
Triacyl Glycerols from Yeast-Catalyzed Batch and Fed-Batch Bioconversion of Hydrolyzed Lignocellulose from Cardoon Stalks

 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Yeast Strain
2.2. Biomass Feedstocks and Steam Explosion Pre-Treatment
2.3. Monosaccharides Production: Enzymatic Hydrolysis of WIS
2.4. Batch and Fed-Batch Experiments
2.5. Extraction of Total Intracellular Lipids
2.6. Determination of Fatty Acids (FAs) Profiles of Triacylglycerols (TAGs) by GC
2.7. Statistical Analysis
3. Results
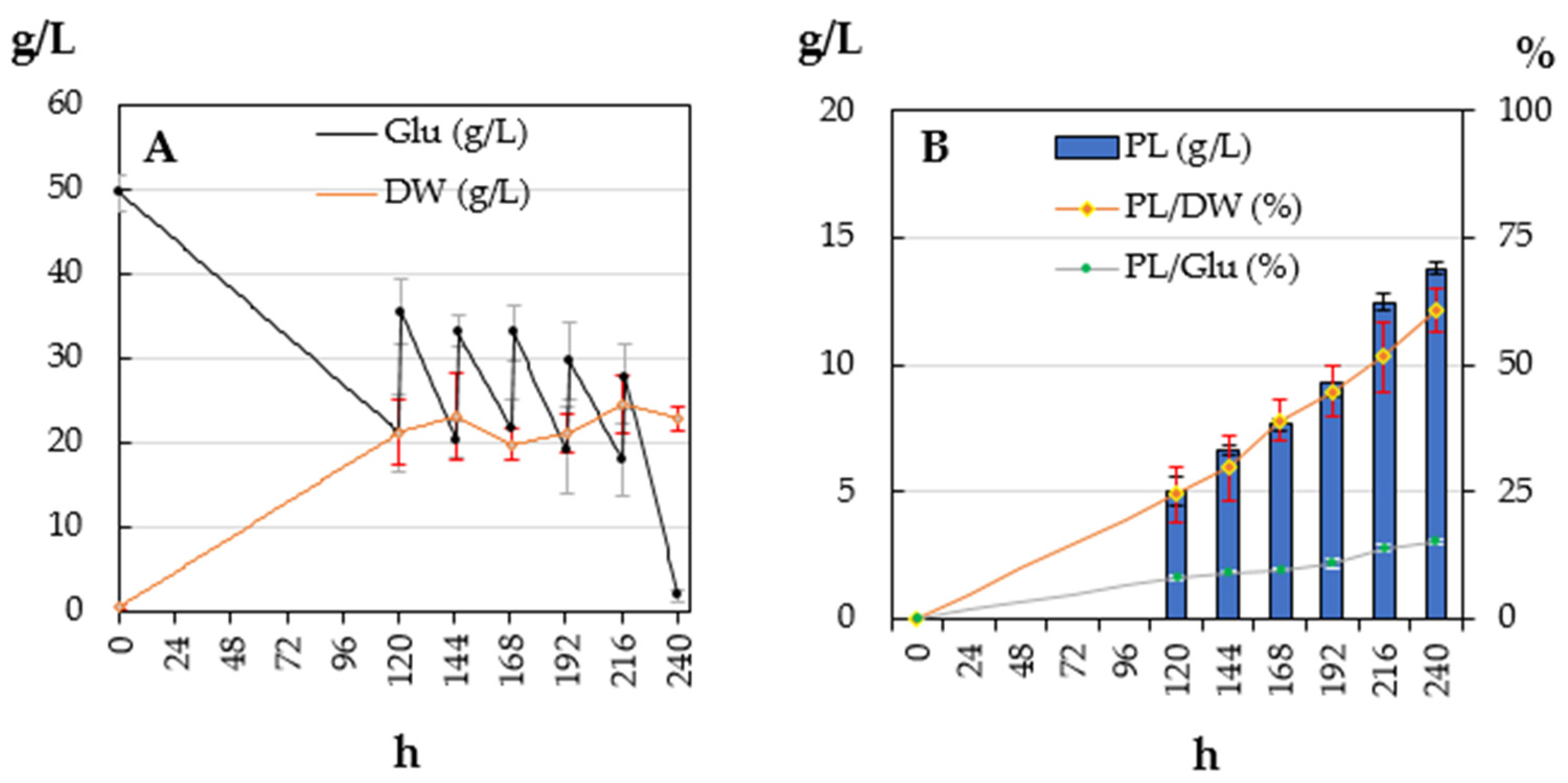
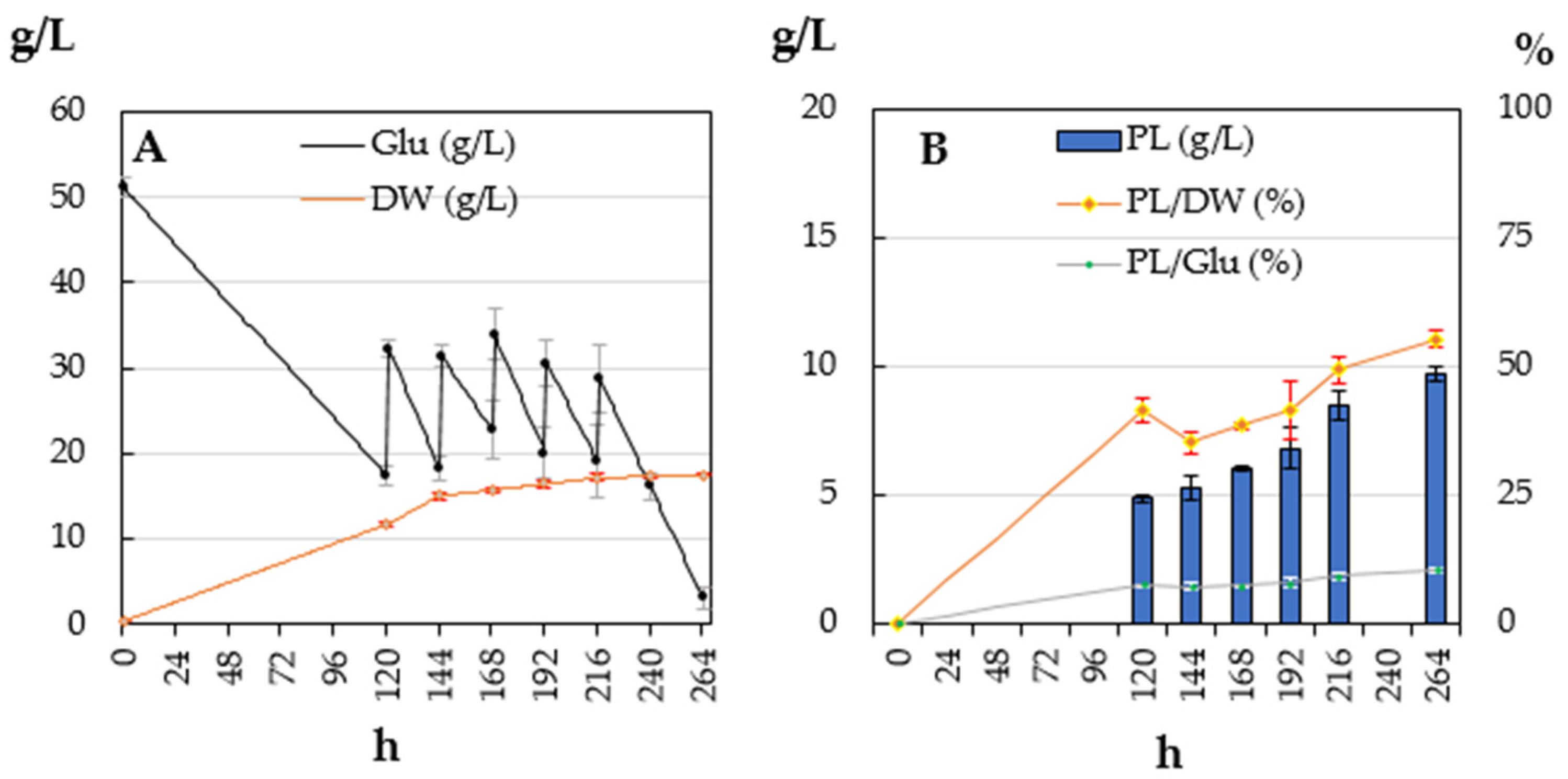
3.1. Lipogenic Aptitude in Batch and Fed-Batch Experiments
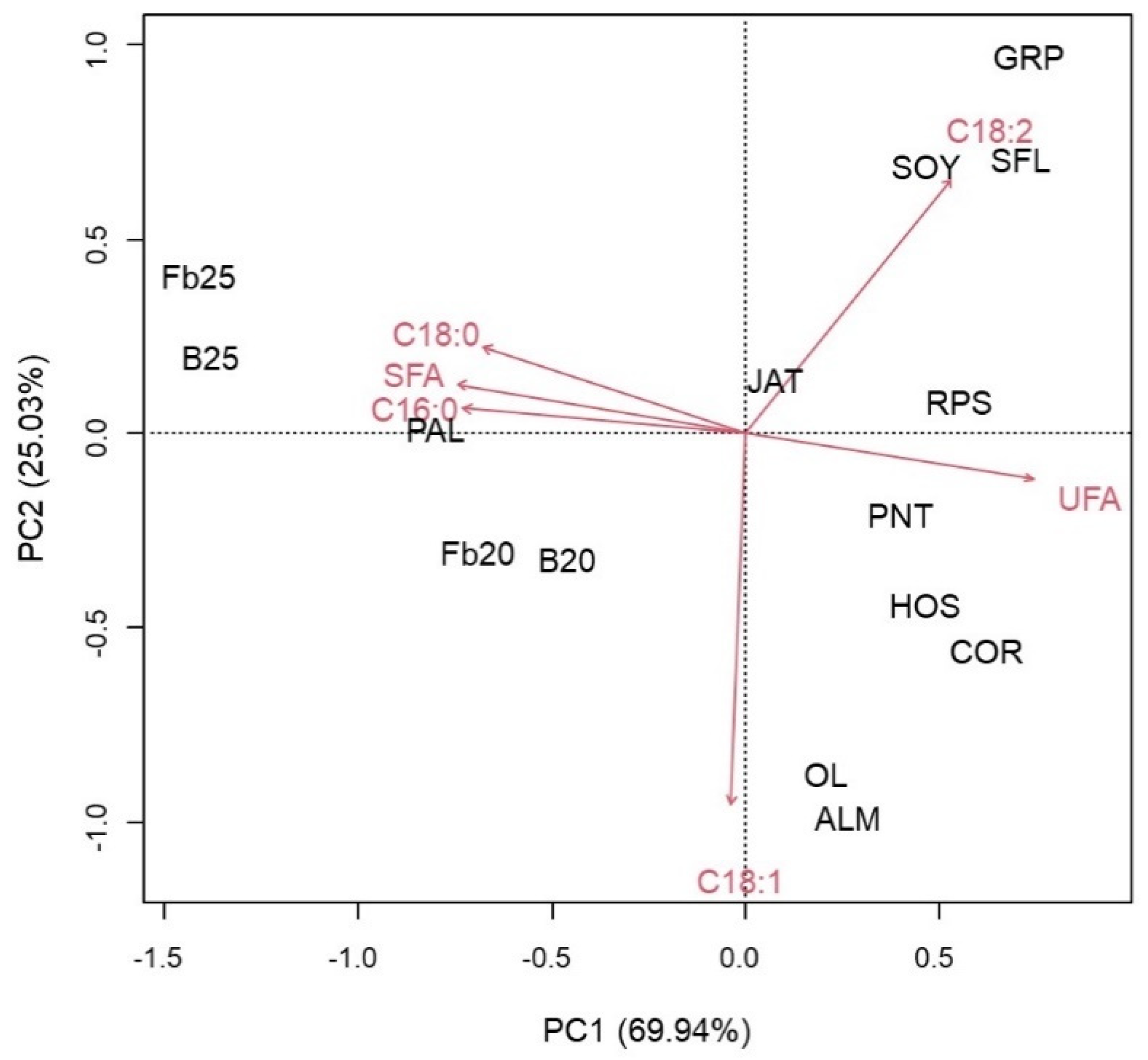
3.2. FAs Profiles of TAGs in Batch and Fed-Batch Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neukirchen, F.; Ries, G. The World of Mineral Deposits: A Beginner’s Guide to Economic Geology; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-34345-3. [Google Scholar]
- Peters, G.P.; Andrew, R.M.; Canadell, J.G.; Friedlingstein, P.; Jackson, R.B.; Korsbakken, J.I.; Le Quéré, C.; Peregon, A. Carbon Dioxide Emissions Continue to Grow amidst Slowly Emerging Climate Policies. Nat. Clim. Chang. 2020, 10, 3–6. [Google Scholar] [CrossRef]
- Umar, M.; Ji, X.; Kirikkaleli, D.; Xu, Q. COP21 Roadmap: Do Innovation, Financial Development, and Transportation Infrastructure Matter for Environmental Sustainability in China? J. Environ. Manag. 2020, 271, 111026. [Google Scholar] [CrossRef]
- Adeniyi, O.M.; Azimov, U.; Burluka, A. Algae Biofuel: Current Status and Future Applications. Renew. Sustain. Energy Rev. 2018, 90, 316–335. [Google Scholar] [CrossRef]
- Zeppini, P.; van den Bergh, J.C.J.M. Global Competition Dynamics of Fossil Fuels and Renewable Energy under Climate Policies and Peak Oil: A Behavioural Model. Energy Policy 2020, 136, 110907. [Google Scholar] [CrossRef]
- Ajanovic, A. Biofuels versus Food Production: Does Biofuels Production Increase Food Prices? Energy 2011, 36, 2070–2076. [Google Scholar] [CrossRef]
- Tsita, K.G.; Kiartzis, S.J.; Ntavos, N.K.; Pilavachi, P.A. Next Generation Biofuels Derived from Thermal and Chemical Conversion of the Greek Transport Sector. Therm. Sci. Eng. Prog. 2020, 17, 100387. [Google Scholar] [CrossRef]
- Abraham, A.; Mathew, A.K.; Park, H.; Choi, O.; Sindhu, R.; Parameswaran, B.; Pandey, A.; Park, J.H.; Sang, B.I. Pretreatment Strategies for Enhanced Biogas Production from Lignocellulosic Biomass. Bioresour. Technol. 2020, 301, 122725. [Google Scholar] [CrossRef]
- Raven, S.; Francis, A.; Srivastava, C.; Kezo, S.; Tiwari, A.; Yadav, A.N.; Singh, S.; Mishra, S.; Gupta, A. Fungal Biofuels: Innovative Approaches. In Recent Advancement in White Biotechnology through Fungi: Volume 2: Perspective for Value-Added Products and Environments; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2019; pp. 385–405. ISBN 978-3-030-14846-1. [Google Scholar]
- Salehi Jouzani, G.; Taherzadeh, M.J. Advances in Consolidated Bioprocessing Systems for Bioethanol and Butanol Production from Biomass: A Comprehensive Review. Biofuel Res. J. 2015, 2, 152–195. [Google Scholar] [CrossRef]
- Salehi Jouzani, G.; Tabatabaei, M.; Aghbashlo, M. Fungi in Fuel Biotechnology; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-44487-7. [Google Scholar]
- Adrio, J.L. Oleaginous Yeasts: Promising Platforms for the Production of Oleochemicals and Biofuels. Biotechnol. Bioeng. 2017, 114, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Z.; Ratledge, C. Single Cell Oils for the 21st Century. In Single Cell Oils; AOCS Publishing: Urbana, IL, USA, 2005; ISBN 978-1-00-304010-1. [Google Scholar]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula Glutinis-Potential Source of Lipids, Carotenoids, and Enzymes for Use in Industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wei, H.; Knoshaug, E.; Van Wychen, S.; Xu, Q.; Himmel, M.E.; Zhang, M. Fatty Alcohol Production in Lipomyces Starkeyi and Yarrowia Lipolytica. Biotechnol. Biofuels 2016, 9, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Huertas, E. Health Effects of Oleic Acid and Long Chain Omega-3 Fatty Acids (EPA and DHA) Enriched Milks. A Review of Intervention Studies. Pharmacol. Res. 2010, 61, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily Yeasts as Oleaginous Cell Factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef]
- Mennella, I.; Savarese, M.; Ferracane, R.; Sacchi, R.; Vitaglione, P. Oleic Acid Content of a Meal Promotes Oleoylethanolamide Response and Reduces Subsequent Energy Intake in Humans. Food Funct. 2015, 6, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Elkacmi, R.; Kamil, N.; Bennajah, M.; Kitane, S. Extraction of Oleic Acid from Moroccan Olive Mill Wastewater. BioMed Res. Int. 2016, 2016, e1397852. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S. Comparative Fatty Acid and Trace Elemental Analysis Identified the Best Raw Material of Jojoba (Simmondsia chinensis) for Commercial Applications. Ann. Agric. Sci. 2018, 63, 37–45. [Google Scholar] [CrossRef]
- Alismaeel, Z.T.; Abbas, A.S.; Albayati, T.M.; Doyle, A.M. Biodiesel from Batch and Continuous Oleic Acid Esterification Using Zeolite Catalysts. Fuel 2018, 234, 170–176. [Google Scholar] [CrossRef]
- Cannilla, C.; Bonura, G.; Costa, F.; Frusteri, F. Biofuels Production by Esterification of Oleic Acid with Ethanol Using a Membrane Assisted Reactor in Vapour Permeation Configuration. Appl. Catal. A Gen. 2018, 566, 121–129. [Google Scholar] [CrossRef]
- Das, M.; Yash, K.; Khan, M.A. Oleaginous Yeast as Potential Lipid Producing Organism: A Review. Plant Arch. 2021, 21, 1830–1833. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of Oleaginous Yeasts. Part II: Technology and Potential Applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous Yeasts for Biodiesel: Current and Future Trends in Biology and Production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, H.; Ahlgren, S.; Sandgren, M.; Passoth, V.; Wallberg, O.; Hansson, P.-A. A Systems Analysis of Biodiesel Production from Wheat Straw Using Oleaginous Yeast: Process Design, Mass and Energy Balances. Biotechnol. Biofuels 2016, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zheng, Y.; Dorgan, K.M.; Chen, S. Oil Production by Oleaginous Yeasts Using the Hydrolysate from Pretreatment of Wheat Straw with Dilute Sulfuric Acid. Bioresour. Technol. 2011, 102, 6134–6140. [Google Scholar] [CrossRef]
- Bonturi, N.; Crucello, A.; Viana, A.J.C.; Miranda, E.A. Microbial Oil Production in Sugarcane Bagasse Hemicellulosic Hydrolysate without Nutrient Supplementation by a Rhodosporidium toruloides Adapted Strain. Process Biochem. 2017, 57, 16–25. [Google Scholar] [CrossRef]
- Tasselli, G.; Filippucci, S.; Borsella, E.; D’Antonio, S.; Gelosia, M.; Cavalaglio, G.; Turchetti, B.; Sannino, C.; Onofri, A.; Mastrolitti, S.; et al. Yeast Lipids from Cardoon Stalks, Stranded Driftwood and Olive Tree Pruning Residues as Possible Extra Sources of Oils for Producing Biofuels and Biochemicals. Biotechnol. Biofuels 2018, 11, 147. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Nassi o Di Nasso, N.; Bonari, E. Long-Term Evaluation of Biomass Production and Quality of Two Cardoon (Cynara cardunculus L.) Cultivars for Energy Use. Biomass Bioenergy 2009, 33, 810–816. [Google Scholar] [CrossRef]
- Ciancolini, A.; Alignan, M.; Pagnotta, M.A.; Vilarem, G.; Crinò, P. Selection of Italian Cardoon Genotypes as Industrial Crop for Biomass and Polyphenol Production. Ind. Crop. Prod. 2013, 51, 145–151. [Google Scholar] [CrossRef]
- Ierna, A.; Mauromicale, G. Cynara cardunculus L. Genotypes as a Crop for Energy Purposes in a Mediterranean Environment. Biomass Bioenergy 2010, 34, 754–760. [Google Scholar] [CrossRef]
- Hsu, Y.-L.; Wu, W.-T. A Novel Approach for Scaling-up a Fermentation System. Biochem. Eng. J. 2002, 11, 123–130. [Google Scholar] [CrossRef]
- Formenti, L.R.; Nørregaard, A.; Bolic, A.; Hernandez, D.Q.; Hagemann, T.; Heins, A.-L.; Larsson, H.; Mears, L.; Mauricio-Iglesias, M.; Krühne, U.; et al. Challenges in Industrial Fermentation Technology Research. Biotechnol. J. 2014, 9, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, C.J.; Nienow, A.W. The Scale-up of Microbial Batch and Fed-Batch Fermentation Processes. Adv. Appl. Microbiol. 2007, 62, 105–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X. Scale-Up of Microbial Fermentation Process. In Manual of Industrial Microbiology and Biotechnology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 669–675. ISBN 978-1-68367-128-2. [Google Scholar]
- Schmidt, F.R. Optimization and Scale up of Industrial Fermentation Processes. Appl. Microbiol. Biotechnol. 2005, 68, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wang, Y.; Wang, J.; Garza, E.; Manow, R.; Zhou, S. Semi-Industrial Scale (30 M3) Fed-Batch Fermentation for the Production of D-Lactate by Escherichia coli Strain HBUT-D15. J. Ind. Microbiol. Biotechnol. 2017, 44, 221–228. [Google Scholar] [CrossRef]
- Ye, H.; He, Y.; Xie, Y.; Sen, B.; Wang, G. Fed-Batch Fermentation of Mixed Carbon Source Significantly Enhances the Production of Docosahexaenoic Acid in Thraustochytriidae sp. PKU#Mn16 by Differentially Regulating Fatty Acids Biosynthetic Pathways. Bioresour. Technol. 2020, 297, 122402. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Yan, S.; Tyagi, R.D.; Drogui, P. Lipid Production from Fed-Batch Fermentation of Crude Glycerol Directed by the Kinetic Study of Batch Fermentations. Fuel 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Fei, Q.; Wewetzer, S.J.; Kurosawa, K.; Rha, C.; Sinskey, A.J. High-Cell-Density Cultivation of an Engineered Rhodococcus opacus Strain for Lipid Production via Co-Fermentation of Glucose and Xylose. Process Biochem. 2015, 50, 500–506. [Google Scholar] [CrossRef]
- Ji, F.; Zhou, Y.; Pang, A.; Ning, L.; Rodgers, K.; Liu, Y.; Dong, R. Fed-Batch Cultivation of Desmodesmus sp. in Anaerobic Digestion Wastewater for Improved Nutrient Removal and Biodiesel Production. Bioresour. Technol. 2015, 184, 116–122. [Google Scholar] [CrossRef]
- Filippucci, S.; Tasselli, G.; Scardua, A.; Di Mauro, S.; Cramarossa, M.R.; Perini, D.; Turchetti, B.; Onofri, A.; Forti, L.; Buzzini, P. Study of Holtermanniella wattica, Leucosporidium creatinivorum, Naganishia adeliensis, Solicoccozyma aeria, and Solicoccozyma terricola for Their Lipogenic Aptitude from Different Carbon Sources. Biotechnol. Biofuels 2016, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Cavalaglio, G.; Gelosia, M.; Giannoni, T.; Barros Lovate Temporim, R.; Nicolini, A.; Cotana, F.; Bertini, A. Acid-catalyzed steam explosion for high enzymatic saccharification and low inhibitor release from lignocellulosic cardoon stalks. Biochem. Eng. J. 2021, 174, 108121. [Google Scholar] [CrossRef]
- Sluiter, J.B.; Ruiz, R.O.; Scarlata, C.J.; Sluiter, A.D.; Templeton, D.W. Compositional Analysis of Lignocellulosic Feedstocks. 1. Review and Description of Methods. J. Agric. Food Chem. 2010, 58, 9043–9053. [Google Scholar] [CrossRef]
- Sluiter, J.; Sluiter, A. Summative Mass Closure; NREL/TP-510-48087; NREL: Golden, CO, USA, 2010; pp. 1–10.
- Cavalaglio, G.; Gelosia, M.; D’Antonio, S.; Nicolini, A.; Pisello, A.L.; Barbanera, M.; Cotana, F. Lignocellulosic Ethanol Production from the Recovery of Stranded Driftwood Residues. Energies 2016, 9, 634. [Google Scholar] [CrossRef] [Green Version]
- Cara, C.; Ruiz, E.; Oliva, J.M.; Sáez, F.; Castro, E. Conversion of Olive Tree Biomass into Fermentable Sugars by Dilute Acid Pretreatment and Enzymatic Saccharification. Bioresour. Technol. 2008, 99, 1869–1876. [Google Scholar] [CrossRef]
- Barakat, A.; de Vries, H.; Rouau, X. Dry Fractionation Process as an Important Step in Current and Future Lignocellulose Biorefineries: A Review. Bioresour. Technol. 2013, 134, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, J.; Blomqvist, J.; Pickova, J.; Bonturi, N.; Sandgren, M.; Passoth, V. Lipid Production from Hemicellulose with Lipomyces Starkeyi in a PH Regulated Fed-Batch Cultivation. Yeast 2016, 33, 451–462. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. Official Methods of Analysis, 16th ed.; Garrard Press: Gaithersburg, MA, USA, 1995. [Google Scholar]
- Li, Q.; Du, W.; Liu, D. Perspectives of Microbial Oils for Biodiesel Production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Z.; Bai, F. High-density cultivation of oleaginous yeast Rhodosporidium toruloides Y4 in fed-batch culture. Enzyme Microb. Technol. 2007, 41, 312–317. [Google Scholar] [CrossRef]
- Hassan, M.; Blanc, P.J.; Louis-marie, G.; Pareilleux, A.; Goma, G. Influence of nitrogen and iron limitations on lipid production by Cryptococcus curvatus grown in batch and fed-batch culture. Process Biochem. 1996, 31, 355–361. [Google Scholar] [CrossRef]
- Vishniac, H.S. Yeast Biodiversity in the Antarctic. In Biodiversity and Ecophysiology of Yeasts; Péter, G., Rosa, C., Eds.; The Yeast Handbook; Springer: Berlin/Heidelberg, Germany, 2006; pp. 419–440. ISBN 978-3-540-30985-7. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 13 November 2021).
- Patel, A.; Arora, N.; Mehtani, J.; Pruthi, V.; Pruthi, P. Assessment of Fuel Properties on the Basis of Fatty Acid Profiles of Oleaginous Yeast for Potential Biodiesel Production. Renew. Sustain. Energy Rev. 2017, 77, 604–616. [Google Scholar] [CrossRef] [Green Version]
- Akbar, E.; Yaakob, Z.; Kamarudin, S.; Ismail, M.; Salimon, J. Characteristic and Composition of Jatropha curcas Oil Seed from Malaysia and Its Potential as Biodiesel Feedstock Feedstock. Eur. J. Sci. Res. 2009, 29, 396–403. [Google Scholar]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, A. Influence of Fatty Acid Composition of Raw Materials on Biodiesel Properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef]
- Liu, J.; Huang, X.; Chen, R.; Yuan, M.; Liu, J. Efficient Bioconversion of High-Content Volatile Fatty Acids into Microbial Lipids by Cryptococcus curvatus ATCC 20509. Bioresour. Technol. 2017, 239, 394–401. [Google Scholar] [CrossRef]
- Murugan, S.; Bharathipriyadharsini, M.; Singaram, J. A Study on the Potential of Oleaginous Yeast in Producing Lipid from Garbage. Int. J. Appl. Eng. Res. 2015, 10, 346–350. [Google Scholar]
- Sitepu, I.R.; Sestric, R.; Ignatia, L.; Levin, D.; German, J.B.; Gillies, L.A.; Almada, L.A.G.; Boundy-Mills, K.L. Manipulation of culture conditions alters lipid content and fatty acid profiles of a wide variety of known and new oleaginous yeast species. Bioresour. Technol. 2013, 144, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Raimondi, S.; Rossi, M.; Leonardi, A.; Bianchi, M.; Rinaldi, T.; Amaretti, A. Getting lipids from glycerol: New perspectives on biotechnological exploitation of Candida freyschussii. Microb. Cell Fact. 2014, 13, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, W.; Li, Z.; Wang, X.; Gao, R.; Zhou, X.; Cheng, S.; Men, Y.; Zheng, L. Approaches to improve the lipid synthesis of oleaginous yeast Yarrowia lipolytica: A review. Renew. Sustain. Energy Rev. 2021, 149, 111386. [Google Scholar] [CrossRef]
- Ratledge, C. The role of malic enzyme as the provider of NADPH in oleaginous microorganisms: A reappraisal and unsolved problems. Biotechnol. Lett. 2014, 36, 1557–1568. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The Role of Temperature, PH and Nutrition in Process Development of the Unique Oleaginous Yeast Metschnikowia Pulcherrima. J. Chem. Technol. Biotechnol. 2020, 95, 1163–1172. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakumoto, N.; Kaneko, Y.; Harashima, S. Mga2p Is a Putative Sensor for Low Temperature and Oxygen to Induce OLE1 Transcription in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2002, 291, 707–713. [Google Scholar] [CrossRef]
- Tezaki, S.; Iwama, R.; Kobayashi, S.; Shiwa, Y.; Yoshikawa, H.; Ohta, A.; Horiuchi, H.; Fukuda, R. Δ12-Fatty Acid Desaturase Is Involved in Growth at Low Temperature in Yeast Yarrowia lipolytica. Biochem. Biophys. Res. Commun. 2017, 488, 165–170. [Google Scholar] [CrossRef]
- Parsons, S.; Abeln, F.; McManus, M.C.; Chuck, C.J. Techno-Economic Analysis (TEA) of Microbial Oil Production from Waste Resources as Part of a Biorefinery Concept: Assessment at Multiple Scales under Uncertainty. J. Chem. Technol. Biotechnol. 2019, 94, 701–711. [Google Scholar] [CrossRef]
- OECD-FAO Agricultural Outlook 2017–2026 Special Focus: Southeast Asia; OECD/FAO: Rome, Italy, 2017.
- Parsons, S.; Raikova, S.; Chuck, C.J. The Viability and Desirability of Replacing Palm Oil. Nat. Sustain. 2020, 3, 412–418. [Google Scholar] [CrossRef]
- Wicke, B.; Sikkema, R.; Dornburg, V.; Faaij, A. Exploring Land Use Changes and the Role of Palm Oil Production in Indonesia and Malaysia. Land Use Policy 2011, 28, 193–206. [Google Scholar] [CrossRef]
- Fayle, T.M.; Turner, E.C.; Snaddon, J.L.; Chey, V.K.; Chung, A.Y.C.; Eggleton, P.; Foster, W.A. Oil Palm Expansion into Rain Forest Greatly Reduces Ant Biodiversity in Canopy, Epiphytes and Leaf-Litter. Basic Appl. Ecol. 2010, 11, 337–345. [Google Scholar] [CrossRef]
- Danielsen, F.; Beukema, H.; Burgess, N.D.; Parish, F.; Brühl, C.A.; Donald, P.F.; Murdiyarso, D.; Phalan, B.; Reijnders, L.; Struebig, M.; et al. Biofuel Plantations on Forested Lands: Double Jeopardy for Biodiversity and Climate. Conserv. Biol. 2009, 23, 348–358. [Google Scholar] [CrossRef]
- Germer, J.; Sauerborn, J. Estimation of the Impact of Oil Palm Plantation Establishment on Greenhouse Gas Balance. Environ Dev. Sustain. 2008, 10, 697–716. [Google Scholar] [CrossRef]
- Bessou, C.; Chase, L.D.C.; Henson, I.E.; Abdul-Manan, A.F.N.; Milà i Canals, L.; Agus, F.; Sharma, M.; Chin, M. Pilot Application of PalmGHG, the Roundtable on Sustainable Palm Oil Greenhouse Gas Calculator for Oil Palm Products. J. Clean. Prod. 2014, 73, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Angerbauer, C.; Siebenhofer, M.; Mittelbach, M.; Guebitz, G.M. Conversion of Sewage Sludge into Lipids by Lipomyces Starkeyi for Biodiesel Production. Bioresour. Technol. 2008, 99, 3051–3056. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Zong, M.H.; Wu, H. Efficient Lipid Production with Trichosporonfermentans and Its Use for Biodiesel Preparation. Bioresour. Technol. 2008, 99, 7881–7885. [Google Scholar] [CrossRef]
- Vaishnav, A. Microbial Polymers: Applications and Ecological Perspectives; Springer Nature: Basingstoke, UK, 2021. [Google Scholar]
- Rossi, M.; Buzzini, P.; Cordisco, L.; Amaretti, A.; Sala, M.; Raimondi, S.; Ponzoni, C.; Pagnoni, U.M.; Matteuzzi, D. Growth, lipid accumulation, and fatty acid composition in obligate psychrophilic, facultative psychrophilic, and mesophilic yeasts. FEMS Microbiol. Ecol. 2009, 69, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Chebbi, H.; Leiva-Candia, D.; Carmona-Cabello, M.; Jaouani, A.; Dorado, M.P. Biodiesel Production from Microbial Oil Provided by Oleaginous Yeasts from Olive Oil Mill Wastewater Growing on Industrial Glycerol. Ind. Crop. Prod. 2019, 139, 111535. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable Biodiesel Production from Oleaginous Yeasts Utilizing Hydrolysates of Various Non-Edible Lignocellulosic Biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Rizwanul Fattah, I.M.; Masjuki, H.H.; Kalam, M.A.; Hazrat, M.A.; Masum, B.M.; Imtenan, S.; Ashraful, A.M. Effect of Antioxidants on Oxidation Stability of Biodiesel Derived from Vegetable and Animal Based Feedstocks. Renew. Sustain. Energy Rev. 2014, 30, 356–370. [Google Scholar] [CrossRef]
- Castellini, M.; Ubertini, S.; Barletta, D.; Baffo, I.; Buzzini, P.; Barbanera, M. Techno-Economic Analysis of Biodiesel Production from Microbial Oil Using Cardoon Stalks as Carbon Source. Energies 2021, 14, 1473. [Google Scholar] [CrossRef]
- Barbanera, M.; Castellini, M.; Tasselli, G.; Turchetti, B.; Cotana, F.; Buzzini, P. Prediction of the Environmental Impacts of Yeast Biodiesel Production from Cardoon Stalks at Industrial Scale. Fuel 2021, 283, 118967. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| T (°C) | DW (g/L of Substrate) | PL (g/L of Substrate) | PL/DW (%) | PL/Glu (%) | PL/d (g/L of Substrate × Day) |
|---|---|---|---|---|---|
| 20 | 17.58 ± 1.03 a | 7.13 ± 0.38 a | 40.7 ± 4.38 a | 14.24 ± 1.83 a | 1.43 ± 0.08 a |
| 25 | 15.96 ± 0.76 a | 6.68 ± 0.48 a | 41.94 ± 3.83 a | 14.64 ± 0.99 a | 1.34 ± 0.10 a |
| T (°C) | C14:0 (%) | C16:0 (%) | C16:1 (%) | C18:0 (%) | C18:1 (%) | C18:2 (%) | |
| Batch | 20 | 0.00 a | 20.99 ± 0.14 a | 0.63 ± 0.05 a | 10.54 ± 0.22 a | 58.95 ± 0.07 a | 8.06 ± 0.15 a |
| 25 | 0.49 ± 0.051 b | 33.13 ± 0.71b | 0.2 ± 0.34 a | 20.38 ± 1.87 b | 44.06 ± 0.9 b | 1.05 ± 0.09 b | |
| Fed-batch | 20 | 0.79 ± 0.11 a | 29.53 ± 0.93 a | 1.46 ± 0.21 a | 7.97± 0.68 a | 56.19± 0.26 a | 2.5 ± 0.44 a |
| 25 | 0.39 ± 0.04 b | 36.67 ± 1.31 b | 1.03 ± 0.07 a | 19.81 ± 1.2 b | 36.75 ± 0.52 b | 4.4 ± 0.52 b | |
| T (°C) | C20:0 (%) | C22:0 (%) | C24:0 (%) | SFA (%) | UFA (%) | DU (%) | |
| Batch | 20 | 0.00 | 0.00 | 0.83 ± 0.04 a | 32.36 | 67.64 | 75.70 |
| 25 | 0.00 | 0.00 | 0.7 ± 0.13 a | 54.69 | 45.31 | 46.36 | |
| Fed-batch | 20 | 0.35 ± 0.26 a | 0.11 ± 0.01 a | 0.6 ± 0.12 a | 39.75 | 60.25 | 63.03 |
| 25 | 0.43 ± 0.02 a | 0.22 ± 0.02 b | 0.2 ± 0.01 b | 57.78 | 42.22 | 46.95 |
| T (°C) | CLSF (%) | OS (h) | CFPP (°C) | KV (mm2/s) | D (g/cm3) | |
| Reference values/ranges | Not specified | ≥6 h | Variable * | 3.5–5 | 0.86–0.90 | |
| Batch experiments | 20 | 10.91 | 4.86 | 4.38 | 4.63 | 0.87 |
| 25 | 18.41 | 5.99 | 17.55 | 4.74 | 0.86 | |
| Fed-batch experiments | 20 | 12.75 | 5.35 | 7.61 | 4.56 | 0.87 |
| 25 | 19.28 | 5.97 | 19.08 | 4.73 | 0.86 | |
| T (°C) | SV (mg KOH/g of oil) | IV (mg I2/100 g) | CN | HHV (MJ/Kg) | ||
| Reference values/ranges | ≥0.50 | ≤120 | ≥51 | Not specified | ||
| Batch experiments | 20 | 192.42 | 65.00 | 58.09 | 39.87 | |
| 25 | 194.61 | 39.74 | 64.21 | 39.86 | ||
| Fed-batch experiments | 20 | 194.91 | 54.14 | 60.50 | 39.86 | |
| 25 | 195.91 | 40.34 | 63.87 | 39.92 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, D.; Sannino, C.; Giannoni, T.; Fabbrizi, G.; Gelosia, M.; Nicolini, A.; Turchetti, B.; Cotana, F.; Buzzini, P. Triacyl Glycerols from Yeast-Catalyzed Batch and Fed-Batch Bioconversion of Hydrolyzed Lignocellulose from Cardoon Stalks. Fermentation 2021, 7, 315. https://doi.org/10.3390/fermentation7040315
Aiello D, Sannino C, Giannoni T, Fabbrizi G, Gelosia M, Nicolini A, Turchetti B, Cotana F, Buzzini P. Triacyl Glycerols from Yeast-Catalyzed Batch and Fed-Batch Bioconversion of Hydrolyzed Lignocellulose from Cardoon Stalks. Fermentation. 2021; 7(4):315. https://doi.org/10.3390/fermentation7040315
Chicago/Turabian StyleAiello, Domenico, Ciro Sannino, Tommaso Giannoni, Giacomo Fabbrizi, Mattia Gelosia, Andrea Nicolini, Benedetta Turchetti, Franco Cotana, and Pietro Buzzini. 2021. "Triacyl Glycerols from Yeast-Catalyzed Batch and Fed-Batch Bioconversion of Hydrolyzed Lignocellulose from Cardoon Stalks" Fermentation 7, no. 4: 315. https://doi.org/10.3390/fermentation7040315