
Kinetics of In Vitro Gas Production and Fitting Mathematical Models of Corn Silage

 , , , and
, , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals, Experimental Design and Treatments
2.2. Chemical Analysis of Silage
2.3. In Vitro Gas Production Data
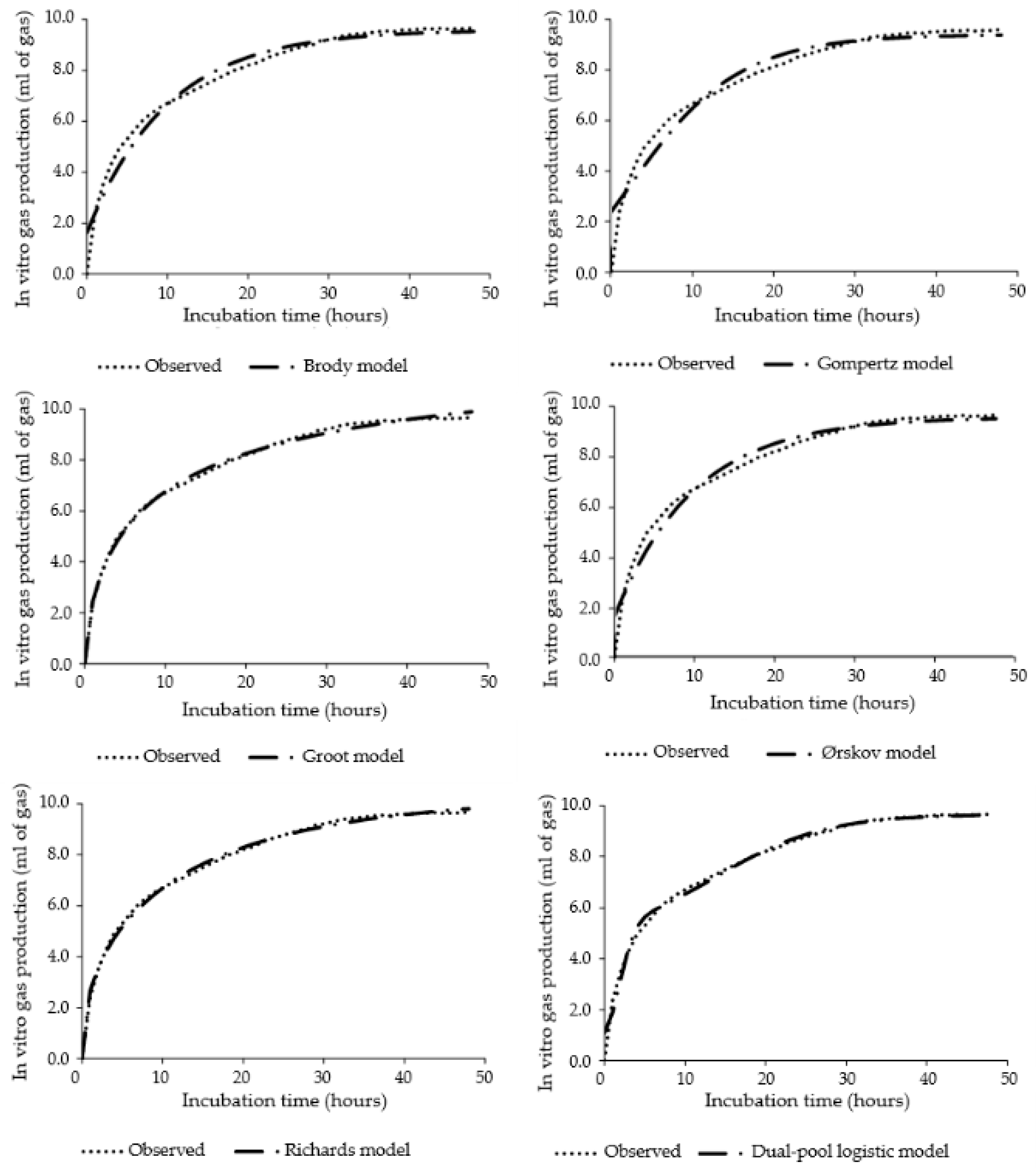
2.4. Models and Curve-Fitting
2.5. Statistical Analysis
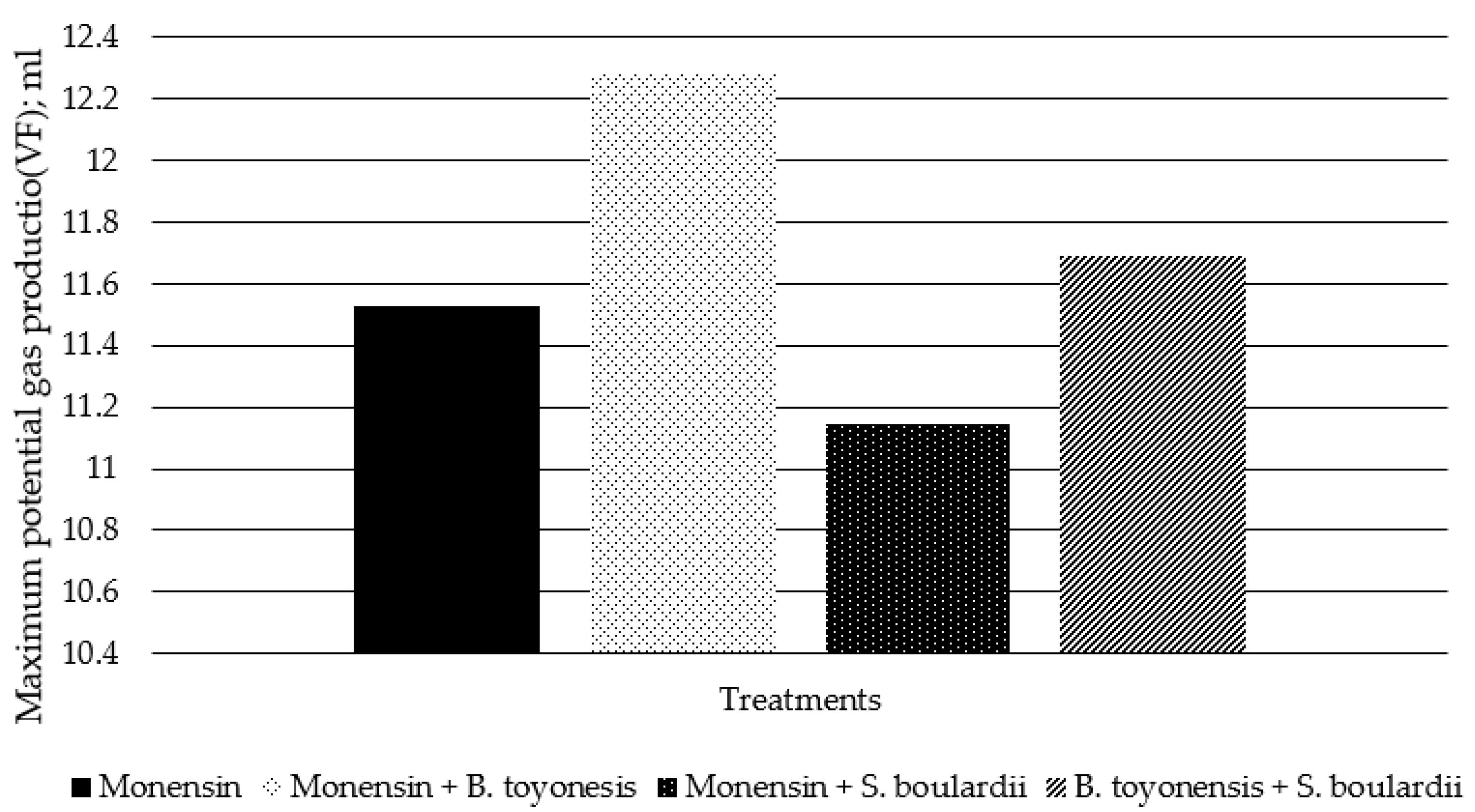
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brody, S. Bioenergetics and Growth; with Special Reference to the Efficiency Complex in Domestic Animals; Reinhold: Oxford, UK, 1945. [Google Scholar]
- Von Bertalanffy, L. Quantitative Laws in Metabolism and Growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Laird, A.K. Dynamics of relative growth. Growth 1965, 29, 249–263. [Google Scholar]
- Savageau, M.A. Growth equations: A general equation and a survey of special cases. Math. Biosci. 1980, 48, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Groot, J.C.J.; Cone, J.W.; Williams, B.A.; Debersaques, F.M.A.; Lantinga, E.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Orskov, E.R.; Mcdonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Schofield, P.; Pitt, R.E.; Pell, A.N. Kinetics of fiber digestion from in vitro gas production. J. Anim. Sci. 1994, 72, 2980–2991. [Google Scholar] [CrossRef]
- Olivo, P.M.; Santos, G.T.; Ítavo, L.C.V.; Silva Junior, R.C.; Leal, E.S.; Prado, R.M. Assessing the nutritional value of agroindustrial co-products and feed through chemical composition, in vitro digestibility, and gas production technique. Acta Sci. Anim. Sci. 2017, 39, 289–295. [Google Scholar] [CrossRef]
- Diaz, T.G.; Branco, A.F.; Ítavo, L.C.V.; Santos, G.T.; Carvalho, S.T.; Teodoro, A.L.; Oliveira, R.L. In vitro gas production kinetics and digestibility in ruminant diets with diferente levels of cashew nut shell liquid. Semin. Cienc. Agrar. 2018, 39, 1669–1682. [Google Scholar] [CrossRef]
- Souza, A.D.V.; Ítavo, L.C.V.; Favaro, S.P.; Ítavo, C.C.B.F.; Petit, H.V.; Dias, A.M.; Morais, M.G.; Reis, F.A.; Roscoe, R. Thermal decomposition, chemical composition, in vitro digestibility and gas production and in situ degradability of oilseed residues from the biofuel industry. Anim. Sci. J. 2018, 89, 79–87. [Google Scholar] [CrossRef]
- Leal, E.S.; Ítavo, L.C.V.; Valle, C.B.; Ítavo, C.C.B.F.; Dias, A.M.; Difante, G.S.; Barbosa-Ferreira, M.; Nonato, L.M.; Melo, G.K.A.; Gurgel, A.L.C. AInfluence of protodioscin content on digestibility and in vitro degradation kinetics in Urochloa brizantha cultivars. Crop Pasture Sci. 2020, 71, 278–284. [Google Scholar] [CrossRef]
- Santana, J.C.S.; Morais, J.A.S.; Difante, G.S.; Ítavo, L.C.V.; Gurgel, A.L.C.; Oliveira, V.S.; Rodrigues, M.J.S.T. In vitro digestion characteristics of various combinations of elephant grass hay, Gliricidia hay or silage, soybean meal and corn meal in rations for sheep. Trop. Grassl. 2020, 8, 147–152. [Google Scholar] [CrossRef]
- Gurgel, A.L.C.; Morais, J.A.S.; Santana, J.C.S.; Difante, G.S.; Emerenciano Neto, J.V.; Ítavo, L.C.V.; Ítavo, C.C.B.F.; Oliveira, V.S.; Rodrigues, M.J.S.T. Mathematical models to adjust the parameters of in vitro cumulative gas production of diets containing preserved Gliricidia. Ciência Rural 2021, 51, e20200993. [Google Scholar] [CrossRef]
- Weiss, C.P.; Beck, P.A.; Gadberry, M.S.; Richeson, J.T.; Wilson, B.K.; Robinson, C.A.; Zhao, J.; Hess, T.; Hubbell, D. Effects of intake of monensin during the stocker phase and subsequent finishing phase on performance and carcass characteristics of finishing beef steers. Appl. Anim. Sci. 2020, 36, 668–676. [Google Scholar] [CrossRef]
- Tseu, R.J.; Perna Junior, F.; Carvalho, R.F.; Sene, G.A.; Tropaldi, C.B.; Peres, A.H.; Rodrigues, P.H.M. Effect of tannins and monensin on feeding behaviour, feed intake, digestive parameters and microbial efficiency of nellore cows. Ital. J. Anim. Sci. 2020, 19, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.R.; Beck, M.R.; Gunter, S.A.; Williams, G.D.; Place, S.E.; Reuter, R.R. An energy and monensin supplement reduces methane emission intensity of stocker cattle grazing winter wheat. Appl. Anim. Sci. 2019, 35, 433–440. [Google Scholar] [CrossRef]
- İnal, F.; Gürbüz, E.; Coşkun, B.; Alataş, M.S.; Çitil, Ö.B.; Polat, E.S.; Şeker, E.; Özcan, C. The Effects of Live Yeast Culture (Saccharomyces cerevisiae) on Rumen Fermentation and Nutrient Degradability in Yearling Lambs. Kafkas Üniversitesi Vet. Fakültesi Derg. 2010, 16, 799–804. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. 2015, 30, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Young, T.R.; Ribeiro, F.R.B.; Sanchez, N.C.B.; Carroll, J.A.; Jennings, M.A.; Cribbs, J.T.; Rathmann, R.J.; Corley, J.R.; Johnson, B.J. Yeast cell wall supplementation alters the performance and health of beef heifers during the receiving period. Prof. Anim. Sci. 2017, 33, 166–175. [Google Scholar] [CrossRef]
- Diaz, T.G.; Branco, A.F.; Jacovaci, F.A.; Jobim, C.C.; Bolson, D.C.; Daniel, J.L.P. Inclusion of live yeast and mannan-oligosaccharides in high grain-based diets for sheep: Ruminal parameters, inflammatory response and rumen morphology. PLoS ONE 2018, 13, e0193313. [Google Scholar] [CrossRef] [Green Version]
- Diaz, T.G.; Branco, A.F.; Jacovaci, F.A.; Jobim, C.C.; Daniel, J.L.P.; Bueno, A.V.I.; Ribeiro, M.G. Use of live yeast and mannan-oligosaccharides in grain-based diets for cattle: Ruminal parameters, nutrient digestibility, and inflammatory response. PLoS ONE 2018, 13, e0207127. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Chagoyán, J.C.V.; Salem, A.Z.M.; Kholif, A.E.; Castañeda, J.S.M.; Camacho, L.M.; Cerrillo-Soto, M.A. Effects of Saccharomyces cerevisiae at direct addition or pre-incubation on in vitro gas production kinetics and degradability of four fibrous feeds. Ital. J. Anim. Sci. 2014, 13, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Wambui, C.; Abdulrazak, S. Effect of yeast supplementation on in vitro ruminal degradability of selected browse species from Kenya. J. Food Agric. Environ. 2010, 8, 553–557. [Google Scholar]
- Opsi, F.; Fortina, R.; Tassone, S.; Bodas, R.; López, S. Effects of inactivated and live cells of Saccharomyces cerevisiae on in vitro ruminal fermentation of diets with different forage: Concentrate ratio. J. Agric. Sci. 2012, 150, 271–283. [Google Scholar] [CrossRef] [Green Version]
- EFSA (European Food Safety Authority). Scientific Opinion on the safety and efficacy of Toyocerin® (Bacillus toyonensis) as a feed additive for chickens for fattening, weaned piglets, pigs for fattening, sows for reproduction, cattle for fattening and calves for rearing and for rabbits for fat. EFSA J. 2014, 12, 17. [Google Scholar] [CrossRef]
- Roos, T.B.; de Moraes, C.M.; Sturbelle, R.T.; Dummer, L.A.; Fischer, G.; Leite, F.P.L. Probiotics Bacillus toyonensis and Saccharomyces boulardii improve the vaccine immune response to Bovine herpesvirus type 5 in sheep. Res. Vet. Sci. 2018, 117, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Zardin, P.B.; Velho, J.P.; Jobim, C.C.; Alessio, D.R.M.; Haygert-Velho, I.M.P.; da Conceição, G.M.; Almeid, P.S.G. Chemical composition of corn silage produced by scientific studies in Brazil—A meta-analysis. Semin. Ciências Agrárias 2017, 38, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. Special Topics—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options 1. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.C.; Diaz, B.L.; Rivera, E.A.B.; Granjeiro, J.M.; Braga, L.M.G.M.; Frajblat, M.; Stephano, M.A. Guia Brasileiro de Produção, Manutenção ou Utilização de Animais em Atividades de Ensino ou Pesquisa: Fascículo 1: Introdução Geral; Ministérios da Ciência, Tecnologia e Inovação, Ed.; Concelho Nacional de Controle de Experimentação Animal: Brasilia, Brazil, 2016; ISBN 9788588063310. [Google Scholar]
- AOAC Official Methods of Analysis of AOAC International. Official Methods of Analysis of AOAC; AOAC International: Rockville, MD, USA, 2005; ISBN 0935584544. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci 1991, 10, 3583–3597. [Google Scholar] [CrossRef]
- Sniffen, C.J.; O’Connor, J.D.; Van Soest, P.J.; Fox, D.G.; Russell, J.B. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992, 70, 3562–3577. [Google Scholar] [CrossRef]
- Marten, G.C.; Barnes, R.F. Prediction of energy digestibility of forages with in vitro rumen fermentation and fungal enzyme systems. Stand. Anal. Methodol. Feed. 1979, 4, 61–71. [Google Scholar]
- Tedeschi, L.O. Assessment of the adequacy of mathematical models. Agric. Syst. 2006, 89, 225–247. [Google Scholar] [CrossRef]
- NRCS. Animal Diets and Feed Management; United States Department of Agriculture Natural Resources Conservation Service: Washington, DC, USA, 2012; pp. 1–14.
- National Research Council. Nutrient Requirements of Dairy Cattle; National Academies Press: Washington, DC, USA, 2001; ISBN 978-0-309-06997-7. [Google Scholar]
- Nematpour, A.; Eshghizadeh, H.R.; Zahedi, M. Comparing the Corn, Millet and Sorghum as Silage Crops Under Different Irrigation Regime and Nitrogen Fertilizer Levels. Int. J. Plant Prod. 2021, 15, 351–361. [Google Scholar] [CrossRef]
- Bryant, H.T.; Blaser, R.E.; Hammes, R.C.; Huber, J.T. Evaluation of Corn Silage Harvested at Two Stages of Maturity 1. Agron. J. 1966, 58, 253–255. [Google Scholar] [CrossRef]
- Colenbrander, V.F.; Martin, G. Relationships Between Stage of Maturity of the Corn Plant at Time of Harvest for Corn Silage and Chemical Composition1,2. J. Dairy Sci. 1965, 54, 533–536. [Google Scholar] [CrossRef]
- Wang, M.; Tang, S.X.; Tan, Z.L. Modeling in vitro gas production kinetics: Derivation of Logistic-Exponential (LE) equations and comparison of models. Anim. Feed Sci. Technol. 2011, 165, 137–150. [Google Scholar] [CrossRef]
- Mjoun, K. 72 Evaluation of Different Gas Production Models When Applied to Ruminants Feeds. J. Anim. Sci. 2018, 96, 37–38. [Google Scholar] [CrossRef]
- Mello, R.; Magalhães, A.L.R.; Breda, F.C.; Regazzi, A.J. Modelos para ajuste da produção de gases em silagens de girassol e milho. Pesqui. Agropecuária Bras. 2008, 43, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Elghandour, M.M.Y.; Vázquez, J.C.; Salem, A.Z.M.; Kholif, A.E.; Cipriano, M.M.; Camacho, L.M.; Márquez, O. In vitro gas and methane production of two mixed rations influenced by three different cultures of Saccharomyces cerevisiae. J. Appl. Anim. Res. 2017, 45, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Macome, F.M.; Pellikaan, W.F.; Hendriks, W.H.; Dijkstra, J.; Hatew, B.; Schonewille, J.T.; Cone, J.W. In vitro gas and methane production of silages from whole-plant corn harvested at 4 different stages of maturity and a comparison with in vivo methane production. J. Dairy Sci. 2017, 100, 8895–8905. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhao, M.; Wang, X.; Yu, Z.; Na, R. Ensiling alfalfa with whole crop corn improves the silage quality and in vitro digestibility of the silage mixtures. Grassl. Sci. 2017, 63, 211–217. [Google Scholar] [CrossRef]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Faria, B.N.; Reis, R.B.; Maurício, R.M.; Lana, A.M.Q.; Soares, S.R.V.; Saturnino, H.M.; Coelho, S.G. Efeitos da adição de propilenoglicol ou monensina à silagem de milho sobre a cinética de degradação dos carboidratos e produção cumulativa de gases in vitro. Arq. Bras. De Med. Veterinária E Zootec. 2008, 60, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Liu, Z.; Yu, Z.; Zhu, W. Monensin and nisin affect rumen fermentation and microbiota differently in vitro. Front. Microbiol. 2017, 8, 1111. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.P.; Mariezcurrena, M.D.; Mariezcurrena, M.A.; Lagunas, B.C.; Elghandour, M.M.M.Y.; Kholif, A.M.; Kholif, A.E.; Almaraz, E.M.; Salem, A.Z.M. Influence of live cells or cells extract of Saccharomyces cerevisiae on in vitro gas production of a total mixed ration. Ital. J. Anim. Sci. 2015, 14, 590–595. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | Diet |
|---|---|
| Ingredient (% DM of diet) | |
| Corn silage (Roughage) | 30.0 |
| Concentrate a | |
| Ground corn | 45.500 |
| Soybean meal | 8.517 |
| Soybean hull | 14.000 |
| Starea® | 0.817 |
| Mineral | 1.166 |
| Chemical composition of diet (g/kg DM of diet) | |
| Dry matter | 604.938 |
| Organic matter | 953.388 |
| Crude protein | 152.441 |
| Ethereal extract | 22.919 |
| Neutral detergent fiber | 381.568 |
| Acid detergent fiber | 208.071 |
| Models | Equation | N° of Parameters |
|---|---|---|
| Gompertz | P(t) = VF e(−b.e(−kt)) | 3 |
| Groot | P(t) = VF/(1 + (bk/tk)) | 3 |
| Ørskov | P(t) = VF + b(1 − e(−kt)) | 3 |
| Brody | P(t) = VF(1 − b.e(−kt)) | 3 |
| Richards | P(t) = VF (1 − b.e(−kt))m | 4 |
| Dual-pool Logistic | P(t) = V1F/(1 + e(2−4.k1(t−λ))) + V2F/(1 + e(2−4.k2(t−λ))) | 5 |
| Model | Mean | SD | R2 | p-Value | CCC | RMSEP | Decomposition of MSEP (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| ME | SB | RE | |||||||
| Gompertz | 7.88 | 2.02 | 0.736 | 0.664 | 0.845 | 1.23 | 0.007 | 0.100 | 99.893 |
| Groot | 7.87 | 2.10 | 0.770 | 0.999 | 0.871 | 1.14 | 0.000 | 0.000 | 100.000 |
| Ørskov | 7.87 | 2.08 | 0.751 | 0.999 | 0.858 | 1.19 | 0.000 | 0.000 | 100.000 |
| Brody | 7.87 | 2.08 | 0.751 | 0.999 | 0.858 | 1.19 | 0.000 | 0.000 | 100.000 |
| Richards | 7.87 | 2.10 | 0.771 | 0.998 | 0.870 | 1.15 | 0.000 | 0.000 | 100.000 |
| Dual-pool Logistic | 7.88 | 2.05 | 0.700 | 0.569 | 0.865 | 1.16 | 0.009 | 0.140 | 99.85 |
| Observed data | 7.87 | 2.40 | - | - | - | - | - | - | - |
| Parameters | Treatments | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| Monensin | Monensin + B. toyonensis | Monensin + S. boulardii | B. toyonensis + S. boulardii | |||
| VF | 9.16 b | 11.11 ab | 9.22 b | 12.08 a | 0.0022 | 0.433 |
| k | 3.15 b | 5.24 a | 4.01 ab | 4.40 a | 0.0193 | 0.452 |
| λ | 1.07 a | 1.11 a | 0.66 b | 0.81 b | <0.001 | 0.038 |
| Parameters | Treatments | p-Value | SEM | |||
|---|---|---|---|---|---|---|
| Monensin | Monensin + B. toyonensis | Monensin + S. boulardii | B. toyonensis + S. boulardii | |||
| V1F | 3.966 b | 4.767 ab | 4.364 ab | 4.829 a | 0.024 | 0.114 |
| k1 | 0.513 | 0.506 | 0.535 | 0.482 | 0.921 | 0.026 |
| λ | 0.279 b | 0.440 ab | 0.345 ab | 0.569 a | 0.045 | 0.039 |
| V2F | 7.559 | 7.519 | 6.782 | 6.860 | 0.758 | 0.329 |
| k2 | 0.040 | 0.038 | 0.034 | 0.034 | 0.731 | 0.002 |
| VF | 11.525 | 12.286 | 11.146 | 11.689 | 0.762 | 0.378 |
| R2 | 0.958 | 0.980 | 0.972 | 0.981 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva Zornitta, C.; Ítavo, L.C.V.; Ítavo, C.C.B.F.; dos Santos, G.T.; Dias, A.M.; dos Santos Difante, G.; Gurgel, A.L.C. Kinetics of In Vitro Gas Production and Fitting Mathematical Models of Corn Silage. Fermentation 2021, 7, 298. https://doi.org/10.3390/fermentation7040298
da Silva Zornitta C, Ítavo LCV, Ítavo CCBF, dos Santos GT, Dias AM, dos Santos Difante G, Gurgel ALC. Kinetics of In Vitro Gas Production and Fitting Mathematical Models of Corn Silage. Fermentation. 2021; 7(4):298. https://doi.org/10.3390/fermentation7040298
Chicago/Turabian Styleda Silva Zornitta, Camila, Luis Carlos Vinhas Ítavo, Camila Celeste Brandão Ferreira Ítavo, Geraldo Tadeu dos Santos, Alexandre Menezes Dias, Gelson dos Santos Difante, and Antonio Leandro Chaves Gurgel. 2021. "Kinetics of In Vitro Gas Production and Fitting Mathematical Models of Corn Silage" Fermentation 7, no. 4: 298. https://doi.org/10.3390/fermentation7040298