
Optimization of Soybean Meal Fermentation for Aqua-Feed with Bacillus subtilis natto Using the Response Surface Methodology

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Starter Culture Preparation
2.3. Solid State Fermentation and Single Factor Tests
2.4. Optimization of Fermentation Process by RSM
2.5. Samples
2.6. Degree of Protein Hydrolysis Analysis
2.7. Nutritional Analysis
2.8. ANFs Assays of SBM and FSM
2.9. SDS-Polyacrylamide Gel Electrophoresis (PAGE) for Protein Separation
2.10. Statistical Analysis
3. Results and Discussion
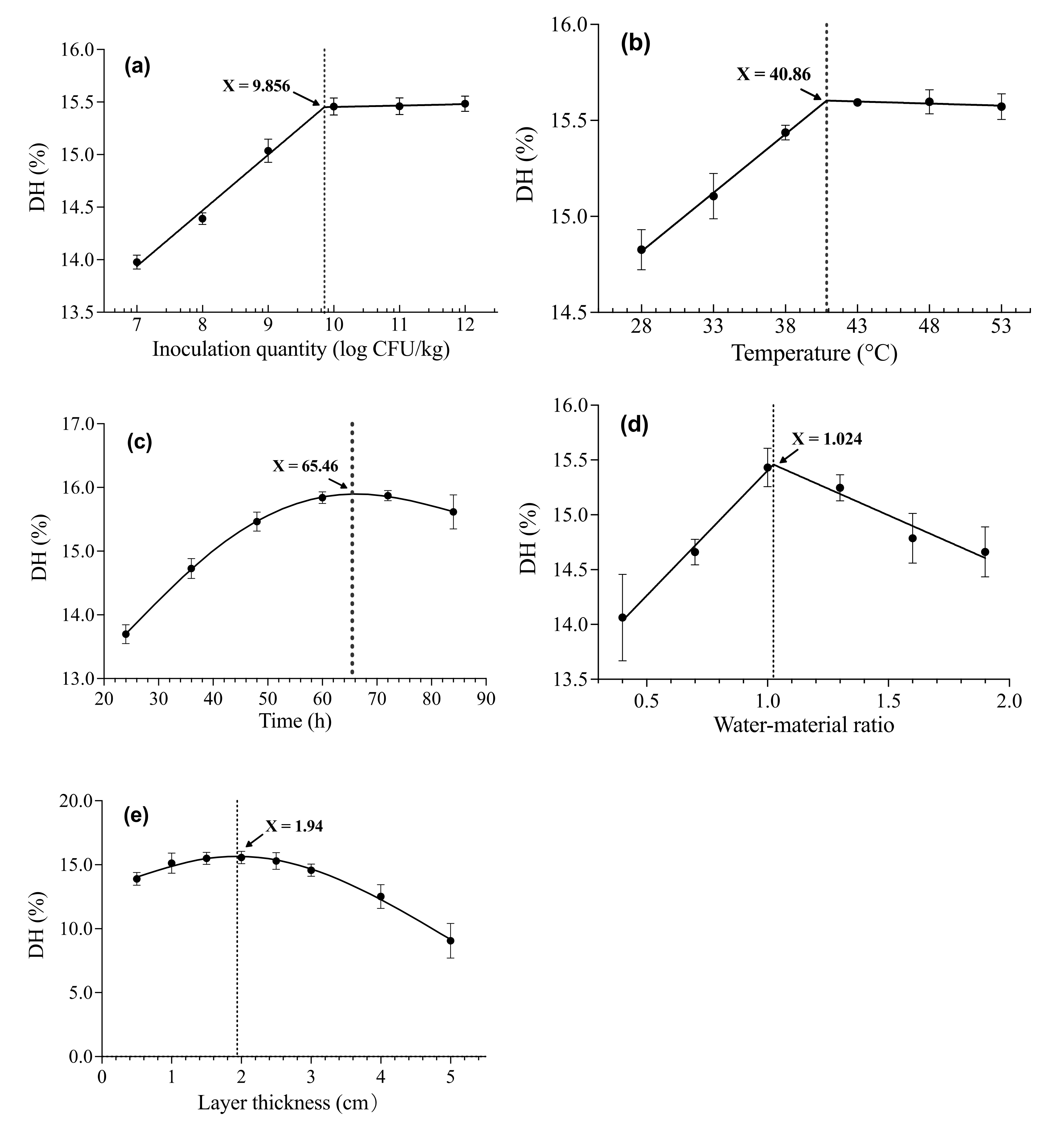
3.1. Single Factor Tests
3.1.1. Effect of Inoculation Quantity of B. s. natto on the DH
3.1.2. Effect of Fermentation Temperature on the DH
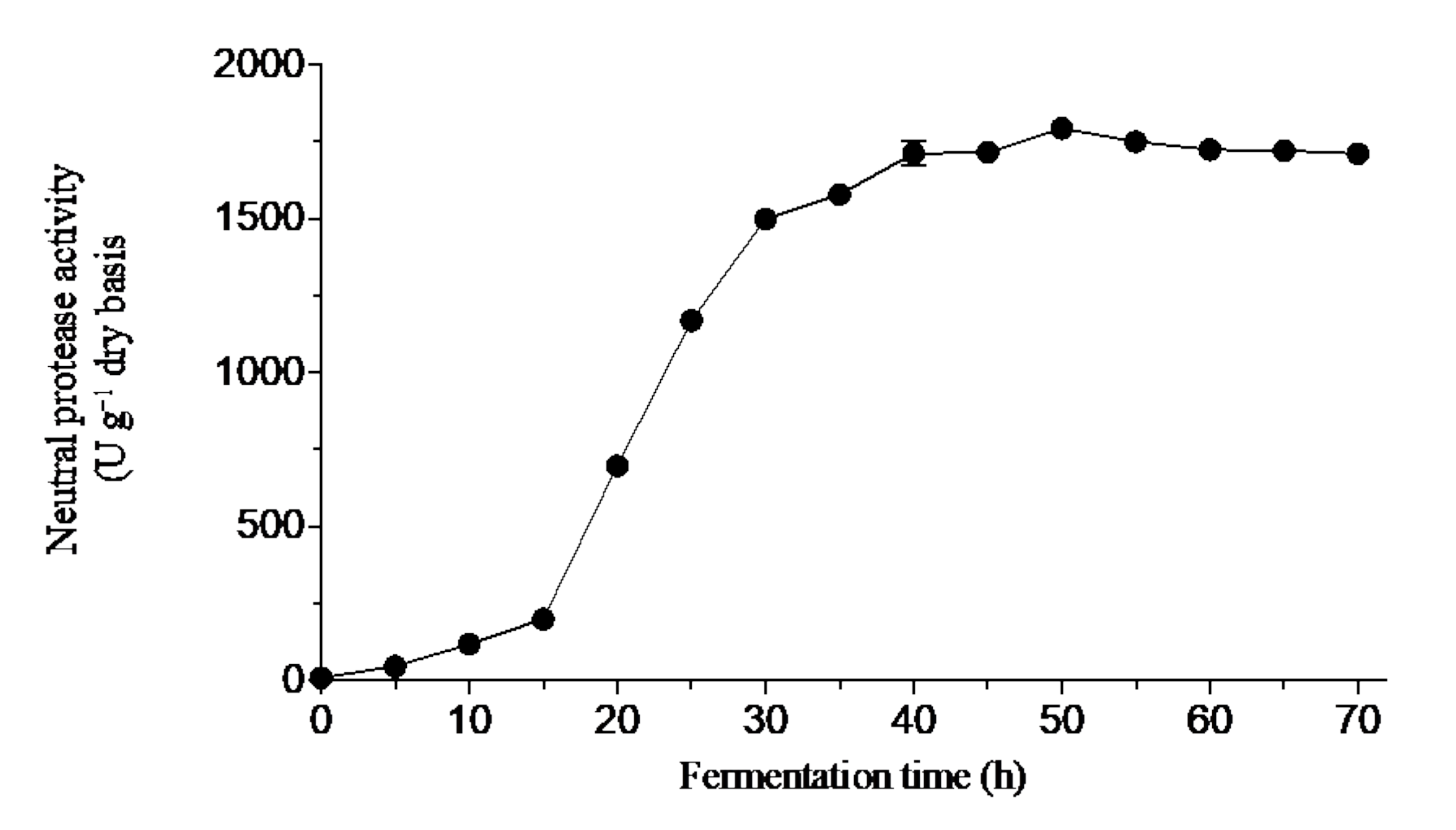
3.1.3. Effect of Fermentation Time on the DH
3.1.4. Effect of Water–Material Ratio on the DH
3.1.5. Effect of SBM Layer Thickness on the DH
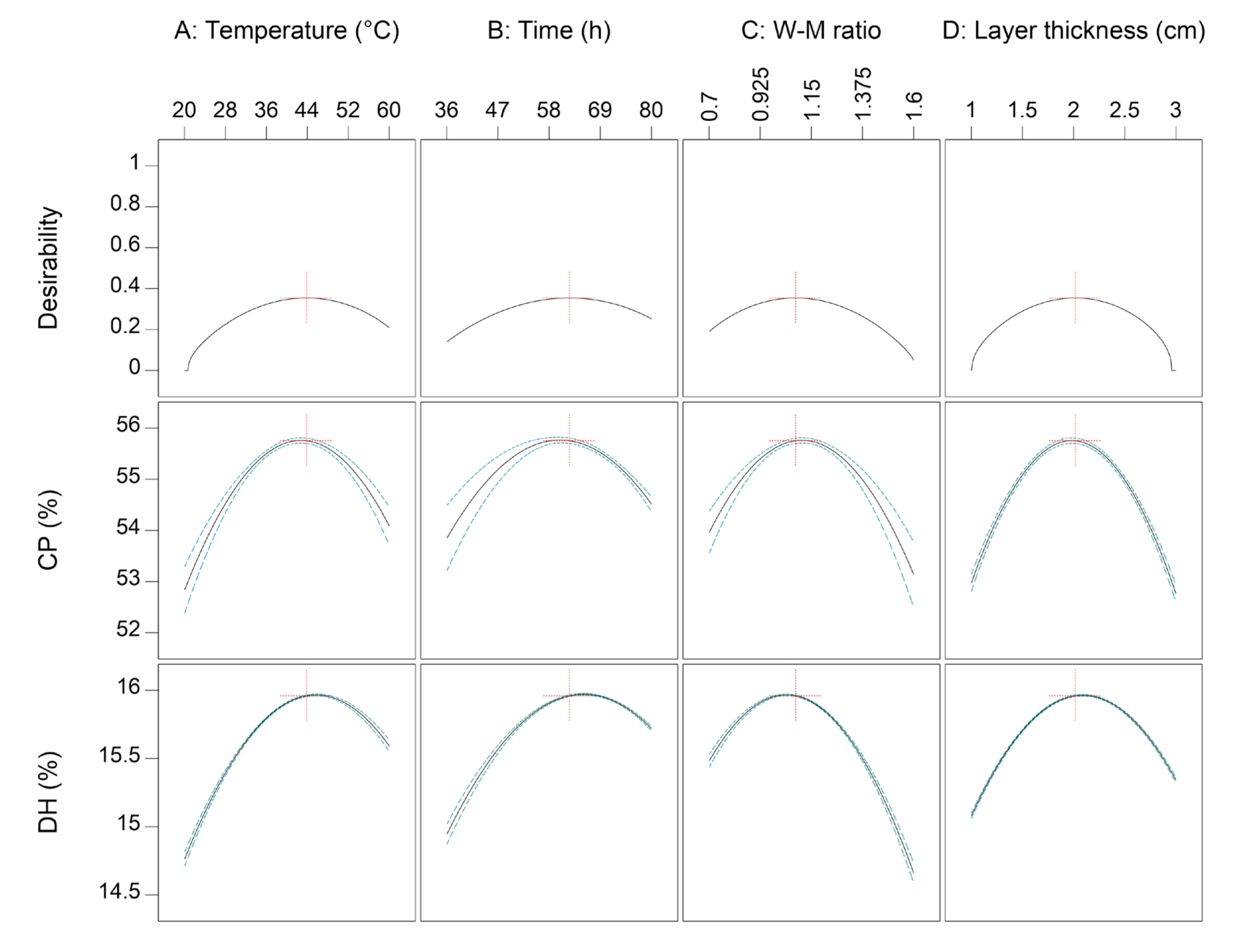
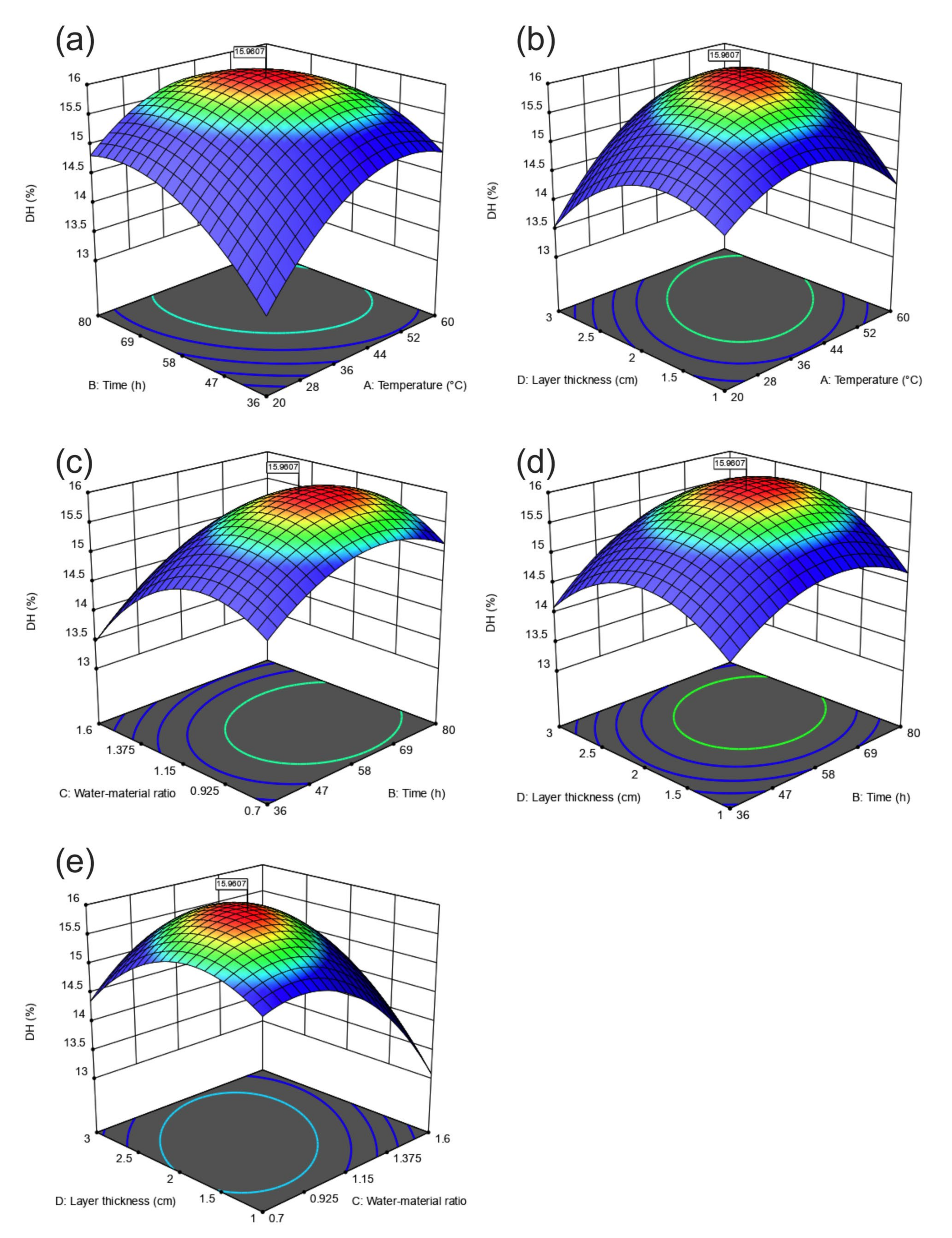
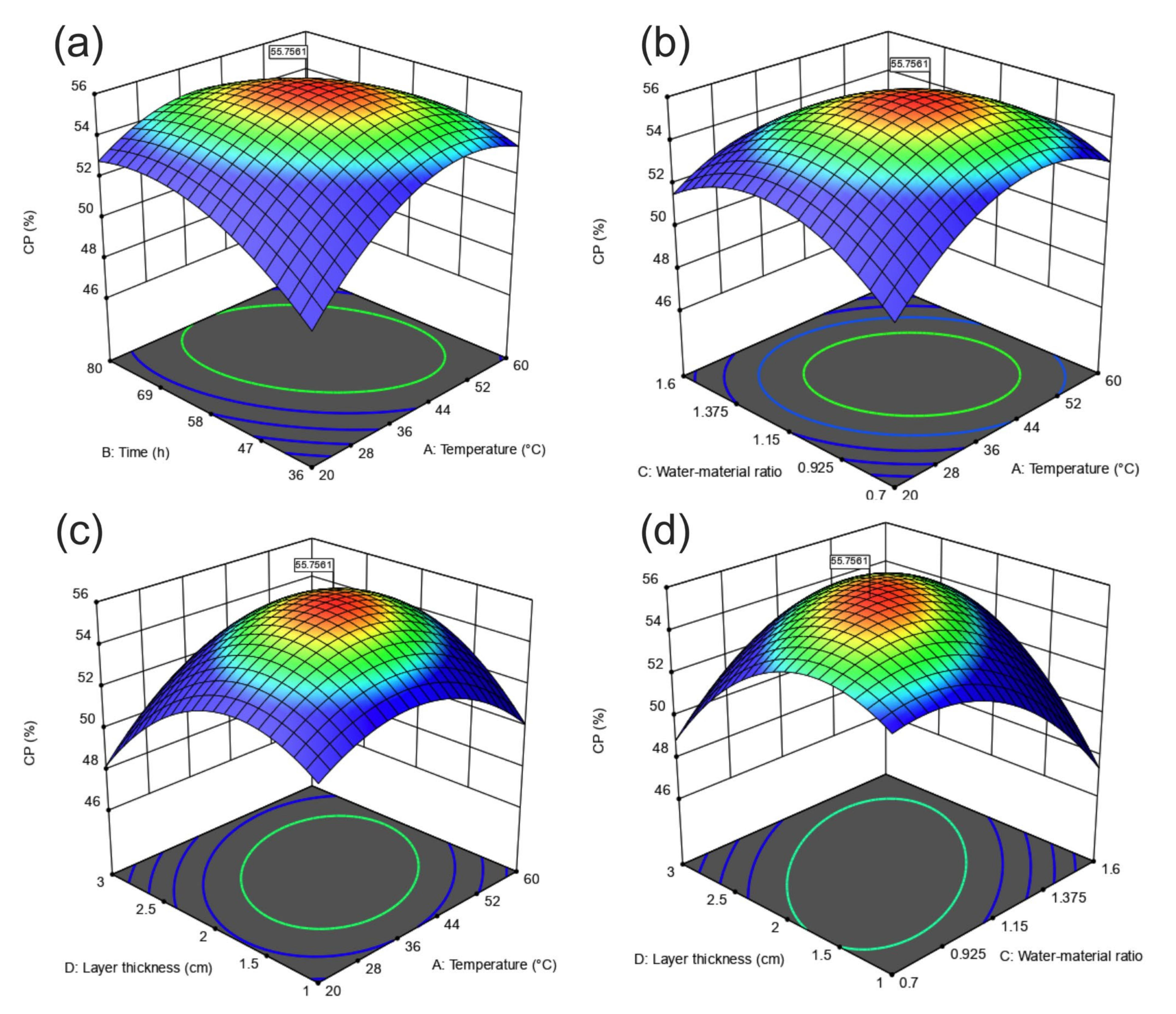
3.2. Further Optimization Using CCD
3.2.1. Variance and Regression Analysis of CCD Design
3.2.2. Response Surface Interaction Analysis
3.2.3. Validation of the Optimization Fermentation Medium
3.3. Comparison of Nutritional Values of FSM and SBM
3.3.1. Nutrient Composition
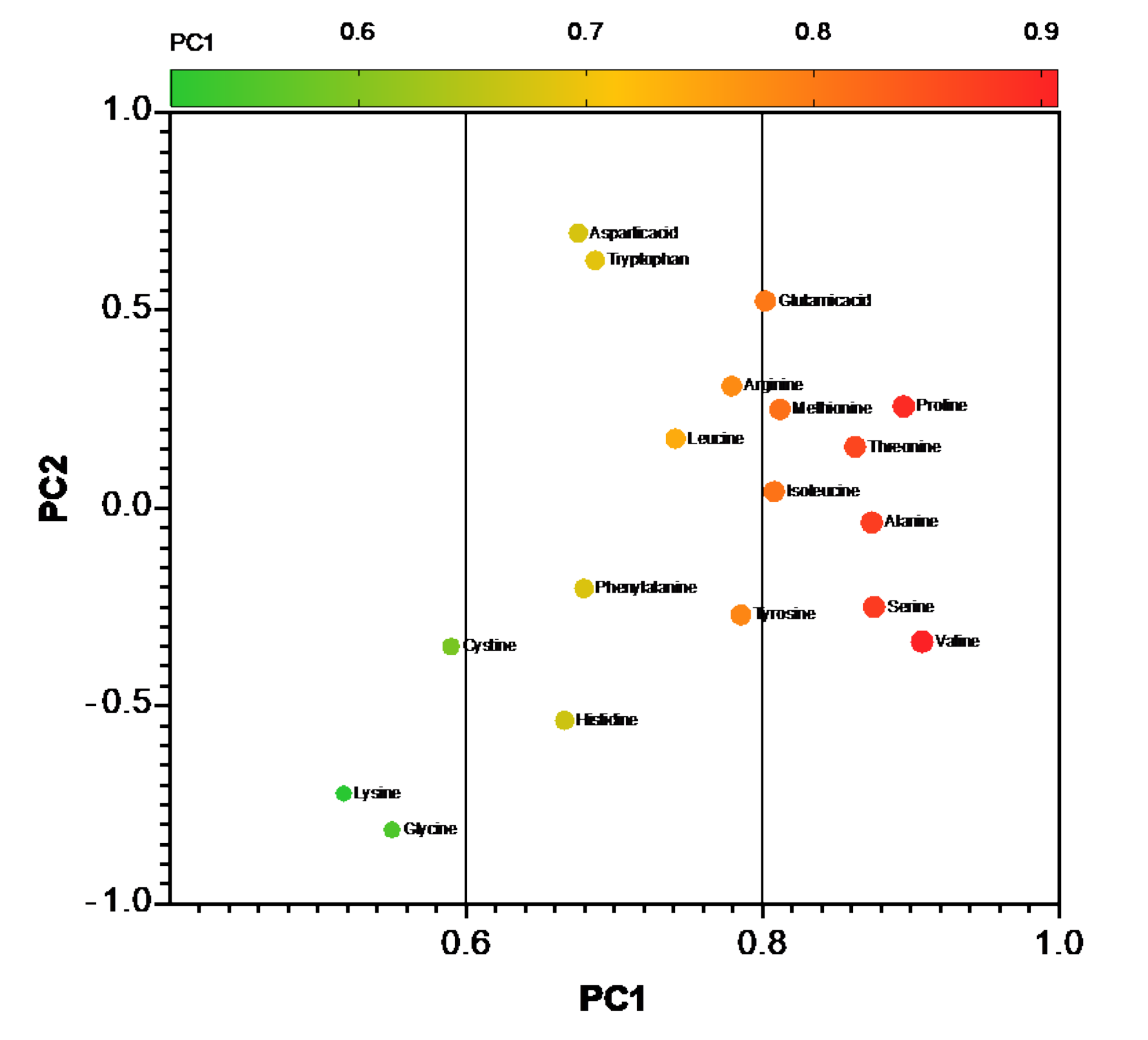
3.3.2. Amino Acid Composition of SBM and FSM
3.3.3. ANFs Composition
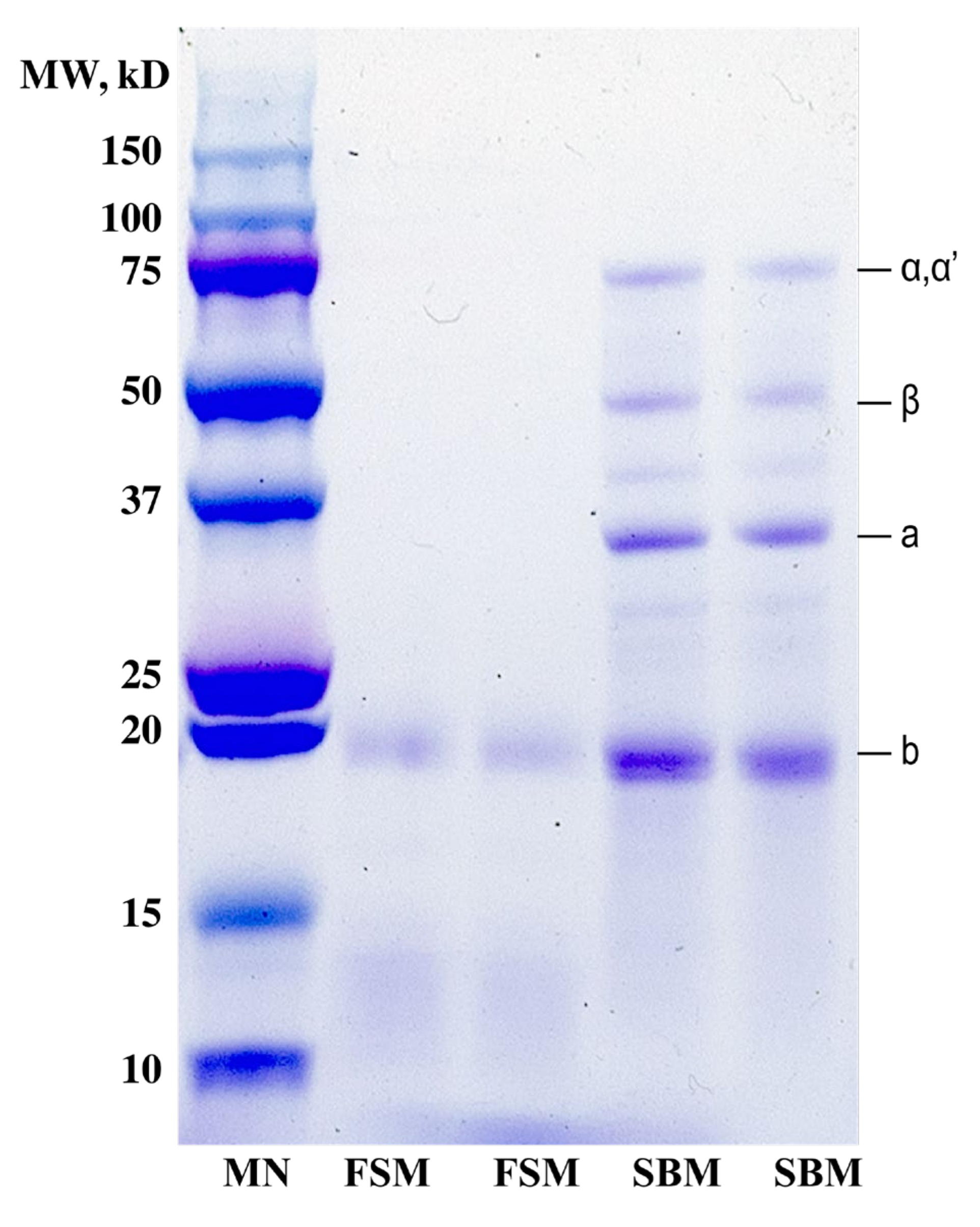
3.3.4. SDS-PAGE Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Luthada-Raswiswi, R.; Mukaratirwa, S.; O’Brien, G. Animal protein sources as a substitute for fishmeal in aquaculture diets: A systematic review and meta-analysis. Appl. Sci. 2021, 11, 3854. [Google Scholar] [CrossRef]
- Ayadi, F.Y.; Rosentrate, K.A.; Muthukumar, K. Alternative protein sources for aquaculture feeds. J. Aquac. Feed. Sci. Nutr. 2012, 4, 1–26. [Google Scholar] [CrossRef]
- Zhou, Q.-C.; Tan, B.-P.; Mai, K.-S.; Liu, Y.-J. Apparent digestibility of selected feed ingredients for juvenile cobia Rachycentron canadum. Aquaculture 2004, 241, 441–451. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2016. Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016; ISBN 9789251091852. [Google Scholar]
- Shao, J.; Wang, L.; Shao, X.; Liu, M. Dietary different replacement levels of fishmeal by fish silage could influence growth of Litopenaeus vannamei by regulating mTOR at transcriptional level. Front. Physiol. 2020, 11, 359. [Google Scholar] [CrossRef]
- Cashion, T.; Le Manach, F.; Zeller, D.; Pauly, D. Most fish destined for fishmeal production are food-grade fish. Fish Fish. 2017, 18, 837–844. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Fish matters: Importance of aquatic foods in human nutrition and global food supply. Rev. Fish. Sci. 2013, 21, 22–38. [Google Scholar] [CrossRef]
- Lim, C.; Webster, C.D.; Lee, C.S. Alternative Protein Sources in Aquaculture Diets; Haworth Press: New York, NY, USA, 2008; ISBN 978-0-367-38774-7. [Google Scholar]
- Roy, S.S.; Pal, R. Microalgae in aquaculture: A review with special references to nutritional value and fish dietetics. Proc. Zool. Soc. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Bae, A.Y.; Donaldson, E.; Sitek, A.J.; Fitzgerald, D.S.; Edelson, O.F. Towards sustainable aquafeeds: Evaluating substitution of fishmeal with lipid-extracted microalgal co-product (Nannochloropsis oculata) in diets of juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0201315. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.; Zhao, C.; Zhu, J.; Hu, J.; Dong, X.; Sun, L. Dietary soybean meal affects intestinal homoeostasis by altering the microbiota, morphology and inflammatory cytokine gene expression in northern snakehead. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yaghoubi, M.; Mozanzadeh, M.T.; Marammazi, J.G.; Safari, O.; Gisbert, E. Dietary replacement of fish meal by soy products (soybean meal and isolated soy protein) in silvery-black porgy juveniles (Sparidentex hasta). Aquaculture 2016, 464, 50–59. [Google Scholar] [CrossRef]
- Perera, E.; Simó-Mirabet, P.; Shin, H.S.; Rosell-Moll, E.; Naya-Catalá, F.; Heras, V.D.L.; Martos-Sitcha, J.A.; Karalazos, V.; Armero, E.; Arizcun, M.; et al. Selection for growth is associated in gilthead sea bream (Sparus aurata) with diet flexibility, changes in growth patterns and higher intestine plasticity. Aquaculture 2019, 507, 349–360. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Bakke, A.M.; Chikwati, E.M.; Venold, F.F.; Sahlmann, C.; Holm, H.; Penn, M.H.; Oropeza-Moe, M.; Krogdahl, Å. Bile enhances glucose uptake, reduces permeability, and modulates effects of lectins, trypsin inhibitors and saponins on intestinal tissue. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 168, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Marjara, I.S.; Chikwati, E.M.; Valen, E.C.; Krogdahl, A.; Bakke, A.M. Transcriptional regulation of IL-17A and other inflammatory markers during the development of soybean meal-induced enteropathy in the distal intestine of Atlantic salmon (Salmo salar L.). Cytokine 2012, 60, 186–196. [Google Scholar] [CrossRef]
- Refstie, S.; Korsøen, Ø.J.; Storebakken, T.; Baeverfjord, G.; Lein, I.; Roem, A.J. Differing nutritional responses to dietary soybean meal in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Aquaculture 2000, 190, 49–63. [Google Scholar] [CrossRef]
- Gu, M.; Bai, N.; Xu, B.; Xu, X.; Jia, Q.; Zhang, Z. Protective effect of glutamine and arginine against soybean meal-induced enteritis in the juvenile turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2017, 70, 95–105. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Z.; Dai, J.; Yang, P.; Xu, W.; Ai, Q.; Zhang, W.; Zhang, Y.; Zhang, Y.; Mai, K. Sodium butyrate supplementation in high-soybean meal diets for turbot (Scophthalmus maximus L.): Effects on inflammatory status, mucosal barriers and microbiota in the intestine. Fish Shellfish Immunol. 2019, 88, 65–75. [Google Scholar] [CrossRef]
- Rist, V.T.; Weiss, E.; Sauer, N.; Mosenthin, R.; Eklund, M. Effect of dietary protein supply originating from soybean meal or casein on the intestinal microbiota of piglets. Anaerobe 2014, 25, 72–79. [Google Scholar] [CrossRef]
- Morales-Mena, A.; Martínez-González, S.; Teague, K.D.; Graham, L.E.; Señas-Cuesta, R.; Vuong, C.N.; Lester, H.; Hernandez-Patlan, D.; Solis-Cruz, B.; Fuente-Martinez, B.; et al. Assessment of fermented soybean meal on Salmonella typhimurium infection in neonatal turkey poults. Animals 2020, 10, 1849. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhu, M.; Cao, H.; Zhang, X.; Wang, Z.; Zhang, X.; Li, X.; Hu, J.; Yang, G.; Shi, X. Impact of fermented corn–soybean meal on gene expression of immunity in the blood, level of secretory immunoglobulin A, and mucosa-associated bacterial community in the intestine of grower–finisher pigs. Front. Vet. Sci. 2020, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of fermentation in improving nutritional quality of soybean meal—A review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.-J.; Lee, C.-H.; Kim, S.W. Aspergillus oryzaeGB-107 Fermentation Improves Nutritional Quality of Food Soybeans and Feed Soybean Meals. J. Med. Food 2004, 7, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Heng, X.; Chen, H.; Lu, C.; Feng, T.; Li, K.; Gao, E. Study on synergistic fermentation of bean dregs and soybean meal by multiple strains and proteases. LWT 2022, 154, 112626. [Google Scholar] [CrossRef]
- Teleky, B.-E.; Martău, G.A.; Vodnar, D.C. Physicochemical effects of Lactobacillus plantarum and Lactobacillus casei cocultures on soy-wheat flour dough fermentation. Foods 2020, 9, 1894. [Google Scholar] [CrossRef] [PubMed]
- Hölker, U.; Lenz, J. Solid-state fermentation—Are there any biotechnological advantages? Curr. Opin. Microbiol. 2005, 8, 301–306. [Google Scholar] [CrossRef]
- Rosales, E.; Pazos, M.; Ángeles Sanromán, M. Chapter 15—Solid-state fermentation for food applications. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 319–355. ISBN 978-0-444-63990-5. [Google Scholar]
- Dos Santos, M.M.; da Rosa, A.S.; Dal’Boit, S.; Mitchell, D.A.; Krieger, N. Thermal denaturation: Is solid-state fermentation really a good technology for the production of enzymes? Bioresour. Technol. 2004, 93, 261–268. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Mitchell, D. New developments in solid state fermentation: I-bioprocesses and products. Process. Biochem. 2000, 35, 1153–1169. [Google Scholar] [CrossRef]
- Martău, G.-A.; Unger, P.; Schneider, R.; Venus, J.; Vodnar, D.C.; López-Gómez, J.P. Integration of solid state and submerged fermentations for the valorization of organic municipal solid waste. J. Fungi 2021, 7, 766. [Google Scholar] [CrossRef]
- Weng, T.M.; Chen, M.T. Changes of protein in natto (a fermented soybean food) affected by fermenting time. Food Sci. Technol. Res. 2010, 16, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, W.; Aoyagi, A. History of Nattō and Its Relatives (1405–2012): Extensively Annotated Bibliography and Sourcebook; Soyinfo Center: Lafayette, CA, USA, 2012; ISBN 978-1-928914-42-6. [Google Scholar]
- Esaki, H.; Onozaki, H.; Osawa, T. Antioxidative activity of fermented soybean products. In Food Phytochemicals for Cancer Prevention I; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1993; Volume 546, pp. 353–360. ISBN 978-0-8412-2768-2. [Google Scholar]
- Sella, S.R.B.R.; Bueno, T.; de Oliveira, A.A.B.; Karp, S.G.; Soccol, C.R. Bacillus subtilis natto as a potential probiotic in animal nutrition. Crit. Rev. Biotechnol. 2021, 41, 355–369. [Google Scholar] [CrossRef]
- Ju, S.; Cao, Z.; Wong, C.; Liu, Y.; Foda, M.F.; Zhang, Z.; Li, J. Isolation and optimal fermentation condition of the Bacillus subtilis subsp. natto strain WTC016 for nattokinase production. Fermentation 2019, 5, 92. [Google Scholar] [CrossRef] [Green Version]
- Berikten, D.; Kivanç, M. Optimization of solid-state fermentation for phytase production by Thermomyces lanuginosus using response surface methodology. Prep. Biochem. Biotechnol. 2014, 44, 834–848. [Google Scholar] [CrossRef]
- Xiong, Y.-H.; Liu, J.-Z.; Song, H.-Y.; Ji, L.-N. Enhanced production of extracellular ribonuclease from Aspergillus niger by optimization of culture conditions using response surface methodology. Biochem. Eng. J. 2004, 21, 27–32. [Google Scholar] [CrossRef]
- Xu, L.; Yu, J.-Q.; Wang, X.-Y.; Xu, N.; Liu, J.-L. Microwave extraction optimization using the response surface methodology of Fructus Meliae toosendan polysaccharides and its antioxidant activity. Int. J. Biol. Macromol. 2018, 118, 1501–1510. [Google Scholar] [CrossRef]
- Sridhar, R.; Sivakumar, V.; Immanuel, V.P.; Maran, J.P. Development of model for treatment of pulp and paper industry bleaching effluent using response surface methodology. Environ. Prog. Sustain. Energy 2012, 31, 558–565. [Google Scholar] [CrossRef]
- Maran, J.P.; Manikandan, S. Response surface modeling and optimization of process parameters for aqueous extraction of pigments from prickly pear (Opuntia ficus-indica) fruit. Dye. Pigment. 2012, 95, 465–472. [Google Scholar] [CrossRef]
- Leni, G.; Soetemans, L.; Caligiani, A.; Sforza, S.; Bastiaens, L. Degree of hydrolysis affects the techno-functional properties of lesser mealworm protein hydrolysates. Foods 2020, 9, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Schasteen, C.S.; Wu, J.; Schulz, M.G.; Parsons, C.M. Correlation of an immobilized digestive enzyme assay with poultry true amino acid digestibility for soybean meal. Poult. Sci. 2007, 86, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Latimer, G.W. Official Methods of Analysis of AOAC International, 21st ed.; AOAC International: Rockville, MD, USA, 2019; ISBN 978-0-935584-89-9. [Google Scholar]
- Thiex, N.J.; Manson, H.; Anderson, S.; Persson, J.; Bogren, E.; Bolek, G.; Budde, D.; Ellis, C.; Eriksson, S.; Field, G.; et al. Determination of crude protein in animal feed, forage, grain, and oilseeds by using block digestion with a copper catalyst and steam distillation into boric acid: Collaborative study. J. AOAC Int. 2002, 85, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Căpriţă, R.; Căpriţă, A.; Creţescu, I. Protein solubility as quality index for processed soybean. Anim. Sci. Biotechnol. 2010, 43, 375–378. [Google Scholar]
- Araba, M.; Dale, N. Evaluation of protein solubility as an indicator of overprocessing soybean meal. Poult. Sci. 1990, 69, 76–83. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Governa, P.; Cusi, M.G.; Borgonetti, V.; Sforcin, J.M.; Terrosi, C.; Baini, G.; Miraldi, E.; Biagi, M. Beyond the biological effect of a chemically characterized poplar propolis: Antibacterial and antiviral activity and comparison with flurbiprofen in cytokines release by LPS-stimulated human mononuclear cells. Biomedicines 2019, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Sun-Waterhouse, D.; Zhao, M.; Zhao, Q.; Qiu, C.; Su, G. Effects of solid-state fermentation and proteolytic hydrolysis on defatted soybean meal. LWT 2018, 97, 496–502. [Google Scholar] [CrossRef]
- Teshima, S.-I.; Kanazawa, A.; Yamashita, M. Dietary value of several proteins and supplemental amino acids for larvae of the prawn Penaeus japonicus. Aquaculture 1986, 51, 225–235. [Google Scholar] [CrossRef]
- Kader, A.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Bulbul, M. Supplemental effects of some crude ingredients in improving nutritive values of low fishmeal diets for red sea bream, Pagrus Major. Aquaculture 2010, 308, 136–144. [Google Scholar] [CrossRef]
- Chen, J.; Wedekind, K.; Escobar, J.; Vazquez-Añón, M. Trypsin inhibitor and urease activity of soybean meal products from different countries and impact of trypsin inhibitor on ileal amino acid digestibility in pig. J. Am. Oil Chem. Soc. 2020, 97, 1151–1163. [Google Scholar] [CrossRef]
- Hei, W.; Li, Z.; Ma, X.; He, P. Determination of beta-conglycinin in soybean and soybean products using a sandwich enzyme-linked immunosorbent assay. Anal. Chim. Acta 2012, 734, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Pusztai, A.; Grant, G. Assessment of lectin inactivation by heat and digestion. In Lectin Methods and Protocols; Rhodes, J.M., Milton, J.D., Eds.; Methods in Molecular MedicineTM; Humana Press: Totowa, NJ, USA, 1998; pp. 505–514. ISBN 978-1-59259-593-8. [Google Scholar]
- Barampama, Z.; Simard, R.E. Nutrient composition, protein quality and antinutritional factors of some varieties of dry beans (Phaseolus vulgaris) grown in Burundi. Food Chem. 1993, 47, 159–167. [Google Scholar] [CrossRef]
- Kvasnicka, F.; Ahmadová-Vavrousová, R.; Frias, J.; Price, K.R.; Kadlec, P. A Rapid HPLC method for the determination of raffinose family of oligosaccharides in pea seeds. J. Liq. Chromatogr. Relat. Technol. 1996, 19, 135–147. [Google Scholar] [CrossRef]
- Reichwald, K.; Hatzack, F. Application of a modified Haug and Lantzsch method for the rapid and accurate photometrical phytate determination in soybean, wheat, and maize meals. J. Agric. Food Chem. 2008, 56, 2888–2891. [Google Scholar] [CrossRef] [PubMed]
- Shiu, Y.-L.; Wong, S.-L.; Guei, W.-C.; Shin, Y.-C.; Liu, C.-H. Increase in the plant protein ratio in the diet of white shrimp, Litopenaeus vannamei (Boone), using Bacillus subtilis E20-fermented soybean meal as a replacement. Aquac. Res. 2015, 46, 382–394. [Google Scholar] [CrossRef]
- Lenth, R.V. Response-surface methods in R, using rsm. J. Stat. Softw. 2020, 32, 1–17. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Zhang, J.; Xu, C.; Chen, X.; Ruan, X.; Zhang, Y.; Xu, H.; Guo, Y.; Xu, J.; Lv, P.; Wang, Z. Engineered Bacillus subtilis harbouring gene of d-tagatose 3-epimerase for the bioconversion of d-fructose into d-psicose through fermentation. Enzym. Microb. Technol. 2020, 136, 109531. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Feng, J.-T.; Zhang, Q.; Zhang, X. Optimization of fermentation condition for antibiotic production by Xenorhabdus nematophila with response surface methodology. J. Appl. Microbiol. 2008, 104, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Delabona, P.D.S.; Pirota, R.D.P.B.; Codima, C.A.; Tremacoldi, C.R.; Rodrigues, A.; Farinas, C.S. Effect of initial moisture content on two Amazon rainforest Aspergillus strains cultivated on agro-industrial residues: Biomass-degrading enzymes production and characterization. Ind. Crop. Prod. 2013, 42, 236–242. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Reinitz, G. Protein Dispersibility Index (PDI) as a quality control measure for soy flour used in brown trout starter feed. Progress. Fish-Cult. 1984, 46, 161–164. [Google Scholar] [CrossRef]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1988, 37, 837–841. [Google Scholar] [CrossRef]
- Moktan, B.; Saha, J.; Sarkar, P.K. Antioxidant activities of soybean as affected by Bacillus-fermentation to kinema. Food Res. Int. 2008, 41, 586–593. [Google Scholar] [CrossRef]
- Bhanja, T.; Kumari, A.; Banerjee, R. Enrichment of phenolics and free radical scavenging property of wheat koji prepared with two filamentous fungi. Bioresour. Technol. 2009, 100, 2861–2866. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Y.; Ferrari, E.; Henner, D.J. Cloning of the neutral protease gene of Bacillus subtilis and the use of the cloned gene to create an in vitro-derived deletion mutation. J. Bacteriol. 1984, 160, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.-C.; Ling, M.-Y. Biochemical changes in soy sauce prepared with extruded and traditional raw materials. Food Res. Int. 1998, 31, 487–492. [Google Scholar] [CrossRef]
- Klapper, B.F.; Jameson, D.M.; Mayer, R.M. Factors affecting the synthesis and release of the extracellular protease of Aspergillus oryzae NRRL. Biochim. Biophys. Acta BBA Gen. Subj. 1973, 304, 513–519. [Google Scholar] [CrossRef]
- Sarkar, P.; Jones, L.; Craven, G.; Somerset, S.; Palmer, C. Amino acid profiles of kinema, a soybean-fermented food. Food Chem. 1997, 59, 69–75. [Google Scholar] [CrossRef]
- Han, X.; Nagano, H.; Phromraksa, P.; Tsuji, M.; Shimoyamada, M.; Kasuya, S.; Suzuki, T.; Khamboonruang, C. Hydrolysis of soybean 7S and 11S globulins using Bacillus subtilis. Food Sci. Technol. Res. 2012, 18, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Bone, R. Pathological Effects of Soybean Anti-Nutritional Factors on Summer Flounder (Paralichthys dentatus) Tissues. Master’s Thesis, University of Rhode Island, Kingston, RI, USA, 2013. [Google Scholar]
- Escaffre, A.M.; Zambonino-Infante, J.L.; Cahu, C.L.; Mambrini, M.; Bergot, P.; Kaushik, S. Nutritional value of soy protein concentrate for larvae of common carp (Cyprinus carpio) based on growth performance and digestive enzyme activities. Aquaculture 1997, 153, 63–80. [Google Scholar] [CrossRef]
- Peumans, W.J.; Van Damme, E. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef]
- Buttle, L.; Burrells, A.; Good, J.; Williams, P.; Southgate, P. The binding of soybean agglutinin (SBA) to the intestinal epithelium of Atlantic salmon, Salmo salar and Rainbow trout, Oncorhynchus mykiss, fed high levels of soybean meal. Vet. Immunol. Immunopathol. 2001, 80, 237–244. [Google Scholar] [CrossRef]
- He, L.; Han, M.; Qiao, S.; He, P.; Li, D.; Li, N.; Ma, X. Soybean antigen proteins and their intestinal sensitization activities. Curr. Protein Pept. Sci. 2015, 16, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Storebakken, T.; Shearer, K.; Roem, A. Availability of protein, phosphorus and other elements in fish meal, soy-protein concentrate and phytase-treated soy-protein-concentrate-based diets to Atlantic salmon, Salmo salar. Aquaculture 1998, 161, 365–379. [Google Scholar] [CrossRef]
- Carter, C.; Hauler, R. Fish meal replacement by plant meals in extruded feeds for Atlantic salmon, Salmo salar L. Aquaculture 2000, 185, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, N.; Ray, A.K. Effect of fermentation on the nutritive value of sesame seed meal in the diets for rohu, Labeo rohita (Hamilton), fingerlings. Aquac. Nutr. 1999, 5, 229–236. [Google Scholar] [CrossRef]
- Zhao, S.; Hu, N.; Huang, J.; Liang, Y.; Zhao, B. High-yield spore production from Bacillus licheniformis by solid state fermentation. Biotechnol. Lett. 2008, 30, 295–297. [Google Scholar] [CrossRef] [PubMed]
- Teng, D.; Gao, M.; Yang, Y.; Liu, B.; Tian, Z.; Wang, J. Bio-modification of soybean meal with Bacillus subtilis or Aspergillus oryzae. Biocatal. Agric. Biotechnol. 2012, 1, 32–38. [Google Scholar] [CrossRef]
- Msadek, T. When the going gets tough: Survival strategies and environmental signaling networks in Bacillus subtilis. Trends Microbiol. 1999, 7, 201–207. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Strain | Inoculation Quantity (log CFU/kg) | Temperature (°C) | Time (h) | Water–Material Ratio | Layer Thickness (cm) | Initial pH |
|---|---|---|---|---|---|---|
| B.s. natto | 10.0 | 40.0 | 48.0 | 1.0 | 2.0 | 7.5 |
| Code | Independent Variables | Levels | ||||
|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | ||
| A | Temperature (°C) | 31.00 | 36.00 | 41.00 | 46.00 | 51.00 |
| B | Time (h) | 54.00 | 60.00 | 66.00 | 72.00 | 78.00 |
| C | Water–material ratio | 0.90 | 1.00 | 1.10 | 1.20 | 1.30 |
| D | Layer thickness (cm) | 1.60 | 1.80 | 2.00 | 2.20 | 2.40 |
| Runs | A | B | C | D | Response 1: CP (%) | Response 2: DH (%) |
|---|---|---|---|---|---|---|
| 1 | 36.00 | 72.00 | 1.20 | 1.80 | 54.86 | 15.53 |
| 2 | 36.00 | 60.00 | 1.00 | 1.80 | 55.24 | 15.73 |
| 3 | 41.00 | 66.00 | 1.10 | 2.00 | 55.59 | 15.94 |
| 4 | 41.00 | 66.00 | 1.10 | 2.40 | 53.64 | 15.56 |
| 5 | 41.00 | 66.00 | 1.10 | 2.00 | 55.64 | 15.93 |
| 6 | 46.00 | 60.00 | 1.20 | 2.20 | 55.49 | 15.86 |
| 7 | 46.00 | 72.00 | 1.20 | 1.80 | 54.36 | 15.53 |
| 8 | 46.00 | 72.00 | 1.20 | 2.20 | 54.91 | 15.87 |
| 9 | 51.00 | 66.00 | 1.10 | 2.00 | 55.15 | 15.89 |
| 10 | 41.00 | 54.00 | 1.10 | 2.00 | 55.53 | 15.76 |
| 11 | 36.00 | 72.00 | 1.20 | 2.20 | 54.81 | 15.67 |
| 12 | 41.00 | 66.00 | 1.10 | 1.60 | 53.99 | 15.36 |
| 13 | 31.00 | 66.00 | 1.10 | 2.00 | 55.05 | 15.62 |
| 14 | 41.00 | 66.00 | 0.90 | 2.00 | 55.16 | 15.87 |
| 15 | 41.00 | 66.00 | 1.10 | 2.00 | 55.83 | 15.92 |
| 16 | 46.00 | 60.00 | 1.00 | 2.20 | 55.03 | 15.85 |
| 17 | 36.00 | 72.00 | 1.00 | 2.20 | 54.25 | 15.65 |
| 18 | 46.00 | 72.00 | 1.00 | 2.20 | 54.53 | 15.85 |
| 19 | 46.00 | 60.00 | 1.00 | 1.80 | 55.34 | 15.80 |
| 20 | 36.00 | 72.00 | 1.00 | 1.80 | 55.15 | 15.71 |
| 21 | 41.00 | 66.00 | 1.10 | 2.00 | 55.75 | 15.94 |
| 22 | 36.00 | 60.00 | 1.00 | 2.20 | 54.40 | 15.56 |
| 23 | 41.00 | 78.00 | 1.10 | 2.00 | 54.87 | 15.78 |
| 24 | 46.00 | 60.00 | 1.20 | 1.80 | 54.87 | 15.59 |
| 25 | 46.00 | 72.00 | 1.00 | 1.80 | 54.84 | 15.72 |
| 26 | 41.00 | 66.00 | 1.10 | 2.00 | 55.75 | 15.93 |
| 27 | 36.00 | 60.00 | 1.20 | 2.20 | 55.11 | 15.56 |
| 28 | 41.00 | 66.00 | 1.10 | 2.00 | 55.65 | 15.95 |
| 29 | 41.00 | 66.00 | 1.30 | 2.00 | 55.30 | 15.68 |
| 30 | 36.00 | 60.00 | 1.20 | 1.80 | 55.01 | 15.50 |
| ANOVA for Quadratic Model | ANOVA for Reduced Quadratic Model | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | Sum of Squares | df | Mean Square | F-value | p-value | Source | Sum of Squares | df | Mean Square | F-value | p-value |
| Model | 0.7664 | 14 | 0.0547 | 972.72 | <0.0001 | Model | 0.7664 | 13 | 0.059 | 1111.15 | <0.0001 |
| A | 0.1185 | 1 | 0.1185 | 2106.2 | <0.0001 | A | 0.1185 | 1 | 0.1185 | 2234.09 | <0.0001 |
| B | 0.0005 | 1 | 0.0005 | 8.65 | 0.0101 | B | 0.0005 | 1 | 0.0005 | 9.18 | 0.008 |
| C | 0.0534 | 1 | 0.0534 | 948.5 | <0.0001 | C | 0.0534 | 1 | 0.0534 | 1006.1 | <0.0001 |
| D | 0.053 | 1 | 0.053 | 942.48 | <0.0001 | D | 0.053 | 1 | 0.053 | 999.71 | <0.0001 |
| AB | 0.0073 | 1 | 0.0073 | 129.66 | <0.0001 | AB | 0.0073 | 1 | 0.0073 | 137.53 | <0.0001 |
| AC | 4.73 × 10−6 | 1 | 4.73 × 10−6 | 0.0841 | 0.7758 | ||||||
| AD | 0.0417 | 1 | 0.0417 | 741.42 | <0.0001 | AD | 0.0417 | 1 | 0.0417 | 786.44 | <0.0001 |
| BC | 0.0009 | 1 | 0.0009 | 16.83 | 0.0009 | BC | 0.0009 | 1 | 0.0009 | 17.85 | 0.0006 |
| BD | 0.0072 | 1 | 0.0072 | 128.3 | <0.0001 | BD | 0.0072 | 1 | 0.0072 | 136.09 | <0.0001 |
| CD | 0.0452 | 1 | 0.0452 | 802.89 | <0.0001 | CD | 0.0452 | 1 | 0.0452 | 851.65 | <0.0001 |
| A2 | 0.057 | 1 | 0.057 | 1013.04 | <0.0001 | A2 | 0.057 | 1 | 0.057 | 1074.56 | <0.0001 |
| B2 | 0.05 | 1 | 0.05 | 887.75 | <0.0001 | B2 | 0.05 | 1 | 0.05 | 941.66 | <0.0001 |
| C2 | 0.0479 | 1 | 0.0479 | 850.7 | <0.0001 | C2 | 0.0479 | 1 | 0.0479 | 902.36 | <0.0001 |
| D2 | 0.392 | 1 | 0.392 | 6964.4 | <0.0001 | D2 | 0.392 | 1 | 0.392 | 7387.3 | <0.0001 |
| Residual | 0.0008 | 15 | 0.0001 | Residual | 0.0008 | 16 | 0.0001 | ||||
| Lack of Fit | 0.0004 | 10 | 0 | 0.478 | 0.85 | Lack of Fit | 0.0004 | 11 | 0 | 0.4395 | 0.8811 |
| Pure Error | 0.0004 | 5 | 0.0001 | Pure Error | 0.0004 | 5 | 0.0001 | ||||
| Cor Total | 0.7673 | 29 | Cor Total | 0.7673 | 29 | ||||||
| Credibility analysis of the regression equations for Quadratic model | Credibility analysis of the regression equations for Reduced Quadratic model | ||||||||||
| Std. Dev. | 0.0075 | R2 | 0.9989 | Std. Dev. | 0.0073 | R2 | 0.9989 | ||||
| Mean | 15.74 | Adjusted R2 | 0.9979 | Mean | 15.74 | Adjusted R2 | 0.998 | ||||
| C.V. % | 0.0477 | Predicted R2 | 0.9961 | C.V. % | 0.0463 | Predicted R2 | 0.9967 | ||||
| ANOVA for Quadratic Model | ANOVA for Reduced Quadratic Model | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | Sum of Squares | df | Mean Square | F-value | p-value | Source | Sum of Squares | df | Mean Square | F-value | p-value |
| Model | 8.29 | 14 | 0.5924 | 148.94 | <0.0001 | Model | 8.28 | 12 | 0.6903 | 168.99 | <0.0001 |
| A | 0.0214 | 1 | 0.0214 | 5.39 | 0.0348 | A | 0.0214 | 1 | 0.0214 | 5.24 | 0.0351 |
| B | 0.706 | 1 | 0.706 | 177.52 | <0.0001 | B | 0.706 | 1 | 0.706 | 172.85 | <0.0001 |
| C | 0.0358 | 1 | 0.0358 | 9 | 0.009 | C | 0.0358 | 1 | 0.0358 | 8.76 | 0.0088 |
| D | 0.1415 | 1 | 0.1415 | 35.58 | <0.0001 | D | 0.1415 | 1 | 0.1415 | 34.64 | <0.0001 |
| AB | 0.1222 | 1 | 0.1222 | 30.71 | <0.0001 | AB | 0.1222 | 1 | 0.1222 | 29.91 | <0.0001 |
| AC | 0.0469 | 1 | 0.0469 | 11.8 | 0.0037 | AC | 0.0469 | 1 | 0.0469 | 11.49 | 0.0035 |
| AD | 0.3134 | 1 | 0.3134 | 78.81 | <0.0001 | AD | 0.3134 | 1 | 0.3134 | 76.74 | <0.0001 |
| BC | 0.0054 | 1 | 0.0054 | 1.35 | 0.2633 | ||||||
| BD | 0.0044 | 1 | 0.0044 | 1.11 | 0.3091 | ||||||
| CD | 0.8085 | 1 | 0.8085 | 203.28 | <0.0001 | CD | 0.8085 | 1 | 0.8085 | 197.93 | <0.0001 |
| A2 | 0.5457 | 1 | 0.5457 | 137.22 | <0.0001 | A2 | 0.5457 | 1 | 0.5457 | 133.61 | <0.0001 |
| B2 | 0.3734 | 1 | 0.3734 | 93.89 | <0.0001 | B2 | 0.3734 | 1 | 0.3734 | 91.42 | <0.0001 |
| C2 | 0.3226 | 1 | 0.3226 | 81.12 | <0.0001 | C2 | 0.3226 | 1 | 0.3226 | 78.99 | <0.0001 |
| D2 | 5.86 | 1 | 5.86 | 1472.95 | <0.0001 | D2 | 5.86 | 1 | 5.86 | 1434.18 | <0.0001 |
| Residual | 0.0597 | 15 | 0.004 | Residual | 0.0694 | 17 | 0.0041 | ||||
| Lack of Fit | 0.0198 | 10 | 0.002 | 0.2477 | 0.9711 | Lack of Fit | 0.0295 | 12 | 0.0025 | 0.3086 | 0.9558 |
| Pure Error | 0.0399 | 5 | 0.008 | Pure Error | 0.0399 | 5 | 0.008 | ||||
| Cor Total | 8.35 | 29 | Cor Total | 8.35 | 29 | ||||||
| Credibility analysis of the regression equations for Quadratic model | Credibility analysis of the regression equations for Reduced Quadratic model | ||||||||||
| Std. Dev. | 0.0631 | R2 | 0.9929 | Std. Dev. | 0.0639 | R2 | 0.9917 | ||||
| Mean | 55.04 | Adjusted R2 | 0.9862 | Mean | 55.04 | Adjusted R2 | 0.9858 | ||||
| C.V. % | 0.1146 | Predicted R2 | 0.9795 | C.V. % | 0.1161 | Predicted R2 | 0.9781 | ||||
| Response | Predicted Value | Observed Value | p-Value | 95% Confidence Interval | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| CP (%) | 55.76 | 55.71 ± 0.17 | 0.673 | 55.30 | 56.12 |
| DH (%) | 15.96 | 15.85 ± 0.08 | 0.137 | 15.66 | 16.04 |
| Items | SBM | FSM | p-Value | Change (%) |
|---|---|---|---|---|
| Dry matter (%) | 90.56 ± 0.09 | 91.85 ± 0.08 | <0.001 | 1.4 |
| CP (%) | 50.72 ± 0.30 | 55.71 ± 0.17 | <0.001 | 9.8 |
| CL (%) | 2.08 ± 0.07 | 1.77 ± 0.05 | 0.004 | −14.9 |
| CF (%) | 5.80 ± 0.21 | 4.98 ± 0.33 | 0.022 | −14.1 |
| CA (%) | 6.33 ± 0.12 | 6.68 ± 0.05 | 0.010 | 5.4 |
| PDI (% of CP) | 31.20 ± 0.70 | 21.90 ± 0.79 | <0.001 | −29.8 |
| KPS (% of CP) | 80.43 ± 1.96 | 94.47 ± 0.81 | <0.001 | 17.5 |
| DH (%) | 5.81 ± 0.18 | 16.01 ± 0.11 | <0.001 | 175.3 |
| Total phenol (mg·g−1) | 3.25 ± 0.18 | 13.32 ± 1.13 | <0.001 | 309.4 |
| Flavonoids (mg·g−1) | 1.36 ± 0.18 | 4.49 ± 0.27 | <0.001 | 231.0 |
| Items | SBM | FSM | p-Value | Change (%) |
|---|---|---|---|---|
| EAA | ||||
| Lysine | 2.93 ± 0.12 | 3.18 ± 0.21 | 0.149 | 8.5 |
| Methionine | 0.70 ± 0.06 | 0.85 ± 0.04 | 0.028 | 20.9 |
| Isoleucine | 2.37 ± 0.08 | 2.56 ± 0.06 | 0.025 | 8.0 |
| Leucine | 3.87 ± 0.15 | 4.25 ± 0.25 | 0.086 | 9.8 |
| Valine | 2.10 ± 0.11 | 2.53 ± 0.13 | 0.012 | 20.5 |
| Arginine | 3.54 ± 0.25 | 3.90 ± 0.05 | 0.070 | 10.2 |
| Threonine | 1.91 ± 0.13 | 2.35 ± 0.20 | 0.032 | 23.2 |
| Tryptophan | 0.65 ± 0.04 | 0.75 ± 0.03 | 0.029 | 14.9 |
| Histidine | 1.45 ± 0.10 | 1.59 ± 0.07 | 0.103 | 9.9 |
| Phenylalanine | 2.49 ± 0.15 | 2.74 ± 0.12 | 0.086 | 10.1 |
| NEAA | ||||
| Cystine | 0.84 ± 0.07 | 0.92 ± 0.02 | 0.157 | 9.1 |
| Tyrosine | 1.84 ± 0.10 | 2.01 ± 0.10 | 0.098 | 9.2 |
| Serine | 2.17 ± 0.11 | 2.55 ± 0.13 | 0.018 | 17.7 |
| Glutamic acid | 8.17 ± 0.62 | 9.18 ± 0.09 | 0.049 | 12.3 |
| Proline | 2.51 ± 0.14 | 2.85 ± 0.05 | 0.016 | 13.7 |
| Glycine | 2.00 ± 0.17 | 2.17 ± 0.18 | 0.297 | 8.3 |
| Alanine | 2.11 ± 0.11 | 2.34 ± 0.06 | 0.033 | 11.2 |
| Aspartic acid | 5.70 ± 0.20 | 6.05 ± 0.25 | 0.128 | 6.1 |
| EAA | 22.01 ± 0.41 | 24.70 ± 0.20 | <0.001 | 12.2 |
| NEAA | 25.34 ± 0.54 | 28.07 ± 0.28 | 0.001 | 10.8 |
| Total Amino Acids | 47.35 ± 0.68 | 52.77 ± 0.11 | <0.001 | 11.4 |
| Total free amino acid | 0.46 ± 0.00 | 6.33 ± 0.09 | <0.001 | 1276.1 |
| Items | SBM | FSM | p-Value | Change (%) |
|---|---|---|---|---|
| Trypsin inhibitor | 3.50 ± 0.10 | 1.62 ± 0.09 | <0.001 | −53.67 |
| Glycinin | 79.20 ± 0.44 | 17.63 ± 1.68 | <0.001 | −77.74 |
| β-Conglycinin | 106.38 ± 4.15 | 30.17 ± 6.91 | <0.001 | −71.64 |
| Lectins | 3.35 ± 0.20 | 0.02 ± 0.01 | <0.001 | −99.40 |
| Raffinose | 18.43 ± 1.05 | 1.67 ± 0.23 | <0.001 | −90.94 |
| Stachyose | 11.92 ± 1.55 | 0.55 ± 0.08 | <0.001 | −95.38 |
| Phytic acid | 1.73 ± 0.11 | 0.13 ± 0.01 | <0.001 | −92.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Dossou, S.; Wang, W.; Zhang, X.; Shadrack, R.S.; Mzengereza, K.; Zhu, K.; et al. Optimization of Soybean Meal Fermentation for Aqua-Feed with Bacillus subtilis natto Using the Response Surface Methodology. Fermentation 2021, 7, 306. https://doi.org/10.3390/fermentation7040306
Zhang Y, Ishikawa M, Koshio S, Yokoyama S, Dossou S, Wang W, Zhang X, Shadrack RS, Mzengereza K, Zhu K, et al. Optimization of Soybean Meal Fermentation for Aqua-Feed with Bacillus subtilis natto Using the Response Surface Methodology. Fermentation. 2021; 7(4):306. https://doi.org/10.3390/fermentation7040306
Chicago/Turabian StyleZhang, Yukun, Manabu Ishikawa, Shunsuke Koshio, Saichiro Yokoyama, Serge Dossou, Weilong Wang, Xiaoxiao Zhang, Ronick Spenly Shadrack, Kumbukani Mzengereza, Kehua Zhu, and et al. 2021. "Optimization of Soybean Meal Fermentation for Aqua-Feed with Bacillus subtilis natto Using the Response Surface Methodology" Fermentation 7, no. 4: 306. https://doi.org/10.3390/fermentation7040306