Changes Occurring in Spontaneous Maize Fermentation: An Overview


 and
and 
Abstract
:1. Introduction
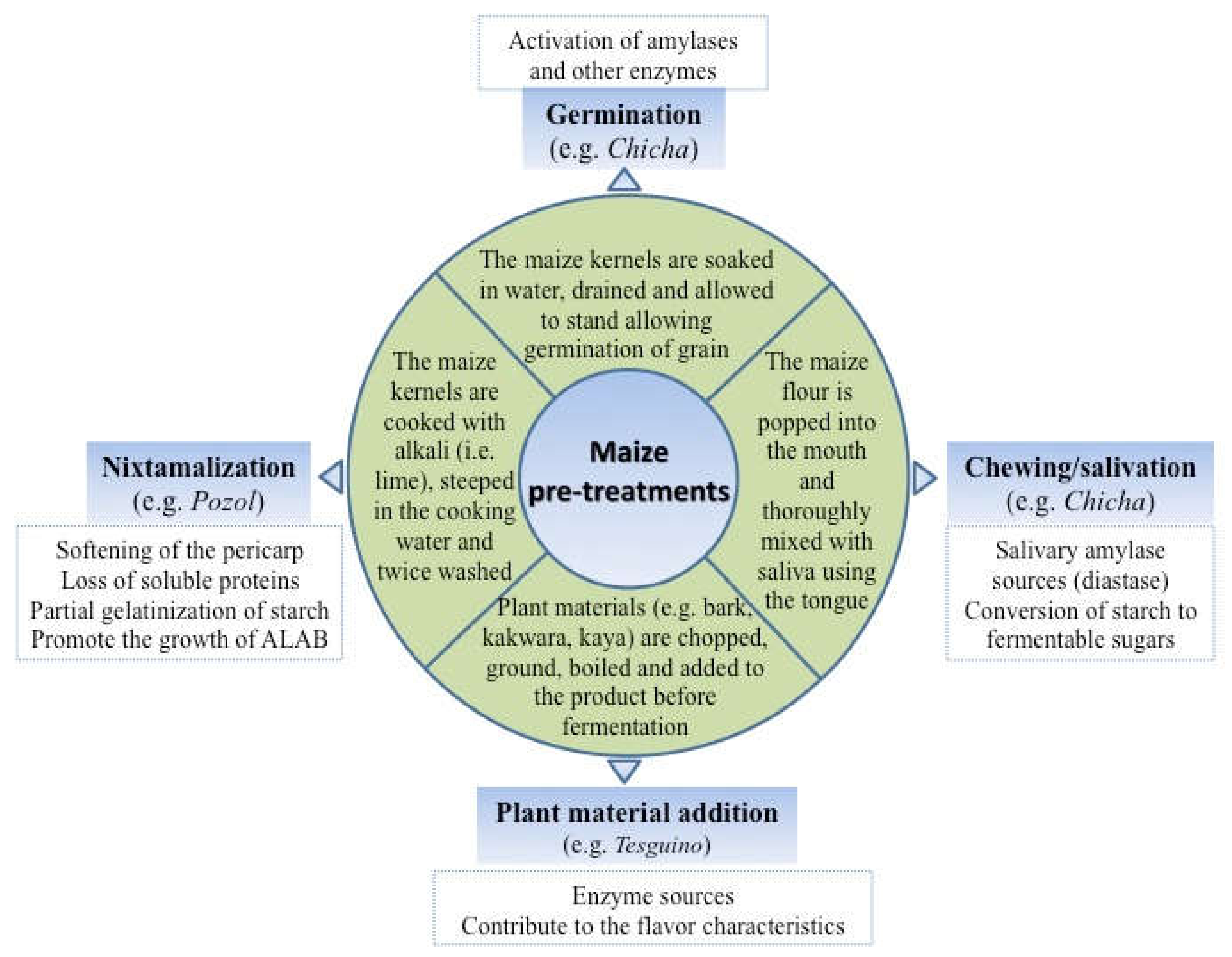
2. Maize Fermentation
3. Production of Amylolytic Enzymes
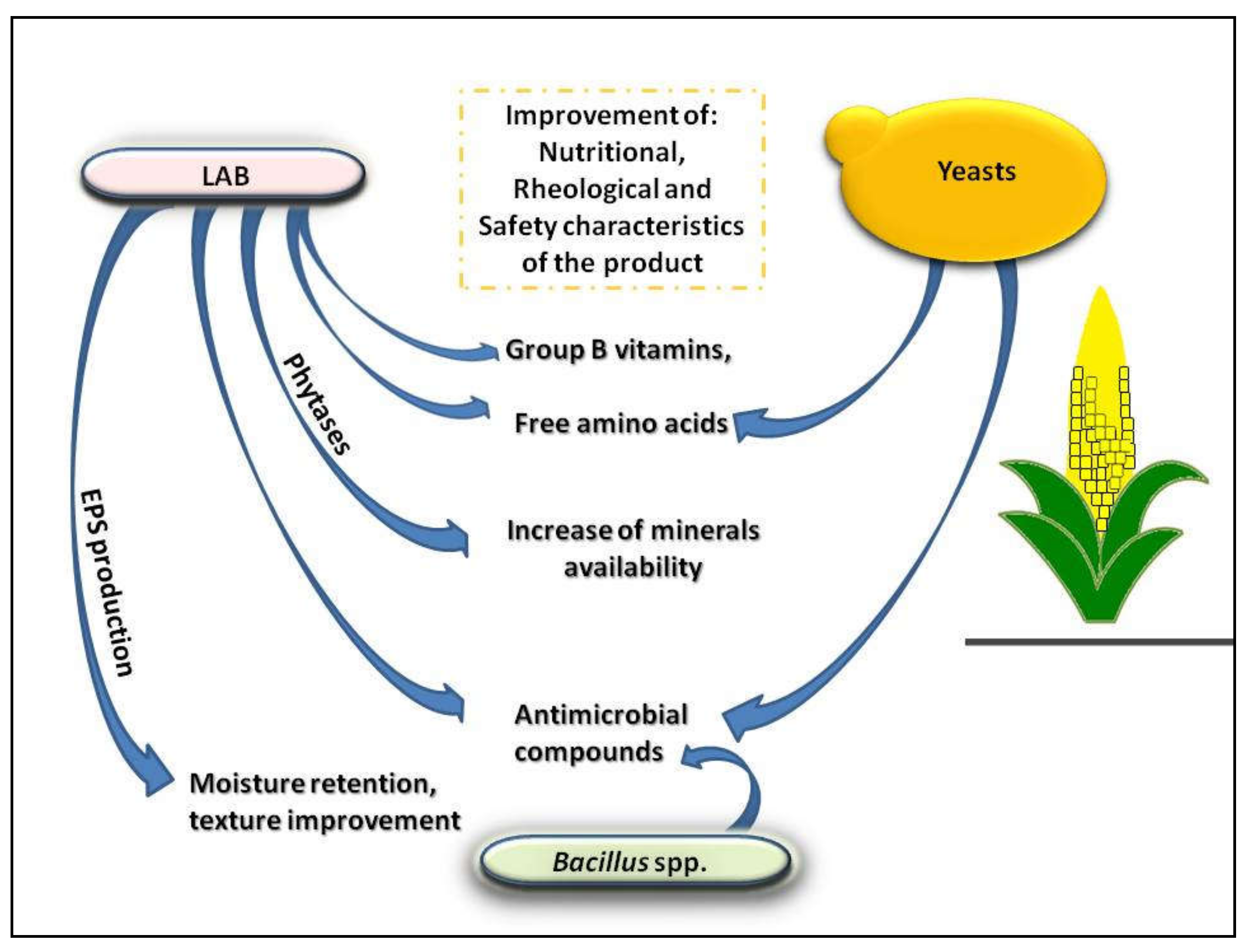
4. Production of Exopolysaccharides (EPS)
5. Vitamins and Amino Acids Increase
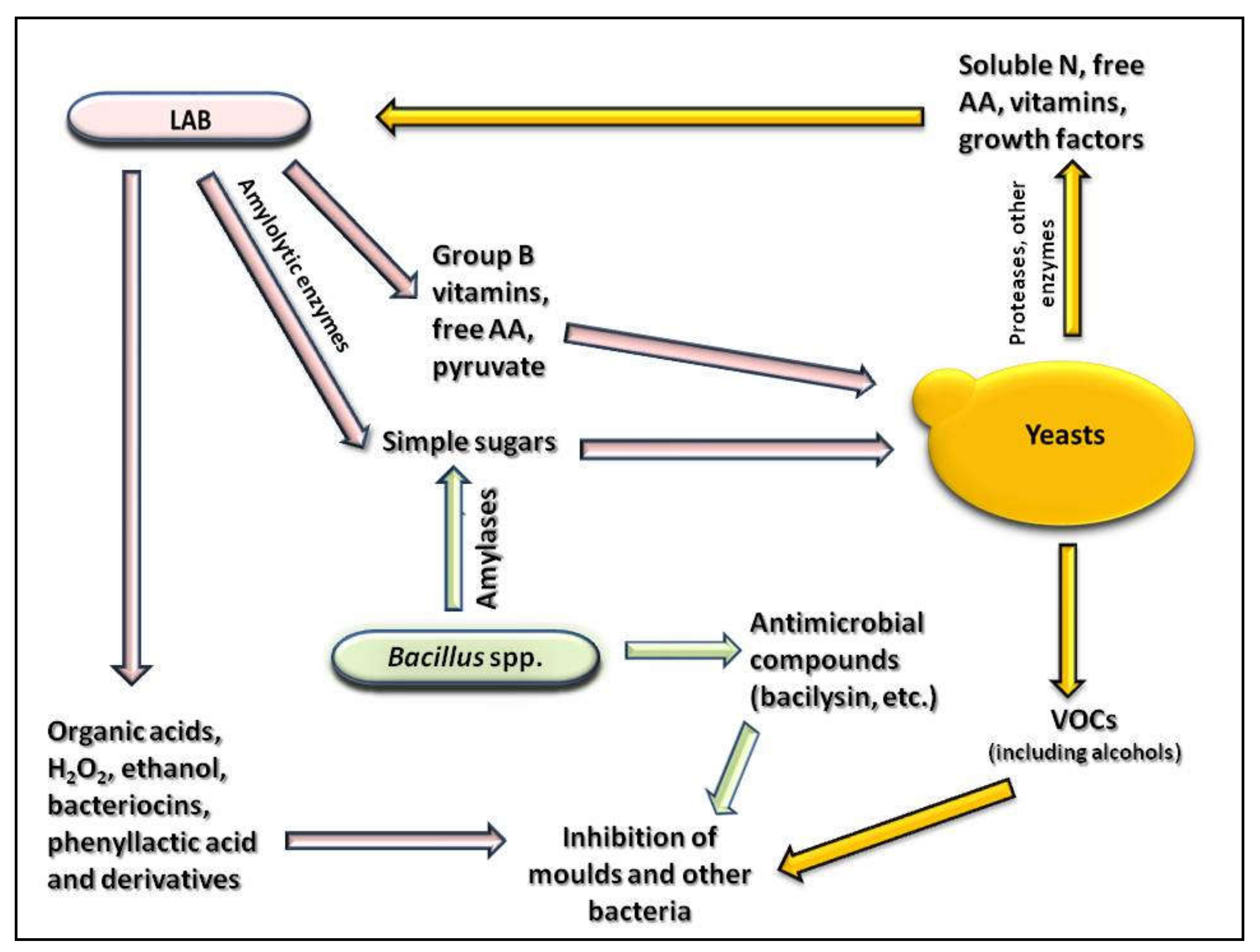
6. Production of Antimicrobial Compounds
7. Reduction of Phytates
8. Reduction of Mycotoxins
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nwokoro, O.; Chukwu, B.C. Studies on Akamu, a traditional fermented maize food. Rev. Chil. Nutr. 2012, 39, 180–184. [Google Scholar]
- Obinna-Echem, P.C.; Kuri, V.; Beal, J. Evaluation of the microbial community, acidity and proximate composition of akamu, a fermented maize food. J. Sci. Food Agric. 2014, 94, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Obinna-Echem, P.C.; Beal, J.; Kuri, V. Effect of processing method on the mineral content of Nigerian fermented maize infant complementary food – Akamu. LWT-Food Sci. Technol. 2015, 61, 145–151. [Google Scholar] [CrossRef]
- Pérez-Cataluña, A.; Elizaquível, P.; Carrasco, P.; Espinosa, J.; Reyes, D.; Wacher, C.; Aznar, R. Diversity and dynamics of lactic acid bacteria in Atole agrio, a traditional maize-based fermented beverage from South-Eastern Mexico, analysed by high throughput sequencing and culturing. Antonie van Leeuwenhoek 2018, 111, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Väkeväinen, K.; Valderrama, A.; Espinosa, J.; Centurión, D.; Rizo, J.; Reyes-Duarte, D.; Díaz-Ruiz, G.; von Wright, A.; Elizaquível, P.; Esquivel, K.; et al. Characterization of lactic acid bacteria recovered from atole agrio, a traditional Mexican fermented beverage. LWT-Food Sci. Technol. 2018, 88, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Steinkraus, K.H. Handbook of Indigenous Fermented Food, 2nd ed.; Marcel Dekker: New York, NY, USA, 1996. [Google Scholar]
- Von Mollendorff, J.W.; Vaz-Velho, M.; Todorov, S.D. Boza, a traditional cereal-based fermented beverage: A rich source of probiotics and bacteriocin-producing lactic acid bacteria. In Functional Properties of Traditional Foods; Kristbergsson, K., Ötles, S., Eds.; Springer: Boston, MA, USA, 2016; pp. 157–188. [Google Scholar]
- Okaru, A.O.; Abuga, K.O.; Kibwage, I.O.; Hausler, T.; Luy, B.; Kuballa, T.; Rhem, J.; Lachenmeier, D.W. Aflatoxin contamination in unrecorded beers from Kenya–A health risk beyond ethanol. Food Control 2017, 79, 344–348. [Google Scholar] [CrossRef]
- Osorio-Cadavid, E.; Chaves-López, C.; Tofalo, R.; Paparella, A.; Suzzi, G. Detection and identification of wild yeasts in Champús, a fermented Colombian maize beverage. Food Microbiol. 2008, 25, 771–777. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Grande-Tovar, C.D.; Cuervo-Mulet, R.; Delgado-Ospina, J.; Paparella, A. Traditional fermented foods and beverages from a microbiological and nutritional perspective: The Colombian heritage. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1031–1048. [Google Scholar] [CrossRef] [Green Version]
- Elizaquível, P.; Pérez-Cataluña, A.; Yépez, A.; Aristimuño, C.; Jiménez, E.; Cocconcelli, P.S.; Vignolo, G.; Aznar, R. Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chicha, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 2015, 198, 9–18. [Google Scholar] [CrossRef]
- Mendoza, L.M.; Neef, A.; Vignolo, G.; Belloch, C. Yeast diversity during the fermentation of Andean chicha: A comparison of high-throughput sequencing and culture-dependent approaches. Food Microbiol. 2017, 67, 1–10. [Google Scholar] [CrossRef]
- Assohoun, M.C.N.; Djéni, T.N.; N’Guessan, F.K.; Koussemon, M. Preliminary study on antimicrobial properties of Lactic acid bacteria involved in the fermentation of corn dough during doklu processing in Côte D’Ivoire. Food 2012, 6, 65–70. [Google Scholar]
- Assohoun, M.C.; Djeni, T.N.; Koussémon-Camara, M.; Brou, K. Effect of fermentation process on nutritional composition and aflatoxins concentration of doklu, a fermented maize based food. Food Nutr. Sci. 2013, 4, 1120. [Google Scholar]
- Assohoun-Djeni, N.M.C.; Djeni, N.T.; Messaoudi, S.; Lhomme, E.; Koussemon-Camara, M.; Ouassa, T.; Chobert, J.M.; Onno, B.; Dousset, X. Biodiversity, dynamics and antimicrobial activity of lactic acid bacteria involved in the fermentation of maize flour for doklu production in Côte d’Ivoire. Food Control 2016, 62, 397–404. [Google Scholar] [CrossRef]
- Greppi, A.; Rantsiou, K.; Padonou, W.; Hounhouigan, J.; Jespersen, L.; Jakobsen, M.; Cocolin, L. Determination of yeast diversity in ogi, mawè, gowé and tchoukoutou by using culture-dependent and-independent methods. Int. J. Food Microbiol. 2013, 165, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Adinsi, L.; Akissoé, N.H.; Dalodé-Vieira, G.; Anihouvi, V.B.; Fliedel, G.; Mestres, C.; Hounhouigan, J.D. Sensory evaluation and consumer acceptability of a beverage made from malted and fermented cereal: Case of gowe from Benin. Food Sci Nutr 2015, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Adinsi, L.; Mestres, C.; Akissoé, N.; Vieira-Dalodé, G.; Anihouvi, V.; Durand, N.; Hounhouigan, D.J. Comprehensive quality and potential hazards of gowe, a malted and fermented cereal beverage from West Africa. A diagnostic for a future re-engineering. Food Control. 2017, 82, 18–25. [Google Scholar] [CrossRef]
- Gadaga, T.H.; Mutukumira, A.N.; Narvhus, J.A.; Feresu, S.B. A review of traditional fermented foods and beverages of Zimbabwe. Int. J. Food Microbiol. 1999, 53, 1–11. [Google Scholar] [CrossRef]
- Kalui, C.M.; Mathara, J.M.; Kutima, P.M.; Kiiyukia, C.; Wongo, L.E. Partial characterisation and identification of lactic acid bacteria involved in the production of ikii: A traditional fermented maize porridge by the Kamba of Kenya. J. Trop Microbiol. Biotechnol. 2008, 4, 3–15. [Google Scholar] [CrossRef]
- Kalui, C.M.; Mathara, J.M.; Kutima, P.M.; Kiiyukia, C.; Wongo, L.E. Functional characteristics of Lactobacillus plantarum and Lactobacillus rhamnosus from ikii, a Kenyan traditional fermented maize porridge. Afr. J. Biotechnol. 2009, 8, 4363–4373. [Google Scholar]
- Mkandawire, P.; Luginaah, I.; Tobias, J. Landscapes of economic deprivation and locally distilled liquor (kachasu): An emerging milieu of HIV/AIDS risk in urban Northern Malawi. Environ. Plan A. 2011, 43, 2384–2398. [Google Scholar] [CrossRef]
- Jespersen, L.; Halm, M.; Kpodo, K.; Jakobsen, M. Significance of yeasts and moulds occurring in maize dough fermentation for ‘kenkey’production. Int. J. Food Microbiol. 1994, 24, 239–248. [Google Scholar] [CrossRef]
- Nout, M.J.R.; Kok, B.; Vela, E.; Nche, P.F.; Rombouts, F.M. Acceleration of the fermentation of kenkey, an indigenous fermented maize food of Ghana. Food Res. Int. 1996, 28, 599–604. [Google Scholar] [CrossRef]
- Atter, A.; Ofori, H.; Anyebuno, G.A.; Amoo-Gyasi, M.; Amoa-Awua, W.K. Safety of a street vended traditional maize beverage, ice-kenkey, in Ghana. Food Control. 2015, 55, 200–205. [Google Scholar] [CrossRef]
- Oduro-Yeboah, C.; Mestres, C.; Amoa-Awua, W.; Fliedel, G.; Durand, N.; Matignon, B.; Michodjehoun, V.L.; Saalia, F.K.; Sakyi-Dawson, E.; Abbey, L. Steeping time and dough fermentation affect the milling behaviour and quality of white kenkey(nsiho), a sour stiff dumpling prepared from dehulled maize grains. J. Cereal. Sci. 2016, 69, 377–382. [Google Scholar] [CrossRef]
- Andah, A.; Muller, H.G. Studies on koko, a Ghanaian fermented maize porridge. Ghana J Agric Sci 1973, 6, 103–108. [Google Scholar]
- Oranusi, S.; Dahunsi, S.O. Preliminary study on hazards and critical control points of kokoro, a Nigerian indigenous fermented maize snack. SpringerPlus. 2015, 4, 253. [Google Scholar] [CrossRef] [Green Version]
- Bolade, M.K. Physical and organoleptic characteristics of non-sour ‘kokoro’(a Nigerian maize-based snack) as influenced by flour particle size differential. LWT-Food Sci. Technol. 2018, 87, 287–292. [Google Scholar] [CrossRef]
- Tchikoua, R.; Tatsadjieu, N.L.; Mbofung, C.M.F. Effect of selected lactic acid bacteria on growth of Aspergillus flavus and aflatoxin B 1 production in kutukutu. J Microbiol Res 2015, 5, 84–94. [Google Scholar]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Prado, F.C.; Parada, J.L.; Pandey, A.; Soccol, C.R. Trends in non-dairy probiotic beverages. Food Res. Int. 2008, 41, 111–123. [Google Scholar] [CrossRef]
- Chelule, P.K.; Mbongwa, H.P.; Carries, S.; Gqaleni, N. Lactic acid fermentation improves the quality of amahewu, a traditional South African maize-based porridge. Food Chem. 2010, 122, 656–661. [Google Scholar] [CrossRef]
- Franz, C.M.; Huch, M.; Mathara, J.M.; Abriouel, H.; Benomar, N.; Reid, G.; Galvez, A.; Holzapfel, W.H. African fermented foods and probiotics. Int. J. Food Microbiol. 2014, 190, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Awobusuyi, T.D.; Siwela, M.; Kolanisi, U.; Amonsou, E.O. Provitamin A retention and sensory acceptability of amahewu, a non-alcoholic cereal-based beverage made with provitamin A-biofortified maize. J. Sci. Food Agric. 2016, 96, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Chaves-López, C.; Serio, A.; Delgado-Ospina, J.; Rossi, C.; Grande-Tovar, C.D.; Paparella, A. Exploring the bacterial microbiota of Colombian fermented maize dough “Masa Agria” (Maiz Añejo). Front Microbiol. 2016, 7, 1168. [Google Scholar] [CrossRef]
- Ramos, C.L.; Schwan, R.F. Technological and nutritional aspects of indigenous Latin America fermented foods. Curr. Opin. Food Sci. 2017, 13, 97–102. [Google Scholar] [CrossRef]
- Fowoyo, P.T.; Ogunbanwo, S.T. Phenotypic diversity of lactic acid bacteria isolated from Massa, a fermented maize dough. Afr. J. Microbiol. Res. 2010, 4, 2682–2691. [Google Scholar]
- Hounhouigan, D.J.; Nout, M.J.; Nago, C.M.; Houben, J.H.; Rombouts, F.M. Characterization and frequency distribution of species of lactic acid bacteria involved in the processing of mawé, a fermented maize dough from Benin. Int. J. Food Microbiol. 1993, 18, 279–287. [Google Scholar] [CrossRef]
- Hounhouigan, D.J.; Nout, M.J.; Nago, C.M.; Houben, J.H.; Rombouts, F.M. Composition and microbiological and physical attributes of mawè, a fermented maize dough from Benin. Int. J. Food Sci. Technol. 1993, 28, 513–517. [Google Scholar] [CrossRef]
- Agati, V.; Guyot, J.P.; Morlon-Guyot, J.; Talamond, P.; Hounhouigan, D.J. Isolation and characterization of new amylolytic strains of Lactobacillus fermentum from fermented maize doughs (mawè and ogi) from Benin. J. Appl. Microbiol. 1998, 85, 512–520. [Google Scholar] [CrossRef]
- Zulu, R.M.; Dillon, V.M.; Owens, J.D. Munkoyo beverage, a traditional Zambian fermented maize gruel using Rhynchosia root as amylase source. Int. J. Food Microbiol. 1997, 34, 249–258. [Google Scholar] [CrossRef]
- Foma, R.K.; Destain, J.; Mobinzo, P.K.; Kayisu, K.; Thonart, P. Study of physicochemical parameters and spontaneous fermentation during traditional production of munkoyo, an indigenous beverage produced in Democratic Republic of Congo. Food Control. 2012, 25, 334–341. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Kasase, C.; Toarta, C.; Kassen, R.; Poulain, A.J. Microbial community structure of three traditional Zambian fermented products: Mabisi, chibwantu and munkoyo. PLoS ONE 2013, 8, e63948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omemu, A.M.; Oyewole, O.B.; Bankole, M.O. Significance of yeasts in the fermentation of maize for ogi production. Food Microbiol. 2007, 24, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Omemu, A.M. Fermentation dynamics during production of ogi, a Nigerian fermented cereal porridge. Rep. Opin. 2011, 3, 8–17. [Google Scholar]
- Bolaji, O.T.; Adepoju, P.A.; Olalusi, A.P. Economic implication of industrialization of a popular weaning food ogi production in Nigeria: A review. Afr. J. Food Sci. 2015, 9, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Izah, S.C.; Kigigha, L.T.; Okowa, I.P. Microbial quality assessment of fermented maize Ogi (a cereal product) and options for overcoming constraints in production. Biotechnol Res. 2016, 2, 81–93. [Google Scholar]
- Henshaw, E.; Akpan, J.; Lennox, J.; Frank, E. Bacterial community dynamics during the production of ogi from millet ‘a Nigerian fermented food’ using culture-dependent approach. Imp. J. Interdiscip. Res. 2016, 2, 621–623. [Google Scholar]
- Ekundayo, J.A. The production of pito, a Nigerian fermented beverage. J. Food Technol. 1969, 4, 217–225. [Google Scholar] [CrossRef]
- Onaghise, E.O.; Izuagbe, Y.S. Improved brewing and preservation of pito, a nigerian alcoholic beverage from maize. Eng. Life Sci. 1989, 9, 137–142. [Google Scholar] [CrossRef]
- Louembé, D.; Kéléké, S.; Kobawila, S.C.; Nzoussi, J.P. Variabilité et amélioration de la technologie traditionnelle de production de la pâte fermentée de maïs au Congo. Tropicultura 2004, 22, 211–218. [Google Scholar]
- Abriouel, H.; Omar, N.B.; López, R.L.; Martínez-Cañamero, M.; Keleke, S.; Gálvez, A. Culture-independent analysis of the microbial composition of the African traditional fermented foods poto poto and dégué by using three different DNA extraction methods. Int. J. Food Microbiol. 2006, 111, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Ben-Omar, N.; Abriouel, H.; Keleke, S.; Valenzuela, A.S.; Martínez-Cañamero, M.; López, R.L.; Ortega, E.; Gálvez, A. Bacteriocin-producing Lactobacillus strains isolated from poto poto, a Congolese fermented maize product, and genetic fingerprinting of their plantaricin operons. Int. J. Food Microbiol. 2008, 127, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wacher, C.; Canas, A.; Cook, P.E.; Barzana, E.; Owens, J.D. Sources of microorganisms in pozol, a traditional Mexican fermented maize dough. World J. Microbiol. Biotechnol. 1993, 9, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Ben-Omar, N.; Ampe, F. Microbial community dynamics during production of the Mexican fermented maize dough pozol. Appl. Environ. Microbiol. 2000, 66, 3664–3673. [Google Scholar] [CrossRef] [Green Version]
- Escalante, A.; Wacher, C.; Farres, A. Lactic acid bacterial diversity in the traditional Mexican fermented dough pozol as determined by 16S rDNA sequence analysis. Int. J. Food Microbiol. 2001, 64, 21–31. [Google Scholar] [CrossRef]
- Diaz-Ruiz, G.; Guyot, J.P.; Ruiz-Teran, F.; Morlon-Guyot, J.; Wacher, C. Microbial and physiological characterization of weakly amylolytic but fast-growing lactic acid bacteria: A functional role in supporting microbial diversity in pozol, a Mexican fermented maize beverage. Appl. Environ. Microbiol. 2003, 69, 4367–4374. [Google Scholar] [CrossRef] [Green Version]
- Phister, T.G.; O’Sullivan, D.J.; McKay, L.L. Identification of bacilysin, chlorotetaine, and iturin A produced by Bacillus sp. strain CS93 isolated from pozol, a Mexican fermented maize dough. Appl. Environ. Microbiol. 2004, 70, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Wacher-Rodarte, M.D.C.; Trejo-Muñúzuri, T.P.; Montiel-Aguirre, J.F.; Drago-Serrano, M.E.; Gutiérrez-Lucas, R.L.; Castañeda-Sánchez, J.I.; Sainz-Espuñes, T. Antibiotic resistance and multidrug-resistant efflux pumps expression in lactic acid bacteria isolated from pozol, a nonalcoholic Mayan maize fermented beverage. Food Sci. Nutr. 2015, 4, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Sanni, A.I. Chemical studies on sekete beer. Food Chem. 1989, 33, 187–191. [Google Scholar] [CrossRef]
- Adegoke, G.O.; Nwaigwe, R.N.; Oguntimein, G.B. Microbiological and biochemical changes during the production of sekete- A fermented beverage made from maize. J. Food Sci. Technol. 1995, 32, 516–518. [Google Scholar]
- Sangwan, S.; Kumar, S.; Goyal, S. Maize utilisation in food bioprocessing: An overview. In Maize: Nutrition Dynamics and Novel Uses; Chaudhary, D.P., Kumar, S., Langyan, S., Eds.; Springer: New Delhi, India, 2014; pp. 119–134. [Google Scholar]
- Mugula, J.K.; Nnko, S.A.M.; Narvhus, J.A.; Sørhaug, T. Microbiological and fermentation characteristics of togwa, a Tanzanian fermented food. Int. J. Food Microbiol. 2003, 80, 187–199. [Google Scholar] [CrossRef]
- Hjortmo, S.B.; Hellström, A.M.; Andlid, T.A. Production of folates by yeasts in Tanzanian fermented togwa. FEMS Yeast Res. 2008, 8, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.; Mejia, D. Traditional Fermented Food and Beverages for Improved Livelihoods; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 1–79. [Google Scholar]
- Capozzi, V.; Fragrasso, M.; Russo, P. Microbiological safety and the management of microbial resources in artisanal foods and beverages: The need for a transdisciplinary assessment to conciliate actual trends and risks avoidance. Microorganisms 2020, 8, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves-López, C.; Serio, A.; Rossi, C.; Pepe, A.; Compagnone, E.; Paparella, A. Interaction between Galactomyces geotrichum KL20B, Lactobacillus plantarum LAT3 and Enterococcus faecalis KE06 during milk fermentation. Fermentation 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Ramírez, L.L.; Díaz-Ruiz, G.; Wacher, C. Maize (Zea mays L. subsp. mays) Fermentation. In Fermented Foods, Part II: Technological Interventions; Ray, R.C., Montet, D., Eds.; Taylor & Francis: Abingdon-on-Thames, UK, 2017. [Google Scholar]
- Soro-Yao, A.A.; Brou, K.; Amani, G.; Thonart, P.; Djè, K.M. The use of lactic acid bacteria starter cultures during the processing of fermented cereal-based foods in West Africa: A review. Trop Life Sci. Res. 2014, 25, 81–100. [Google Scholar] [PubMed]
- Achi, O.K.; Ukwuru, M. Cereal-based fermented foods of Africa as functional foods. Int. J. Microbiol. Appl. 2015, 2, 71–83. [Google Scholar]
- Kunene, N.F.; Geornaras, I.; von Holy, A.; Hastings, J.W. Characterization and determination of origin of lactic acid bacteria from a sorghum-based fermented weaning food by analysis of soluble proteins and amplified fragment lenght polymorphism fingerprinting. Appl. Environl. Microbiol. 2000, 66, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Ampe, F.; Miambi, E. Cluster analysis, richness and biodiversity indexes derived from denaturing gradient gel electrophoresis fingerprints of bacterial communities demonstrate that traditional maize fermentations are driven by the transformation process. Int. J. Food Microbiol. 2000, 60, 91–97. [Google Scholar] [CrossRef]
- Bassi, D.; Orrù, L.; Vasquez, J.C.; Cocconcelli, P.S.; Fontana, C. Peruvian chica: A focus on the microbial populations of this ancient maize-based fermented beverage. Microorganisms 2020, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Okeke, C.A.; Ezekiel, C.N.; Nwangburuka, C.C.; Sulyok, M.; Ezeamagu, C.O.; Adeleke, R.A.; Dike, S.K.; Krska, R. Bacterial diversity and mycotoxin reduction during maize fermentation (steeping) for ogi production. Front Microbiol. 2015, 6, 1402. [Google Scholar] [CrossRef] [Green Version]
- Obatolu, V.A.; Adeniyi, P.O.; Ashaye, O.A. Nutritional, sensory and storage quality of Sekete from Zea mays. Int J Food Sci Nutr Eng 2016, 6, 73–80. [Google Scholar]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredlund, E.; Druvefors, U.Ä.; Olstorpe, M.N.; Passoth, V.; Schnürer, J. Influence of ethyl acetate production and ploidy on the anti-mould activity of Pichia anomala. FEMS Microbiol. Lett. 2004, 238, 133–137. [Google Scholar] [PubMed]
- Ponomarova, O.; Gabrielli, N.; Sévin, D.C.; Mülleder, M.; Zirngibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast creates a niche for symbiotic lactic acid bacteria through nitrogen overflow. Cell Syst. 2017, 5, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, E.G.; Rachid, C.C.; Schwan, R.F. Microbial population present in fermented beverage ‘cauim’produced by Brazilian Amerindians. Int. J. Food Microbiol. 2007, 120, 146–151. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Tech. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Teniola, O.D.; Odunfa, S.A.; Holzapfel, W.H. Selection, use and the influence of starter cultures in the nutrition and processing improvement of Ogi. In Proceedings of the 2nd international workshop: Food-based approaches for a healthy nutrition; Brouwer, I.D., Traore, A.S., Treche, S., Eds.; Universitaires de Ouagadougou: Ouagadougou, Burkina Faso, 2005; pp. 697–708. [Google Scholar]
- Reddy, G.; Altaf, M.D.; Naveena, B.J.; Venkateshwar, M.; Kumar, E.V. Amylolytic bacterial lactic acid fermentation-a review. Biotechnol. Adv. 2008, 26, 22–34. [Google Scholar] [CrossRef]
- Humblot, C.; Turpin, W.; Chevalier, F.; Picq, C.; Rochette, I.; Guyot, J.P. Determination of expression and activity of genes involved in starch metabolism in Lactobacillus plantarum A6 during fermentation of a cereal-based gruel. Int. J. Food Microbiol. 2014, 185, 103–111. [Google Scholar] [CrossRef]
- Mestres, C.; Nguyen, T.C.; Adinsi, L.; Hounhouigan, J.D.; Fliedel, G.; Loiseau, G. The interaction between starch hydrolysis and acidification kinetic determines the quality of a malted and fermented sorghum beverage. J. Cereal Sci. 2015, 63, 8–13. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K.; Stoyancheva, G. Starch-modifying enzymes of lactic acid bacteria – structures, properties and applications. Strach Stärke 2013, 65, 34–47. [Google Scholar] [CrossRef]
- Betancourt Botero, S.P.; Bolívar Escobar, G.A.; Ramírez Toro, C. Fermentación de maíz de alta calidad proteica con Lactobacillus plantarum (CPQBA 087-11 DRM) aislado en Colombia de masas tradicionales fermentadas. Rev Argent Microbiol 2013, 45, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Grijalva-Vallejos, N.; Aranda, A.; Matallana, E. Evaluation of yeasts from Ecuadorian chicha by their performance as starters for alcoholic fermentations in the food industry. Int. J. Food Microbiol. 2020, 317, 108462. [Google Scholar] [CrossRef] [PubMed]
- Rahmawati, J.; Dewanti-Hariyadi, R.; Hariyadi, P.; Fardiaz, D.; Richana, N. Isolation and identification of microorganisms during spontaneous fermentation of maize. J. Teknol. Dan. Industri. Pangan. 2013, 24, 33–39. [Google Scholar]
- Resende, L.V.; Pinheiro, L.K.; Miguel, M.G.D.C.P.; Ramos, C.L.; Vilela, D.M.; Schwan, R.F. Microbial community and physicochemical dynamics during the production of ‘Chicha’, a traditional beverage of Indigenous people of Brazil. World J. Microbiol. Biotechnol. 2018, 34, 46. [Google Scholar] [CrossRef]
- Sachdev, S.; Ojha, S.K.; Mishra, S. Bacillus spp. amylase: Production, isolation, characterisation and its application. Int. J. Appl. Sci. Biotechnol. 2016, 4, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Putri, W.D.R.; Marseno, D.W.; Cahyanto, M.N. Role of lactic acid bacteria on structural and physicochemical properties of sour cassava starch. APCBEE Procedia. 2012, 2, 104–109. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Fermentation: Assessment and research. Report of a Joint FAO/WHO Workshop on fermentation as household technology to improve food safety; WHO Document WHO/FNU/FOS/96.1; World Health Organization: Geneva, Switzerland, 1996; pp. 1–79. [Google Scholar]
- Donot, F.; Fontana, A.; Baccou, J.C.; Schorr-Galindo, S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Serrazanetti, D.I.; Guerzoni, M.E.; Corsetti, A.; Vogel, R. Metabolic impact and potential exploitation of the stress reactions in lactobacilli. Food Microbiol. 2009, 26, 700–711. [Google Scholar] [CrossRef]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef]
- Falade, A.T.; Emmambux, M.N.; Buys, E.M.; Taylor, J.R. Improvement of maize bread quality through modification of dough rheological properties by lactic acid bacteria fermentation. J. Cereal Sci. 2014, 60, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Tamani, R.J.; Goh, K.K.T.; Brennan, C.S. Physico-chemical properties of sourdough bread production using selected lactobacilli starter cultures. J. Food Qual. 2013, 36, 245–252. [Google Scholar] [CrossRef]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of exopolysaccharides from lactic acid bacteria. Indian J. Microbiol. 2012, 52, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebayo-tayo, B.C.; Onilude, A.A. Screening of lactic acid bacteria strains isolated from some Nigerian fermented foods for EPS production. World Appl. Sci. J. 2008, 4, 741–747. [Google Scholar]
- Gänzle, M.; Schwab, C. Ecology of exopolysaccharide formation by lactic acid bacteria: Sucrose utilization, stress tolerance, and biofilm formation. In Bacterial Polysaccharides, Current Innovations and Future Trends; Ullrich, M., Ed.; Caister Academic Press: Norfolk, UK, 2009; pp. 263–278. [Google Scholar]
- Galle, S.; Schwab, C.; Arendt, E.; Gänzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Characterization of growth and exopolysaccharide production of selected acetic acid bacteria in buckwheat sourdoughs. Int. J. Food Microbiol. 2016, 239, 103–112. [Google Scholar] [CrossRef]
- Hiran, P.; Kerdchoechuen, O.; Laohakunjit, N. Combined effects of fermentation and germination on nutritional compositions, functional properties and volatiles of maize seeds. J. Cereal Sci. 2016, 71, 207–216. [Google Scholar] [CrossRef]
- Madhu, A.N.; Giribhattanavar, P.; Narayan, M.S.; Prapulla, S.G. Probiotic lactic acid bacterium from kanjika as a potential source of vitamin B12: Evidence from LC-MS, immunological and microbiological techniques. Biotechnol. Lett. 2010, 32, 503–506. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production potency of folate, vitamin B12, and thiamine by lactic acid bacteria isolated from japanese pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; Bottacini, F.; Fosso, B.; Kelleher, P.; Calasso, M.; Di Cagno, R.; Ventura, M.; Picardi, E.; van Sinderen, D.; Gobbetti, M. Lactobacillus rossiae, a vitamin B12 producer, represents a metabolically versatile species within the genus Lactobacillus. PLoS ONE 2014, 9, e107232. [Google Scholar] [CrossRef] [Green Version]
- Murdock, F.A.; Fields, M.L. B-vitamin content of natural lactic acid fermented cornmeal. J. Food Sci. 1984, 49, 373–375. [Google Scholar] [CrossRef]
- Oladeji, B.S.; Akanbi, C.T.; Gbadamosi, S.O. Effects of fermentation on antioxidant properties of flours of a normal endosperm and quality protein maize varrieties. J. Food Meas. Caract. 2017, 11, 1148–1158. [Google Scholar] [CrossRef]
- Yépez, A.; Russo, P.; Spano, G.; Khomenko, I.; Biasoli, F.; Capozzi, V.; Aznar, R. In situ riboflavin fortification of different kefir-like cereal-based beverages using selected Andean LAB. Food Microbiol. 2019, 77, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ejigui, J.; Savoie, L.; Marin, J.; Desrosiers, T. Beneficial changes and drawbacks of a traditional fermentation process on chemical composition and antinutritional factors of yellow maize (Zea mays). J. Biol. Sci. 2005, 5, 590–596. [Google Scholar]
- Saubade, F.; Hemery, Y.M.; Guyot, J.P.; Humblot, C. Lactic acid fermentation as a tool for increasing the folate content of foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3894–3910. [Google Scholar] [CrossRef] [PubMed]
- Kariluoto, S.; Aittamaa, M.; Korhola, M.; Salovaara, H.; Vahteristo, L.; Piironen, V. Effects of yeasts and bacteria on the levels of folates in rye sourdoughs. Int. J. Food Microbiol. 2006, 106, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Fields, M.L.; Yoa, F.G. Nutritional improvement of corn meal by fermentation by Bacillus licheniformis and Enterobacter cloacae. J. Food Prot. 1990, 53, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Maemura, K.; Oki, K.; Shiraishi, N.; Shibayama, Y.; Katsu, K. Gamma-aminobutyric acid GABA and cell proliferation, focus on cancer cells. Histol. Histopathol. 2006, 21, 1135–1141. [Google Scholar]
- Olsen, A.; Halm, M.; Jakobsen, M. The antimicrobial activity of lactic acid bacteria from fermented maize (kenkey) and their interactions during fermentation. J. Appl. Microbiol. 1995, 79, 506–512. [Google Scholar] [CrossRef]
- Olasupo, N.A.; Olukoya, D.K.; Odunfa, S.A. Assessment of a bacteriocin-producing Lactobacillus strain in the control of spoilage of a cereal-based African fermented food. Folia. Microbiol. 1997, 42, 31–34. [Google Scholar] [CrossRef]
- Quillama, E. Producción de bacteriocinas por cepas de Lactobacillus aisladas de chicha de jora. Master’s Thesis, Universidad Nacional Mayor de San Marcos, Lima, Peru, 1998. [Google Scholar]
- Kaktcham, P.M.; Zambou, N.F.; Tchouanguep, F.M.; El-Soda, M.; Choudhary, M.I. Antimicrobial and safety properties of Lactobacilli isolated from two Cameroonian traditional fermented foods. Sci. Pharm. 2012, 80, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Onwuakor, C.E.; Nwaugo, V.O.; Nnadi, C.J.; Emetole, J.M. Effect of varied culture conditions on crude supernatant (bacteriocin) production from four Lactobacillus species isolated from locally fermented maize (Ogi). Am. J. Microbiol. Res. 2014, 2, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Oguntoyinbo, F.A.; Narbad, A. Multifunctional properties of Lactobacillus plantarum strains isolated from fermented cereal foods. J. Funct. Foods 2015, 17, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Ekwem, O.H. Isolation of antimicrobial producing lactobacilli from akamu (a Nigerian fermented cereal gruel). Afr. J. Microbiol. Res. 2014, 8, 718–720. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Moran, S.; Robertson, K.; Paradisi, F.; Rai, D.K.; Murphy, C.D. Production of lipopeptides in Bacillus sp. CS93 isolated from Pozol. FEMS Microbiol. Lett. 2010, 304, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Vats, P.; Banerjee, U.C. Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): An overview. Enzyme Microb. Technol. 2004, 35, 3–14. [Google Scholar] [CrossRef]
- Ragon, M.; Aumelas, A.; Chemardin, P.; Santiago, S.; Moulin, G.; Boze, H. Complete hydrolysis of myo-inositol hexakisphosphate by a novel phytase from Debaryomyces castellii CBS 2923. Appl. Microbiol. Biotechnol. 2008, 78, 47–53. [Google Scholar] [CrossRef]
- Agte, V.V.; Gokhale, M.K.; Chiplonkar, S.A. Effect of natural fermentation on in vitro zinc bioavailability in cereal-legume mixture. Int. J. Food Sci. Tech. 1997, 31, 29–32. [Google Scholar] [CrossRef]
- Coulibaly, A.; Kouakou, B.; Chen, J. Phytic acid in cereal grains: Structure, healthy or harmful ways to reduce phytic acid in cereal grains and their effects on nutritional quality. Amer J. Plant Nutr. Fertil. Tech. 2011, 1, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Afify, A.E.M.M.R.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Bioavailability of iron, zinc, phytate and phytase activity during soaking and germination of white sorghum varieties. PLoS ONE 2011, 6, e25512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markiewicz, L.H.; Honke, J.; Haros, M.; Świątecka, D.; Wróblewska, B. Diet shapes the ability of human intestinal microbiota to degrade phytate–in vitro studies. J. Appl. Microbiol. 2013, 115, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabaza, M.; Shumoy, H.; Muchuweti, M.; Vandamme, P.; Raes, K. Iron and zinc bioaccessibility of fermented maize, sorghum and millets from five locations in Zimbabwe. Food Res. Int. 2018, 103, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Lopez, Y.; Gordon, D.T.; Fields, M.L. Release of phosphorus from phytate by natural lactic acid fermentation. J. Food Sci. 1983, 48, 953–954. [Google Scholar] [CrossRef]
- Sandberg, A.S.; Andlid, T. Phytogenic and microbial phytases in human nutrition. Int. J. Food Sci. Technol. 2002, 37, 823–833. [Google Scholar] [CrossRef]
- Gernah, D.I.; Ariahu, C.C.; Ingbian, E.K. Effects of malting and lactic fermentation on some chemical and functional properties of maize (Zea mays). Am. J. Food Technol. 2011, 6, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Decimo, M.; Quattrini, M.; Ricci, G.; Fortina, M.G.; Brasca, M.; Silvetti, T.; Manini, F.; Erba, D.; Criscuoli, F.; Casiraghi, M.C. Evaluation of microbial consortia and chemical changes in spontaneous maize bran fermentation. AMB Express. 2017, 7, 205. [Google Scholar] [CrossRef]
- Murali, A.; Kapoor, R. Effect of natural and pure culture fermentation of finger millet on zinc availability as predicted from HCl extractability and molar ratios. J. Food Sci. Technol. 2003, 40, 112–114. [Google Scholar]
- Hotz, C.; Gibson, R.S. Assessment of home-based processing methods to reduce the phytate content and phytate/zinc molar ratio of white maize (Zea mays). J. Agric. Food Chem. 2001, 49, 692–698. [Google Scholar] [CrossRef]
- Sreeramulu, G.; Srinivasa, D.S.; Nand, K.; Joseph, R. Lactobacillus amylovorus as a phytase producer in submerged culture. Lett. Appl. Microbiol. 1996, 23, 385–388. [Google Scholar] [CrossRef]
- Nuobariene, L.; Hansen, A.S.; Jespersen, L.; Arneborg, N. Phytase-active yeasts from grain-based food and beer. J. Appl. Microbiol. 2011, 110, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.R.; Sanni, A.I.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Hellström, A.M.; Almgren, A.; Carlsson, N.G.; Svanberg, U.; Andlid, T.A. Degradation of phytate by Pichia kudriavzevii TY13 and Hanseniaspora guilliermondii TY14 in Tanzanian togwa. Int. J. Food. Microbiol. 2012, 153, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Gabiela-Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Abia, W.A.; Warth, B.; Ezekiel, C.N.; Sarkanj, B.; Turner, P.C.; Marko, D.; Krska, R.; Sulyok, M. Uncommon toxic microbial metabolite patterns in traditionally home-processed maize dish (fufu) consumed in rural Cameroon. Food Chem. Toxicol. 2017, 107, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef]
- Styriak, I.; Conková, E. Microbial binding and biodegradation of mycotoxins. Vet. Hum. Toxicol. 2002, 44, 358–361. [Google Scholar] [PubMed]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Dawlal, P.; Brabet, C.; Thantsha, M.S.; Buys, E.M. Potential of lactic acid bacteria for the reduction of fumonisin exposure in African fermented maize based foods. World Mycotoxin. J. 2017, 10, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Nout, M.J.R. Fermented foods and food safety. Food Res. Int. 1994, 27, 291–298. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. The toxicity and decreased concentration of aflatoxin B1 in natural lactic acid fermented maize meal. J. Appl. Microbiol. 2006, 100, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Fandohan, P.; Zoumenou, D.; Hounhouigan, D.J.; Marasas, W.F.O.; Wingfield, M.J.; Hell, K. Fate of aflatoxins and fumonisins during the processing of maize into food products in Benin. Int. J. Food Microbiol. 2005, 98, 249–259. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Mukisa, I.M.; Meeme, R.; Byakika, S.; Wendiro, D.; Sybesma, W.; Kort, R. Probiotic enrichment and reduction of aflatoxins in a traditional African maize-based fermented food. Nutrients 2019, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Oluwafemi, F.; Da-Silva, F.A. Removal of aflatoxins by viable and heat-killed Lactobacillus species isolated from fermented maize. J. Appl. Biosci. 2009, 16, 871–876. [Google Scholar]
- Ezekiel, C.N.; Abia, W.A.; Ogara, I.M.; Sulyok, M.; Warth, B.; Krska, R. Fate of mycotoxins in two popular traditional cereal-based beverages (kunu-zaki and pito) from rural Nigeria. LWT-Food Sci. Technol. 2015, 60, 137–141. [Google Scholar] [CrossRef]
- Nyamete, F.A.; Mourice, B.; Mugula, J.K. Fumonisin B 1 reduction in lactic acid bacteria fermentation of maize porridges. Tanzania. J. Agric. Sci. 2016, 15, 13–20. [Google Scholar]
- Nyamete, F.A.; Bennink, M.; Mugula, J.K. Potential of lactic acid fermentation in reducing aflatoxin B1 in Tanzania maize-based gruel. Afr. J. Food Agric. Nutr. Dev. 2016, 16, 11139–11151. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Song, K.B. In vitro degradation of zearalenone by Bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef] [PubMed]
- Haskard, C.A.; El-Nezami, H.S.; Kankaanpää, P.E.; Salminen, S.; Ahokas, J.T. Surface binding of aflatoxin B1 by lactic acid bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control. 2019, 106, 106731. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name of Product | Country | Food Category | Fermentation Time–Temperature | pH | Ethanol Content (v/v %) | References |
|---|---|---|---|---|---|---|
| Akamu | Nigeria | Porridge | 72 h, 28–30 °C | 3.2–3.9 | - | [1,2,3] |
| Atole agrio | Mexico | Beverage | 6–12 h, 34 °C | 3.9 | - | [4,5] |
| Busaa | Kenya, Nigeria, Ghana | Beverage | 5–7 d + 2–3 d, room T | 3.5–4.0 | 3.5–4.8 | [6,7,8] |
| Champús | Colombia, Ecuador, Perù | Beverage | 24–48 h, 12–15 °C | 3.5–4.0 | 2.5–4.2 | [9,10] |
| Chicha | Argentine | Beverage | 3–7 d + 2 d, room T | 3.6–3.8 | 2.0–12.0 | [11,12] |
| Chicha | Colombia | Beverage | 2–6 d, 18–32 °C | 3.5–4.6 | 2.0–12.0 | [10] |
| Doklu | Côte d’Ivoire | Dough | 2–4 d, room T | 2.2–3.8 | - | [13,14,15] |
| Gowé | Benin | Dough | 16 h, room T | 3.6–4.1 | - | [16,17,18] |
| Ilambazi lokubilisa | Zimbabwe | Porridge | 2–4 d, room T | n.a. | - | [19] |
| Ikii | Kenya | Porridge | n.a. | 3.9 | - | [20,21] |
| Kachasu | Zimbabwe | Beverage | 4–7 d, room T | n.a. | 9.0–41.0 | [7,19,22] |
| Kenkey | Ghana | Dough | 2–4 d, room T | 3.7 | - | [23,24,25,26] |
| Koko | Ghana | Porridge | 2 d, room T | n.a. | - | [7,27] |
| Kokoro | Nigeria | Snack | 24 h, 27–31 °C | 6.2 | - | [28,29] |
| Kutukutu | Cameroon | Dough | 120 h, 25–30 °C | 2.7–3.1 | - | [30] |
| Mahewu/Amahewu | South Africa, Arabian gulf countries | Beverage | 24–72 h, room T | 3.5–3.6 | - | [31,32,33,34,35] |
| Masa agria | Colombia | Dough | 3–5 d, 35–40 °C | 3.1–4.4 | - | [10,36,37] |
| Massa | Nigeria | Snack | 12–24 h, room T | n.a. | - | [38] |
| Mawè | Benin, Togo | Dough | 72 h, 28–32 °C | 3.8–4.2 | - | [16,39,40,41] |
| Munkoyo | Katanga, Zambia, Southern Democratic Republic of Congo | Beverage | 24–48 h, 25–30 °C | 3.5–4.0 | 0.0–2.7 | [42,43,44] |
| Mutwiwa | Zimbabwe | Porridge | room T | n.a. | - | [19] |
| Ogi | Nigeria | Porridge | 48–72 h + 24–48 h, 28–30 °C | 3.8–4.1 | - | [16,45,46,47,48,49] |
| Pito | Nigeria | Beverage | 12 h + 12 h, room T | 4.9 | 3.0–4.0 | [50,51] |
| Poto poto | Congo | Dough | 55 h + 10–11 h, room T | 3.7–3.8 | - | [52,53,54] |
| Pozol | Mexico | Dough | 2–7 d, room T | 4.2–4.6 | - | [55,56,57,58,59,60] |
| Sekete | Nigeria | Beverage | 2–3 d, room T | 2.8–4.3 | 0.9–4.0 | [61,62] |
| Tesguino | Mexico | Beverage | 2–3 d, room T | n.a. | 3.7 | [63] |
| Togwa | Tanzania | Beverage | 12–24 h, room T | 3.1–3.3 | - | [64,65] |
| Name of Product | Lactic Acid Bacteria | Yeasts | References |
|---|---|---|---|
| Akamu | L. lactis, Lb. acidophilus, Lb. amylovorus, Lb. delbrueckii, Lb. fermentum, Lb. helveticus, Lb. plantarum | C. albicans, C. tropicalis, Candida utilis, Clavispora lusitaniae, Rhodotorula glutinis, Saccharomyces paradoxus | [1,2] |
| Atole agrio | E. asini, E. casseliflavus, E. faecium, E. hirae, E. mundtii, L. lactis, L. piscium, Lb. aviarius, Lb. brevis, Lb. casei, Lb. composti, Lb. coryniformis, Lb. curvatus, Lb. dixtrinicus, Lb. mali, Lb. fabifermentans, Lb. paracasei, Lb. paraplantarum, Lb. plantarum, Lb. pentosus, Lb. rhamnosus, Lc. garlicum, Lc. mesenteroides, Lc. pseudomesenteroides, W. cibaria, W. confusa, W. paramesenteroides, P. stilesii, S. equines, W. hellenica, W. oryzae, P. pentosaceus | u.s. | [4,5] |
| Busaa | Lb. brevis, Lb. buchneri, Lb. casei, Lb. helveticus, Lb. plantarum, Lb. salivarius, Lb. viridescens, P. damnosus, P. parvulus | Candida krusei, Saccharomyces cerevisiae | [6,7] |
| Champús | u.s. | Galactomyces geotrichum, Hanseniaspora sp., Issatchenkia orientalis, Pichia fermentans, P. kluyveri, Saccharomyces cerevisiae, Torulospora delbruekii, Zygosaccharomyces fermentati | [9,10] |
| Chicha | E. durans, E. faecalis, E. faecium, E. gallinarum, E. hirae, E. lactis, E. mundtii, L. lactis, Lb. acidophilus, Lb. aviarius, Lb. brevis, Lb. casei, Lb. composti, Lb. crispatus, Lb. diolivorans, Lb. fabifermentans, Lb. farraginis, Lb. fermentum, Lb. harbinensis, Lb. helveticus, Lb. murinus, Lb. odoratitofui, Lb. paracasei, Lb. paraplantarum, Lb. plantarum, Lb. reuteri, Lb. rossiae, Lb. suebicus, Lb. vaccinostercus, Lc. citreum, Lc. lactis, Lc. mesenteroides, Lc. pseudomesenteroides, S. equinus, S. gallolyticus W. cibaria, W. confusa, W.hellenica, W. viridescens | Candida parapsilosis, C. zeylanoides, Cryptococcus carnescens, Cry. flavescens, Cry. magnus, Cry. nemorosus, Hanseniaspora uvarum, Debaryomyces hansenii, Kluyveromyces lactis, K. marxianus, Meyerozyma guilliermondii, Pichia sp., P. fermentans, P. membranifaciens, Rhodotorula mucilaginosa, R. slooffiae, S. cerevisiae, Torulaspora delbrueckii, Wickerhamomyces anomalus, Trichosporon domesticum | [11,12] |
| Chicha | Lactobacillus sp., Leuconostoc sp. | Candida ethanolica, C. oleophila, C. parapsilosis, C. pomicola, C. railenensis, C. sergipensis, C. spandovensis, Hanseniaspora opuntiae, H. uvarum, Issatchenkia sp., Kazachstania exigua, Kodamaea ohmeri, Lodderomyces elongisporus, Metschnikowia koreensis, Monilia candida, Mycoderma vini, Oidium lactis, Pichia sp., P. guilliermondii, Saccharomyces cerevisiae, S. pastorianus, Wickerhamomyces anomalus, W. pijperi | [10] |
| Chica | Lactobacillus plantarum, Lb. fermentum, Weissella cibaria, Leuconostoc sp., Lactococcus sp., S. luteciae, S. alactolyticus | [74] | |
| Doklu | Enterococcus sp., Lactobacillus sp., Lb. fermentum, Lb. plantarum, Pediococcus sp., P. acidilactici, P. pentosaceus, Streptococcus sp., Weissella sp., W. cibaria | u.s. | [13,15] |
| Gowé | u.s. | u.s. | [18] |
| Ikii | Lb. confusus, Lb. fermentum, Lb. plantarum, Lb. rhamnosus, Pediococcus sp. | u.s. | [20,21] |
| Kachasu | - | u.s. | [7] |
| Kenkey | Lb. fermentum, Lb. reuteri | Candida kefir, C. krusei, C. mycoderma, C. tropicalis, Saccharomyces cerevisiae | [23] |
| Koko | Lb. brevis, Lb. plantarum | Saccharomyces cerevisiae | [7] |
| Kokoro | Lactobacillus sp. | - | [28] |
| Mahewu/amahewu | Leuconostoc spp., L. lactis, Lb. delbrueckii, S. lactis | - | [31,32,34] |
| Masa agria | Lactococcus sp., L. lactis, Lactobacillus sp., Lb. amylolyticus, Lb. brevis, Lb. coleohominis, Lb. delbrueckii, Lb. crustorum, Lb. curvatus, Lb. fermentum, Lb. gallinarum, Lb. helveticus, Lb. nagelii, Lb. nantensis, Lb. panis, Lb. plantarum, Lb. pontis, Lb. rossiae, Lb. siliginis, Lb. vaccinostercus, Lc. citreum, P. argentinicus, Streptococcus sp., Weissella sp., W. beninensis, W. confusa, W. fabalis, W. fabaria, W. salipiscis | u.s. | [10,36] |
| Massa | Lb. fermentum, Lb. lactis, Lb. plantarum, Lc. mesenteroides, Pediococcus acidilactici | u.s. | [38] |
| Mawè | L. lactis, Lb. brevis, Lb. buchneri, Lb. confusus, Lb. curvatus, Lb. fermentum, Lb. reuteri, Lb. salivarius, Lc. mesenteroides, Pediococcus acidilactici, P. pentosaceus | Candida krusei, Clavispora lusitaniae, Saccharomyces cerevisiae | [16,39,41] |
| Munkoyo | Lb. brevis, Lb. delbruekii, Lb. fermentum, Lb. plantarum, Lb. rossiae, W. cibaria, W. confusa | Saccharomyces cerevisiae | [43,44] |
| Mutwiwa | Pediococcus pentosaceus | u.s. | [19] |
| Ogi | E. faecalis, Lb. acidophilus, Lb. brevis, Lb. cellobiosus, Lb. fermentum, Lb. paraplantarum, Lb. plantarum, Lc. lactis, Lc. paramesenteroides, P. acidilactici, P. claussenii, P. pentosaceus, S. lactis | Candida albicans, C. krusei, C. tropicalis, C. utilis, Clavispora lusitaniae, Geotrichum candidum, G. fermentans, Rhodotorula glutinis, R. graminis, Saccharomyces cerevisiae, S. pastorianus | [16,45,46,48,49,75] |
| Pito | Lactobacillus sp. | Candida sp., Geotrichum candidum | [50] |
| Poto poto | Enterococcus sp., Lb. acidophilus, Lb. casei, Lb. delbrueckii, Lb. fermentum, Lb. gasseri, Lb. plantarum, Lb. reuteri | - | [53,54] |
| Pozol | Bifidobacterium minimum, Enterococcus sp., E. saccharolyticus, E. sulfureus, Lactococcus sp., L. lactis, Lactobacillus sp., Lb. alimentarius, Lb. casei, Lb. delbrueckii, Lb. fermentum, Lb. plantarum, Leuconostoc sp., Streptococcus sp., S. bovis, S. macedonicus, S. suis, Weissella sp. | u.s. | [55,56,57,58,59,60] |
| Sekete | L. lactis, Lb. brevis, Lb. delbrueckii, Lb. plantarum, Lc. mesenteroides, P. cerevisiae, Streptococcus spp. | Geotrichum sp., Saccharomyces spp., S. cerevisiae | [7,62,76] |
| Tesguino | Lactobacillus sp., Leuconostoc sp., Pediococcus sp., Streptococcus sp. | Brettanomyces sp., Candida guilliermondii, Cryptococcus sp., Geotrichum sp., Hansenula anomala, Pichia sp., Saccharomyces cerevisiae, S. kluyveri | [63] |
| Togwa | Lb. brevis, Lb., cellobiosus, Lb. fermentum, Lb. plantarum, P. pentosaceus, W. confusa | Candida glabrata, C. pelliculosa, C. tropicalis, Issatchenkia orientalis, Kluyveromyces marxianus, Pichia anomala, Saccharomyces cerevisiae | [64,65] |
| Name of Product | Acetic Acid Bacteria | Enterobacteriaceae | Molds | Others | References |
|---|---|---|---|---|---|
| Akamu | - | Enterobacter aerogenes, Escherichia coli, Proteus sp., Serratia sp. | Aspergillus niger, A. oryzae, Mucor circinelloides, Penicillium citrinum, Rhizopus microsporus, R. oligosporus | Bacillus cereus, B. licheniformis, B. subtilis, Pseudomonas alkaligenes, P. aeruginosa | [1] |
| Atole agrio | Acetobacter estunensis, A. indonesiensis, A. pasteurianus, A. tropicalis, Gluconacetobacter sp., Gluconobacter sp., G. frateurii, Kozakia sp. | u.s. | u.s. | - | [4,5] |
| Chicha | - | - | Penicillium sp. | - | [12] |
| Chicha | Acetobacter sp. | - | Aspergillus sp., Penicillium sp. | - | [10] |
| Koko | - | Enterobacter cloacae | - | Acinetobacter sp. | [7] |
| Kokoro | - | Klebsiella sp., Proteus sp. | Alternaria sp., Aspergillus sp., Cephalosporium sp., Fusarium sp., Mucor sp., Penicillium sp., Rhizopus sp. | Bacillus sp., Pseudomonas sp., Staphylococcus sp. | [28] |
| Masa agria | Acetobacter sp., A. cibinongensis, A. fabarum, A. lovaniensis, A. orientalis, Gluconobacter oxydans | Enterobacter aerogenes, Escherichia sp., Pantoea agglomerans, Serratia sp. | - | Acinetobacter sp., A. junii, A. ursingii, Bacteroides sp., Comamonas terrigena, Dechloromonas sp., Delftia, Frateuria aurantia, Gemmata sp., Pseudomonas sp., Sphingobium sp., Sphingomonas sp., Stenotrophomonas bacterium, Sugarcane phytoplasma | [10,36] |
| Munkoyo | - | - | - | Bacillus licheniformis | [44] |
| Ogi | - | Citrobacter sp., Enterobacter sp., Escherichia coli, Klebsiella spp. | Aspergillus flavus, A. niger, A. oryzae, Fusarium subglutinans, Mucor circinelloides, Penicillium citrinum, Rhizopus microsporus, R. nigrigans, R. oligosporus, R. stolonifer | Acinetobacter berezinae, Aerobacter sp., Alcaligenes faecalis, Bacillus cereus, B. licheniformis, B. mycoides, B. subtilis, B. thuringiensis, Bordetella avium, B. bronchisepta, Corynebacterium sp., Micrococcus luteus, Moorella glycerini, Myroides marinus, Pseudomonas aeruginosa, P. hibiscicola, S. aureus | [46,48,49,75] |
| Pito | - | - | Aspergillus versicolor, Penicillium purpurogenum, P. simplicissimum | - | [50] |
| Poto poto | - | Escherichia coli | - | Bacillus sp. | [53] |
| Pozol | - | u.s. | u.s. | Bacillus sp., Clostridium sp, Exiguobacterium acetylicum, E. aurantiacum, Oxalophagus oxalicus | [55,56,57,59,60] |
| Sekete | - | - | Aspergillus flavus, A. niger, Mucor rouxii | Bacilus subtilis, Propionibacterium spp. | [7,62,76] |
| Mycotoxin | Detoxifying Microorganism | Reduction (%) | Strain Origin | Place of Fermentation | Reference |
|---|---|---|---|---|---|
| Aflatoxin B1 | Indigenous microbial communities | 40–60.8 | Ogi | Ogi | [55] |
| 27.5 | Maize meal | Maize meal | [153] | ||
| Lb. brevis | 63 | Kutukutu | Kutukutu | [30] | |
| Lb. buchneri | 64.2 | Kutukutu | Kutukutu | [30] | |
| Lb. rhamnosus and S. thermophilus | 92–100 | Commercial strains | Kwete | [154] | |
| S. lactis and Lb. delbrueckii | 75 | Commercial strains | Maize meal | [145] | |
| Aflatoxin B2 | Indigenous microbial communities | 68–82.8 | Ogi | Ogi | [55] |
| Lb. rhamnosus and S. thermophilus | 91.8–100 | Commercial strains | Kwete | [154] | |
| Aflatoxin M1 | Indigenous microbial communities | 100 | Ogi | Ogi | [55] |
| Aflatoxins | Indigenous microbial communities | 80 | Ogi | Ogi | [153] |
| ≥91 | Mawe | Mawe | [153] | ||
| Lb. acidophilus | 37.5 | Ogi | Maize | [155] | |
| Lb. brevis | 75 | Ogi | Maize | [155] | |
| Lb. casei | 62.5 | Ogi | Maize | [155] | |
| Lb. delbrueckii | 56.25 | Ogi | Maize | [155] | |
| Lb. plantarum | 95 | Ogi | Maize | [155] | |
| Alternariol | Indigenous microbial communities | 96.7 | Kunu-zaki | Kunu-zaki | [156] |
| Alternariolmethylether | Indigenous microbial communities | 96 | Kunu-zaki | Kunu-zaki | [156] |
| Beauvericin | Indigenous microbial communities | 99.9 | Kunu-zaki | Kunu-zaki | [156] |
| Citrinin | Indigenous microbial communities | 33–100 | Ogi | Ogi | [75] |
| Cyclopiazonic acid | Indigenous microbial communities | 98.1–100 | Ogi | Ogi | [75] |
| Deoxynivalenol | Indigenous microbial communities | 98.9 | Kunu-zaki | Kunu-zaki | [156] |
| Enniatins | Indigenous microbial communities | 94.7 | Kunu-zaki | Kunu-zaki | [156] |
| Fumonisin B1 | Back slopped* | 30 | Maize based porridge | Maize based porridge | [157] |
| 68 | Togwa | Togwa | [158] | ||
| Indigenous microbial communities | 20 | Maize based porridge | Maize based porridge | [157] | |
| 55 | Togwa | Togwa | [158] | ||
| 13–88.8 | Ogi | Ogi | [75] | ||
| 99.4 | Kunu-zaki | Kunu-zaki | [156] | ||
| 24.4 | Maize meal | Maize meal | [159] | ||
| Lb. casei | 17 | n.a. | Maize based porridge | [157] | |
| 52 | n.a. | Togwa | [158] | ||
| Lb. delbrueckii ssp. delbrueckii | 69 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 29 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| Lb. fermentum | 17 | n.a. | Maize based porridge | [157] | |
| 55 | n.a. | Togwa | [158] | ||
| Lb. plantarum | 73 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 8 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| 24 | n.a. | Maize based porridge | [157] | ||
| 55 | n.a. | Togwa | [158] | ||
| Pediococcus pentosaceus | 43 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 19 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| 24 | n.a. | Maize based porridge | [157] | ||
| 45 | n.a. | Togwa | [158] | ||
| S. lactis and Lb. delbrueckii | 74.6 | Commercial strains | Maize meal | [159] | |
| Fumonisin B2 | Indigenous microbial communities | 44–80 | Ogi | Ogi | [75] |
| Lb. delbrueckii ssp. delbrueckii | 95 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [150] | |
| 6 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [150] | ||
| Lb. delbrueckii ssp. bulgaricus | 55 | n.a. | in vitro: corn infusion, 24 h, 25°C, pH 4 | [152] | |
| Lb. plantarum | 95 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [143] | |
| 21 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [143] | ||
| Lb. rhamnosus | 80 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Leuconostoc mesenteroides | 65 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Pediococcus pentosaceus | 89 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [150] | |
| 67 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [150] | ||
| Fumonisin B3 | Indigenous microbial communities | 46–95 | Ogi | Ogi | [55] |
| Fumonisins | Indigenous microbial communities | 99.5 | Kunu-zaki | Kunu-zaki | [148] |
| 29 | Ogi | Ogi | [146] | ||
| ≥87 | Mawe | Mawe | [146] | ||
| Fusaproliferin | Indigenous microbial communities | 97.8 | Kunu-zaki | Kunu-zaki | [148] |
| Moniliformin | Indigenous microbial communities | 98.3 | Kunu-zaki | Kunu-zaki | [148] |
| Zearalenone | Indigenous microbial communities | 79–100 | Ogi | Ogi | [75] |
| 76.2 | Kunu-zaki | Kunu-zaki | [156] | ||
| 34.3 | Maize meal | Maize meal | [159] | ||
| Lb. delbrueckii ssp. bulgaricus | 75 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Lb. rhamnosus | 80 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Leuconostoc mesenteroides | 75 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| S. lactis and Lb. delbrueckii | 68.2 | Commercial strains | Maize meal | [159] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves-López, C.; Rossi, C.; Maggio, F.; Paparella, A.; Serio, A. Changes Occurring in Spontaneous Maize Fermentation: An Overview. Fermentation 2020, 6, 36. https://doi.org/10.3390/fermentation6010036
Chaves-López C, Rossi C, Maggio F, Paparella A, Serio A. Changes Occurring in Spontaneous Maize Fermentation: An Overview. Fermentation. 2020; 6(1):36. https://doi.org/10.3390/fermentation6010036
Chicago/Turabian StyleChaves-López, Clemencia, Chiara Rossi, Francesca Maggio, Antonello Paparella, and Annalisa Serio. 2020. "Changes Occurring in Spontaneous Maize Fermentation: An Overview" Fermentation 6, no. 1: 36. https://doi.org/10.3390/fermentation6010036