Adaptive Evolution of Industrial Brewer’s Yeast Strains towards a Snowflake Phenotype


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Media
2.2. Flocculation Assay
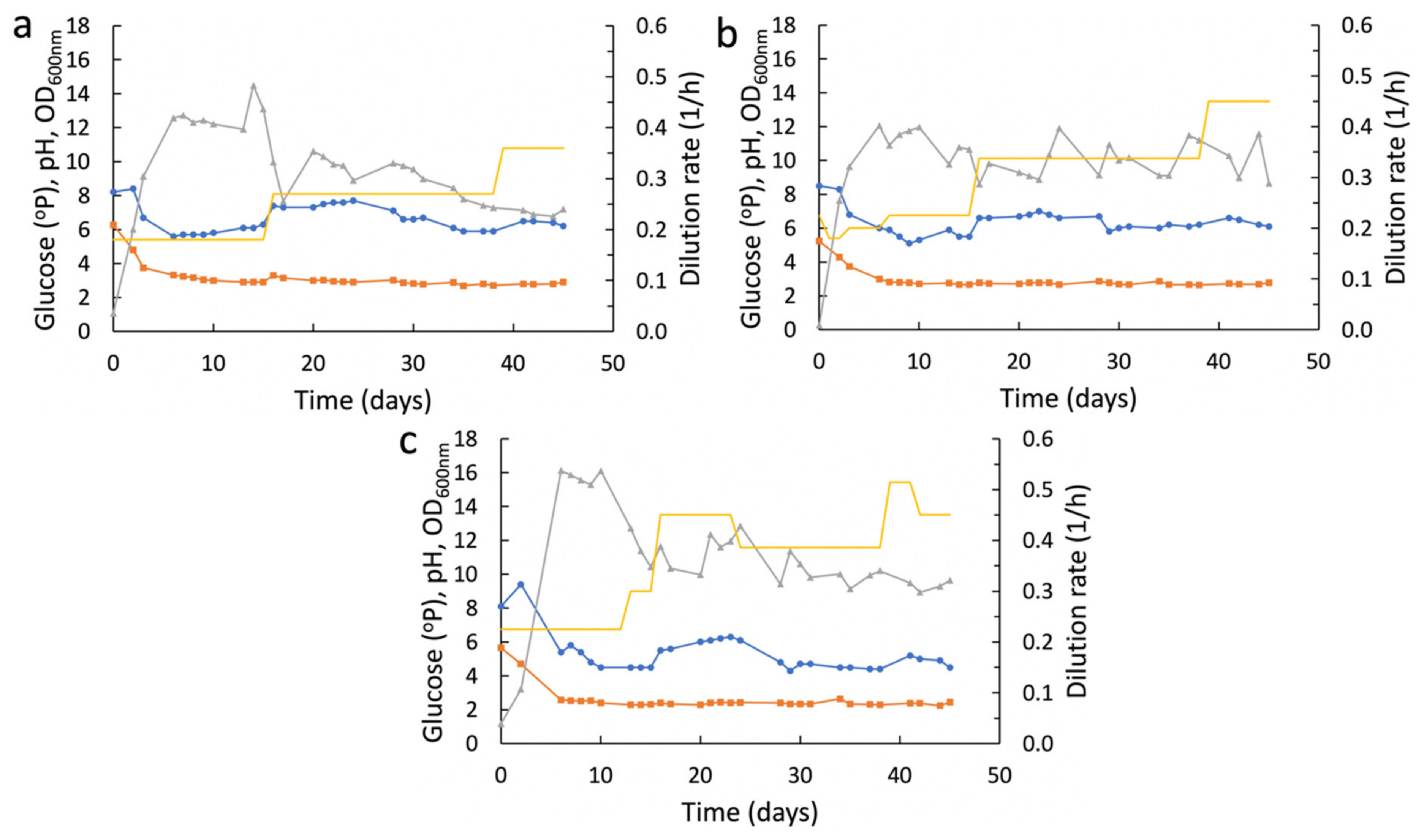
2.3. Experimental Setup with Ministats
2.4. Wort Fermentations in Tall Tubes
3. Results
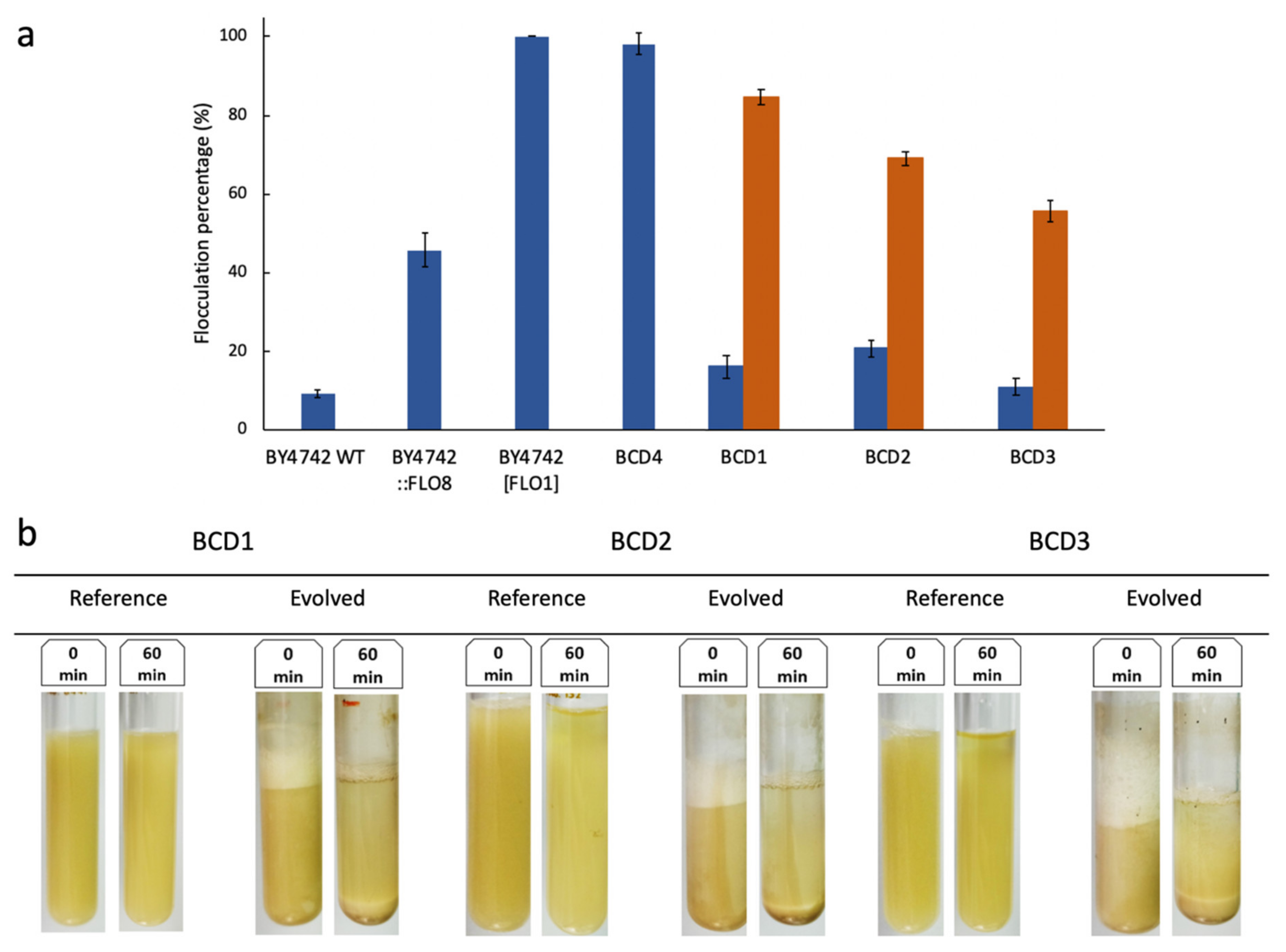
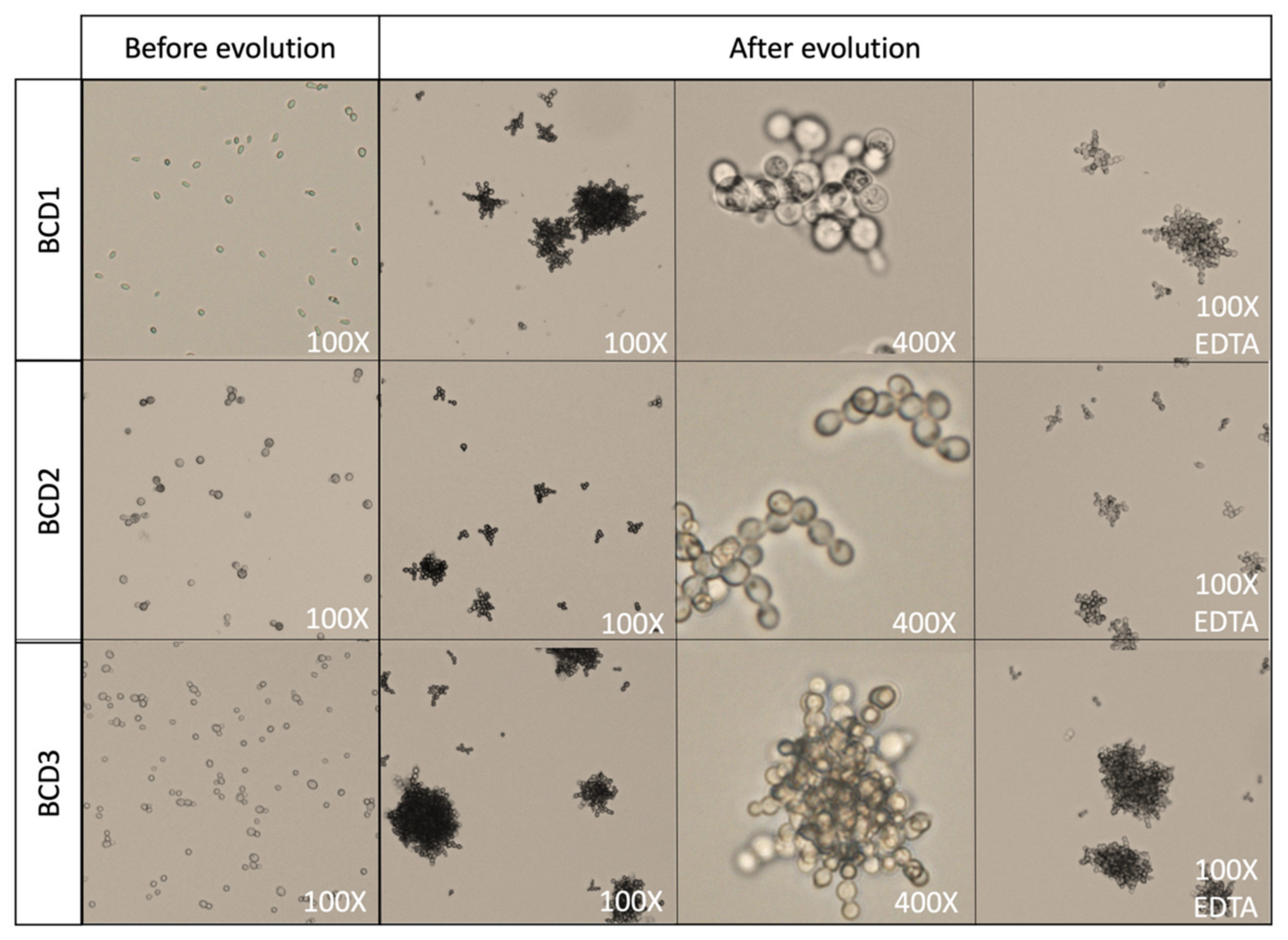
3.1. Adaptive Evolution in Ministats
3.2. Performance of Evolved Strains During Beer Fermentation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bowden, C.P.; Leaver, G.; Melling, J.; Norton, M.G.; Whittington, P.N. Recent and novel developments in the recovery of cells from fermentation broths. In Separations for Biotechnology; Verral, M.S., Hudson, M.J., Eds.; Ellis Horwood: Chichester, UK, 1987; pp. 49–61. [Google Scholar]
- Stratford, M. Yeast flocculation: A new perspective. Adv. Microb. Physiol. 1992, 33, 1–71. [Google Scholar]
- Vidgren, V.; Londesborough, J. 125th anniversary review: Yeast flocculation and sedimentation in brewing. J. Inst. Brew. 2011, 117, 475–487. [Google Scholar] [CrossRef]
- Goossens, K.; Willaert, R. Flocculation protein structure and cell-cell adhesion mechanism in Saccharomyces cerevisiae. Biotechnol. Lett. 2010, 32, 1571–1585. [Google Scholar] [CrossRef] [Green Version]
- Van Mulders, S.E.; Christianen, E.; Saerens, S.M.G.; Daenen, L.; Verbelen, P.J.; Willaert, R.; Verstrepen, K.J.; Delvaux, F.R. Phenotypic diversity of Flo protein family-mediated adhesion in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 178–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willaert, R.G. Adhesins of yeasts: Protein structure and interactions. J. Fungi 2018, 4, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brückner, S.; Mösch, H.-U. Choosing the right lifestyle: Adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2011, 36, 25–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Yeast flocculation: What brewers should know. Appl. Microbiol. Biotechnol. 2003, 61, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.R.; Lawrence, S.J.; Leclaire, J.P.R.; Powell, C.D.; Smart, K.A. Yeast Responses to Stresses Associated with Industrial Brewery Handling. FEMS Microbiol. Rev. 2007, 31, 535–569. [Google Scholar] [CrossRef] [Green Version]
- Soares, E.V. Flocculation in Saccharomyces cerevisiae: A review. J. Appl. Microbiol. 2010, 110, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Dequin, S. The potential of genetic engineering for improving brewing, wine-making and baking yeasts. Appl. Microbiol. Biotechnol. 2001, 56, 577–588. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Duong, C.T.; Nevoigt, E. Genetic improvement of brewer’s yeast: Current state, perspectives and limits. Appl. Microbiol. Biotechnol. 2010, 86, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Conrad, T.M.; Lewis, N.E.; Palsson, B.Ø. Microbial laboratory evolution in the era of genome-scale science. Mol. Syst. Biol. 2011, 7, 509. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, V.A.; Bezdan, D.; Zengler, K. Adaptive laboratory evolution-harnessing the power of biology for metabolic engineering. Curr. Opin. Biotechnol. 2011, 22, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Dragosits, M.; Mattanovich, D. Adaptive laboratory evolution—Principles and applications for biotechnology. Microb. Cell Fact. 2013, 12, 64. [Google Scholar] [CrossRef] [Green Version]
- Sonderegger, M.; Sauer, U. Evolutionary engineering of Saccharomyces cerevisiae for anaerobic growth on xylose. Appl. Environ. Microbiol. 2003, 69, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Dhar, R.; Sägesser, R.; Weikert, C.; Wagner, A. Yeast adapts to a changing stressful environment by evolving cross-protection and anticipatory gene regulation. Mol. Biol. Evol. 2013, 30, 573–588. [Google Scholar] [CrossRef] [Green Version]
- Ekberg, J.; Rautio, J.; Mattinen, L.; Vidgren, V.; Londesborough, J.; Gibson, B.R. Adaptive evolution of the lager brewing yeast Saccharomyces pastorianus for improved growth under hyperosmotic conditions and its influence on fermentation performance. FEMS Yeast Res. 2013, 13, 335–349. [Google Scholar] [CrossRef] [Green Version]
- Strejc, J.; Siříšťová, L.; Karabín, M.; Almeida e Silva, J.B.; Brányik, T. Production of alcohol-free beer with elevated amounts of flavouring compounds using lager yeast mutants. J. Inst. Brew. 2013, 119, 149–155. [Google Scholar] [CrossRef]
- Huuskonen, A.; Markkula, T.; Vidgren, V.; Lima, L.; Mulder, L.; Geurts, W.; Walsh, M.; Londesborough, J. Selection from Industrial Lager Yeast Strains of Variants with Improved Fermentation Performance in Very-High-Gravity Worts. Appl. Environ. Microbiol. 2010, 76, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Zhao, H.; Li, H.; Zhang, Q.; Lei, H.; Zhao, M. Selection of Saccharomyces pastorianus variants with improved fermentation performance under very high gravity wort conditions. Biotechnol. Lett. 2012, 34, 365–370. [Google Scholar] [CrossRef]
- Cadière, A.; Ortiz-Julien, A.; Camarasa, C.; Dequin, S. Evolutionary engineered Saccharomyces cerevisiae wine yeast strains with increased in vivo flux through the pentose phosphate pathway. Metab. Eng. 2011, 13, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Cadière, A.; Aguera, E.; Caillé, S.; Ortiz-Julien, A.; Dequin, S. Pilot-scale evaluation the enological traits of a novel, aromatic wine yeast strain obtained by adaptive evolution. Food Microbiol. 2012, 32, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Weikert, C.; Sauer, U.; Bailey, J.E. Use of a glycerol-limited, long-term chemostat for isolation of Escherichia coli mutants with improved physiological properties. Microbiology 1997, 143, 1567–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppram, R.; Albers, E.; Olsson, L. Evolutionary engineering strategies to enhance tolerance of xylose utilizing recombinant yeast to inhibitors derived from spruce biomass. Biotechnol. Biofuels 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Spira, B.; Zhou, Z.; Feng, L.; Maharjan, R.P.; Li, X.; Li, F.; McKenzie, C.; Reeves, P.R.; Ferenci, T. Divergence involving global regulatory gene mutations in an Escherichia coli population evolving under phosphate limitation. Genome Biol. Evol. 2010, 2, 478–487. [Google Scholar] [CrossRef]
- Conjaerts, A.; Willaert, R. Gravity-Driven Adaptive Evolution of an Industrial Brewer’s Yeast Strain towards a Snowflake Phenotype in a 3D-Printed Mini Tower Fermentor. Fermentation 2017, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.W.; Befort, C.; Kerr, E.O.; Dunham, M.J. Design and use of multiplexed chemostat arrays. J. Vis. Exp. 2013, e50262. [Google Scholar] [CrossRef]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer Deletion Strains derived from Saccharomyces cerevisiae S288C: A Useful set of Strains and Plasmids for PCR-mediated Gene Disruption and Other Applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- D’Hautcourt, O.; Smart, K.A. Measurement of brewing yeast flocculation. J. Am. Soc. Brew. Chem. 1999, 57, 123–128. [Google Scholar] [CrossRef]
- Shimizu, H.; Mizuno, S.; Hiroshima, T.; Shioya, S. Effect of carbon and nitrogen additions on consumption activity of apparent extract of yeast cells in a brewing process. J. Am. Soc. Brew. Chem. 2002, 60, 163–169. [Google Scholar] [CrossRef]
- Verbelen, P.J.; Dekoninck, T.M.L.; Saerens, S.M.G.; Van Mulders, S.E.; Thevelein, J.M.; Delvaux, F.R. Impact of pitching rate on yeast fermentation performance and beer flavour. Appl. Microbiol. Biotechnol. 2009, 82, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Dekoninck, T.M.L.; Mertens, T.; Delvaux, F.; Delvaux, F.R. Influence of beer characteristics on yeast refermentation performance during bottle conditioning of belgian beers. J. Am. Soc. Brew. Chem. 2013, 71, 23–34. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Fankhauser, J.D.; Rogers, D.W.; Greig, D.; Travisano, M. Origins of multicellular evolvability in snowflake yeast. Nat. Commun. 2015, 6, 6102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratcliff, W.C.; Denison, R.F.; Borrello, M.; Travisano, M. Experimental evolution of multicellularity. Proc. Natl. Acad. Sci. USA 2012, 109, 1595–1600. [Google Scholar] [CrossRef] [Green Version]
- Oud, B.; Guadalupe-Medina, V.; Nijkamp, J.F.; De Ridder, D.; Pronk, J.T.; Van Maris, A.J.A.; Daran, J.M. Genome duplication and mutations in ACE2 cause multicellular, fast-sedimenting phenotypes in evolved Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2013, 110, E4223–E4231. [Google Scholar] [CrossRef] [Green Version]
- Djordjević, V.; Willaert, R.; Gibson, B.; Nedović, V. Immobilized Yeast Cells and Secondary Metabolites. In Fungal Metabolites; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 1–40. [Google Scholar]
- Ratcliff, W.C.; Pentz, J.T.; Travisano, M. Tempo and mode of multicellular adaptation in experimentally evolved Saccharomyces cerevisiae. Evolution 2013, 67, 1573–1581. [Google Scholar] [CrossRef]
- Hope, E.A.; Amorosi, C.J.; Miller, A.W.; Dang, K.; Heil, C.S.; Dunham, M.J. Experimental evolution reveals favored adaptive routes to cell aggregation in yeast. Genetics 2017, 206, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- King, L.; Butler, G. Ace2p, a regulator of CTS1 (chitinase) expression, affects pseudohyphal production in Saccharomyces cerevisiae. Curr. Genet. 1998, 34, 183–191. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Flavor chemistry of beer: Part ii: Flavour and threshold of 239 aroma volatiles. MBAA Tech. Q. 1975, 12, 151–168. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayacan, Y.; Van Mieghem, T.; Delvaux, F.; Delvaux, F.R.; Willaert, R. Adaptive Evolution of Industrial Brewer’s Yeast Strains towards a Snowflake Phenotype. Fermentation 2020, 6, 20. https://doi.org/10.3390/fermentation6010020
Kayacan Y, Van Mieghem T, Delvaux F, Delvaux FR, Willaert R. Adaptive Evolution of Industrial Brewer’s Yeast Strains towards a Snowflake Phenotype. Fermentation. 2020; 6(1):20. https://doi.org/10.3390/fermentation6010020
Chicago/Turabian StyleKayacan, Yeseren, Thijs Van Mieghem, Filip Delvaux, Freddy R. Delvaux, and Ronnie Willaert. 2020. "Adaptive Evolution of Industrial Brewer’s Yeast Strains towards a Snowflake Phenotype" Fermentation 6, no. 1: 20. https://doi.org/10.3390/fermentation6010020