan, Y.; Galvez-Cloutier, R.; Buelna, G. Seed pretreatment for increased hydrogen production using mixed-culture systems with advantages over pure-culture systems. Energies 2019, 12, 530. [Google Scholar] [CrossRef] [Green Version]
- Salakkam, A.; Sittijunda, S.; Mamimin, C.; Phanduang, O.; Reungsang, A. Valorization of microalgal biomass for biohydrogen generation: A review. Bioresour. Technol. 2021, 322, 124533. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Fuchs, B.M.; Behrens, S. The identification of microorganisms by fluorescence in situ hybridization. Curr. Opin. Microbiol. 2001, 12, 231–236. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O-Thong, S.; Khongkliang, P.; Mamimin, C.; Singkhala, A.; Prasertsan, P.; Birkeland, N.-K. Draft genome sequence of Thermoanaerobacterium sp. strain PSU-2 isolated from thermophilic hydrogen producing reactor. Genom. Data 2017, 12, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Tolvanen, K.E.; Koskinen, P.E.; Ylikoski, A.I.; Ollikka, P.K.; Hemmilä, I.A.; Puhakka, J.A.; Karp, M.T. Quantitative monitoring of a hydrogen-producing Clostridium butyricum strain from a continuous-flow, mixed culture bioreactor employing real-time PCR. Int. J. Hydrogen Energy 2008, 33, 542–549. [Google Scholar] [CrossRef]
- Angelidaki, I.; Sanders, W. Assessment of the anaerobic biodegradability of macropollutants. Rev. Environ. Sci. Bio/Technol. 2004, 3, 117–129. [Google Scholar] [CrossRef]
- Fangkum, A.; Reungsang, A. Biohydrogen production from mixed xylose/arabinose at thermophilic temperature by anaerobic mixed cultures in elephant dung. Int. J. Hydrogen Energy 2011, 36, 13928–13938. [Google Scholar] [CrossRef]
- Zhang, X.J.; Yu, H.Q. Inhibitory effects of butyrate on biological hydrogen production with mixed anaerobic cultures. J. Environ. Manag. 2005, 74, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Keyser, M.; Witthuhn, C.; Lamprecht, C.; Coetzee, M.; Britz, T. PCR-based DGGE fingerprinting and identification of methanogens detected in three different types of UASB granules. Syst. Appl. Microbiol. 2006, 29, 77–84. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Hung, C.H.; Cheng, C.H.; Cheng, L.H.; Liang, C.M.; Lin, C.Y. Application of Clostridium-specific PCR primers on the analysis of dark fermentation hydrogen-producing bacterial community. Int. J. Hydrogen Energy 2008, 33, 1586–1592. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Sen, B.; Lin, C.-Y. Batch fermentative hydrogen production by enriched mixed culture: Combination strategy and their microbial composition. J. Biosci. Bioeng. 2014, 117, 222–228. [Google Scholar] [CrossRef]
- Cardoso, V.; Romao, B.B.; Silva, F.T.M.; Santos, J.G.; Batista, F.R.X.; Ferreira, J.S. Hydrogen production by dark fermentation. Chem. Eng. Trans. 2014, 38, 481–486. [Google Scholar] [CrossRef]
- Darwin; Cord-Ruwisch, R.; Charles, W. Ethanol and lactic acid production from sugar and starch wastes by anaerobic acidification. Eng. Life Sci. 2018, 18, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Moreira, F.S.; Machado, R.G.; Romão, B.B.; Batista, F.R.X.; Ferreira, J.S.; Cardoso, V.L. Improvement of hydrogen production by biological route using repeated batch cycles. Process. Biochem. 2017, 58, 60–68. [Google Scholar] [CrossRef]
- Sikora, A.; Błaszczyk, M.; Jurkowski, M.; Zielenkiewicz, U. Lactic acid bacteria in hydrogen-producing consortia. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; Kongo, M., Ed.; IntechOpen: London, UK, 2013; pp. 487–514. [Google Scholar]
- Dimitrellos, G.; Lyberatos, G.; Antonopoulou, G. Does acid addition improve liquid hot water pretreatment of lignocellulosic biomass towards biohydrogen and biogas production. Sustainability 2020, 12, 8935. [Google Scholar] [CrossRef]
- Morsy, F.M.; Elbadry, M.; Elbahloul, Y. Semidry acid hydrolysis of cellulose sustained by autoclaving for production of reducing sugars for bacterial biohydrogen generation from various cellulose feedstock. PeerJ 2021, 9, e11244. [Google Scholar] [CrossRef] [PubMed]
- O-Thong, S.; Prasertsan, P.; Karakashev, D.B.; Angelidaki, I. Thermophilic fermentative hydrogen production by the newly isolated Thermoanaerobacterium thermosaccharolyticum PSU-2. Int. J. Hydrogen Energy 2008, 33, 1204–1214. [Google Scholar] [CrossRef]
- Su, X.; Zhao, W.; Xia, D. The diversity of hydrogen-producing bacteria and methanogens within an in-situ coal seam. Biotechnol. Biofuels 2018, 11, 245. [Google Scholar] [CrossRef]
- Burrell, P.C.; O’Sullivan, C.; Song, H.; Clarke, W.P.; Blackall, L.L. Identification, detection, and spatial resolution of Clostridium populations responsible for cellulose degradation in a methanogenic landfill leachate bioreactor. Appl. Environ. Microbiol. 2004, 70, 2414–2419. [Google Scholar] [CrossRef] [Green Version]
- Rasolofo, E.A.; St-Gelais, D.; LaPointe, G.; Roy, D. Molecular analysis of bacterial population structure and dynamics during cold storage of untreated and treated milk. Int. J. Food Microbiol. 2010, 138, 108–118. [Google Scholar] [CrossRef]
- Smith, C.J.; Nedwell, D.B.; Dong, L.F.; Osborn, A.M. Evaluation of quantitative polymerase chain reaction-based approaches for determining gene copy and gene transcription numbers in environmental samples. Environ. Microbiol. 2006, 8, 804–815. [Google Scholar] [CrossRef]
- Chang, J.; Wu, J.; Wen, F.; Hung, K.; Chen, Y.; Hsiao, C.; Lin, C.; Huang, C. Molecular monitoring of microbes in a continuous hydrogen-producing system with different hydraulic retention time. Int. J. Hydrogen Energy 2008, 33, 1579–1585. [Google Scholar] [CrossRef]
- Okonkwo, O.; Lakaniemi, A.-M.; Santala, V.; Karp, M.; Mangayil, R. Quantitative real-time PCR monitoring dynamics of Thermotoga neapolitana in synthetic co-culture for biohydrogen production. Int. J. Hydrogen Energy 2018, 43, 3133–3141. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism Name | Primer Name | Sequence (5′→3′) | Coverage (%) | Specificity (%) | Sequence Length (bp) | References |
|---|---|---|---|---|---|---|
| Clostridium sp. | Chis150F | AAAGGRAGATTAATACCGCATAA | 88.6 | 99.6 | 540 | [25] |
| ClostIR | TTCTTCCTAATCTCTACGCA | |||||
| Thermoanaerobacterium sp. | TherF | GTGGAGAACACGGAGGAAGG | 21.3 | 100 | 93 | This study |
| TherR | CCCTCTGTTCAGGCCATTGT | |||||
| Bacillus licheniformis | BacdF | TGGCTCAGGACGAACGCTG | 100 | 100 | 82 | This study |
| BacdR | CCGCTGACCTAAGGGAGCAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saripan, K.; Mamimin, C.; Imai, T.; Sittijunda, S.; Reungsang, A. q-PCR Methodology for Monitoring the Thermophilic Hydrogen Producers Enriched from Elephant Dung. Fermentation 2022, 8, 506. https://doi.org/10.3390/fermentation8100506
Saripan K, Mamimin C, Imai T, Sittijunda S, Reungsang A. q-PCR Methodology for Monitoring the Thermophilic Hydrogen Producers Enriched from Elephant Dung. Fermentation. 2022; 8(10):506. https://doi.org/10.3390/fermentation8100506
Chicago/Turabian StyleSaripan, Khamanitjaree, Chonticha Mamimin, Tsuyoshi Imai, Sureewan Sittijunda, and Alissara Reungsang. 2022. "q-PCR Methodology for Monitoring the Thermophilic Hydrogen Producers Enriched from Elephant Dung" Fermentation 8, no. 10: 506. https://doi.org/10.3390/fermentation8100506