
Downregulation of Exosomal hsa-miR-551b-3p in Obesity and Its Link to Type 2 Diabetes Mellitus

 , ,
, ,
Abstract
:
1. Introduction
2. Results
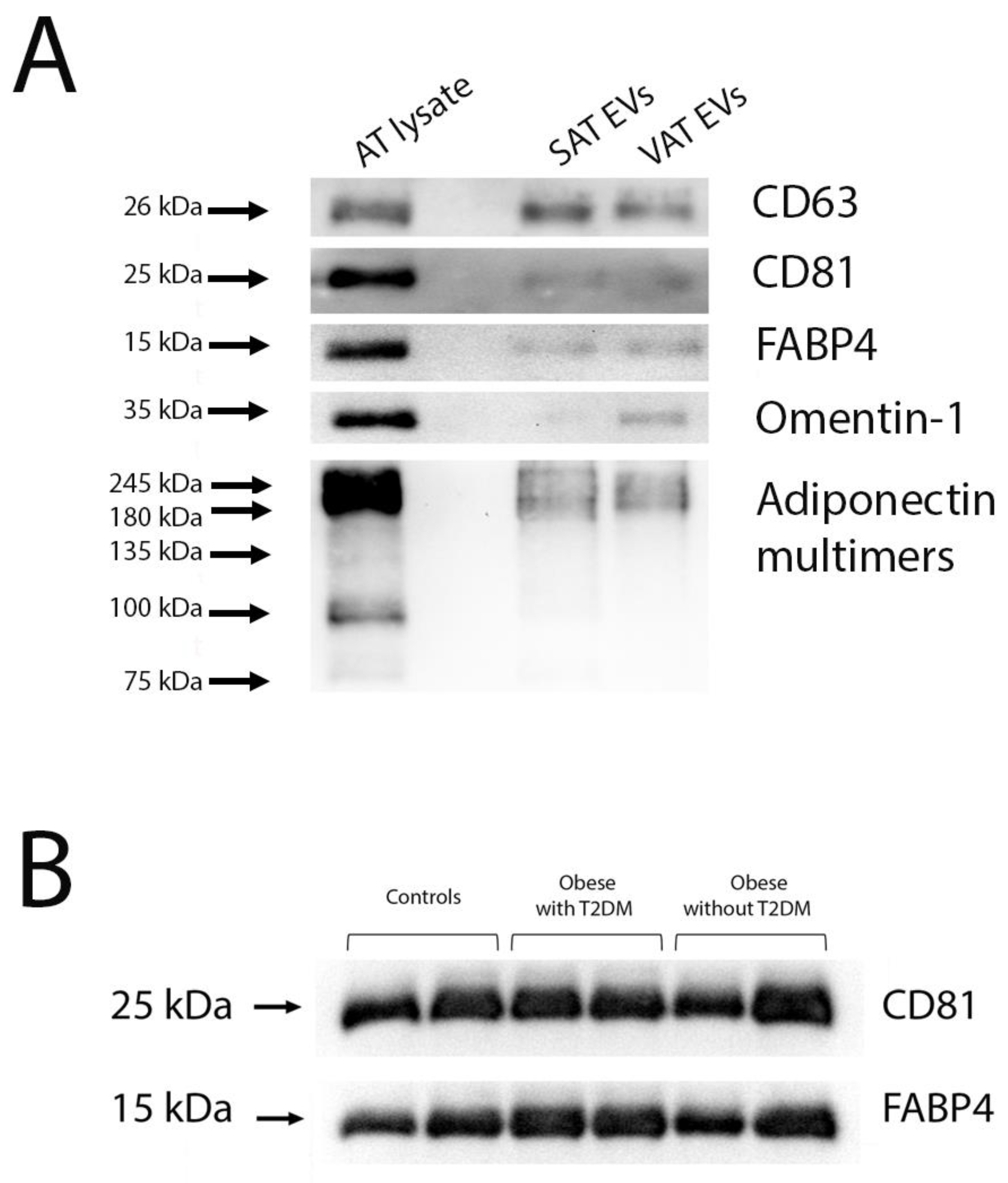
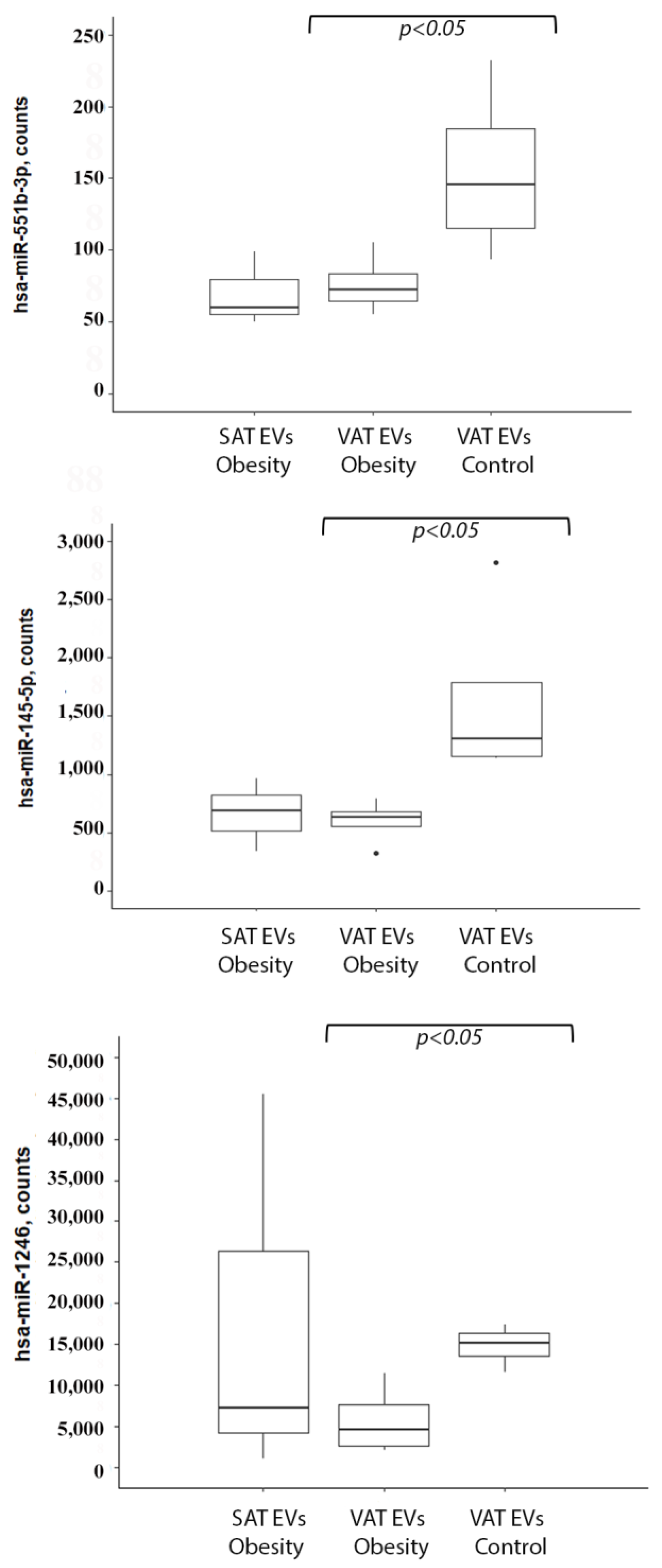
2.1. miRNA Profiling of Adipose Tissue EVs
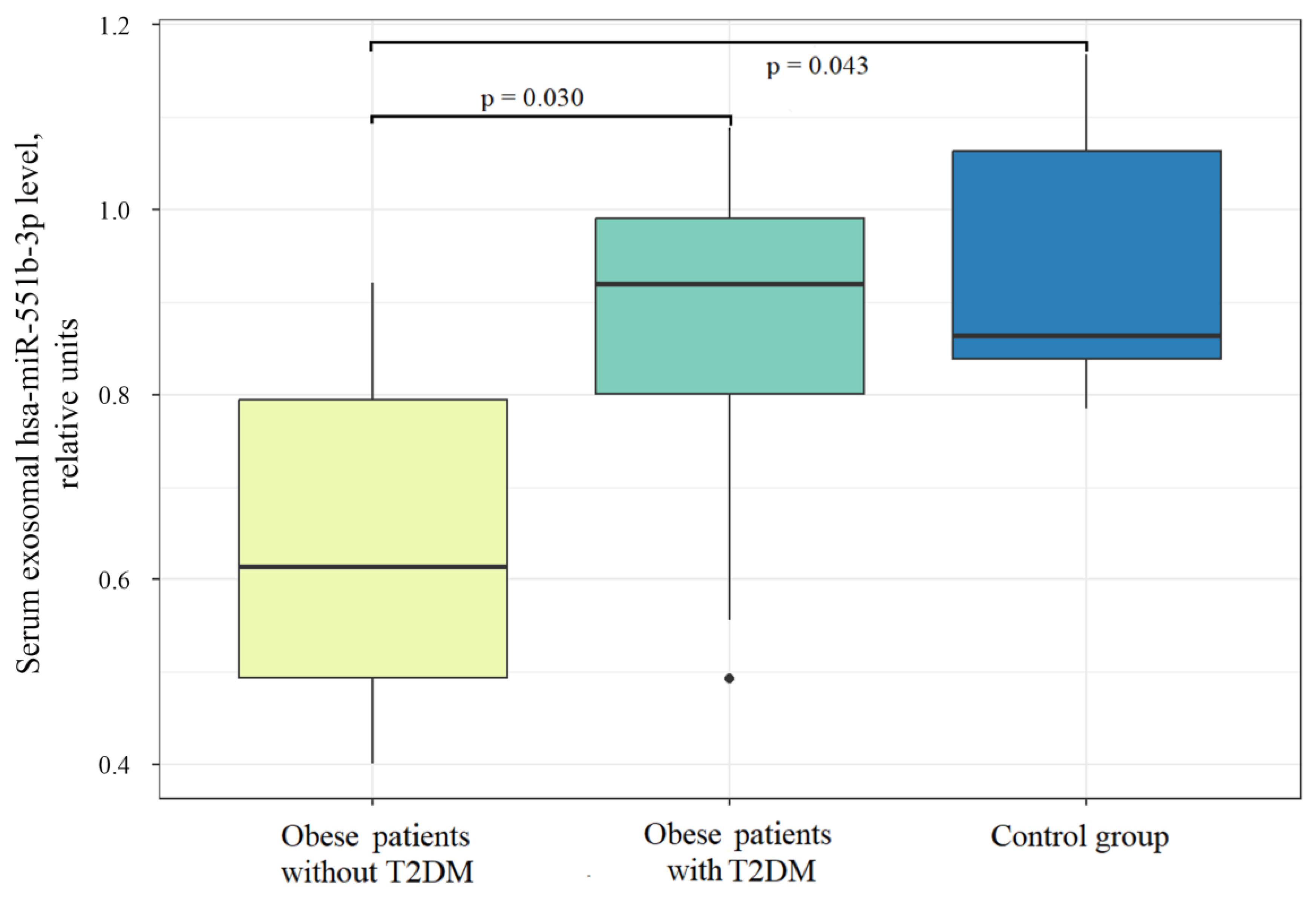
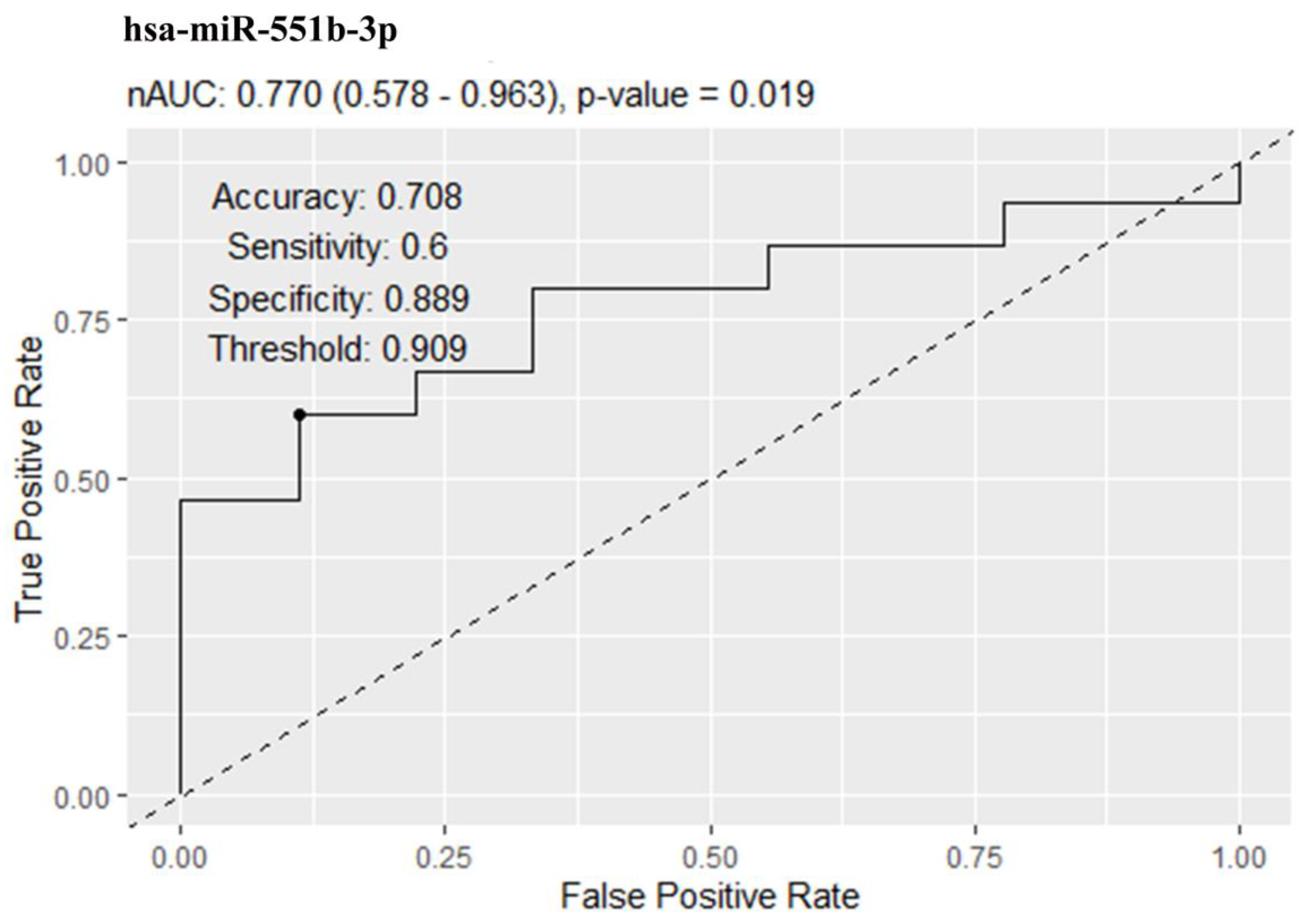
2.2. miRNA Levels in Serum EVs in Obesity and T2DM
3. Discussion
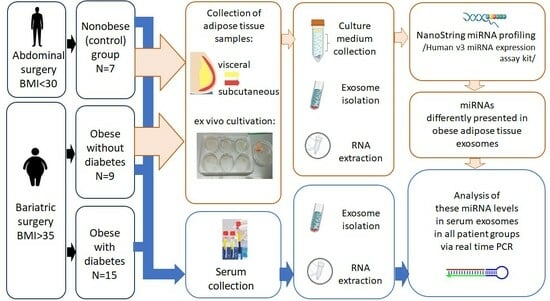
4. Materials and Methods
4.1. Study Participants
4.2. Serum Collection
4.3. Adipose Tissue Cultivation
4.4. Western Blotting
4.5. NanoString
4.6. NanoString Data Normalization and Analysis
4.7. GO and Pathway Enrichment Analysis
4.8. Serum Exosomal miRNA Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruze, R.; Liu, T.; Zou, X.; Song, J.; Chen, Y.; Xu, R.; Yin, X.; Xu, Q. Obesity and type 2 diabetes mellitus: Connections in epidemiology, pathogenesis, and treatments. Front. Endocrinol. 2023, 14, 1161521. [Google Scholar] [CrossRef] [PubMed]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Cione, E.; Cannataro, R.; Gallelli, L.; De Sarro, G.; Caroleo, M.C. Exosome microRNAs in Metabolic Syndrome as Tools for the Early Monitoring of Diabetes and Possible Therapeutic Options. Pharmaceuticals 2021, 14, 1257. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Kuang, G.; Wu, Y.; Ou, C. Emerging roles of exosomal miRNAs in diabetes mellitus. Clin. Transl. Med. 2021, 11, e468. [Google Scholar] [CrossRef]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Raffort, J.; Hinault, C.; Dumortier, O.; Van Obberghen, E. Circulating microRNAs and diabetes: Potential applications in medical practice. Diabetologia 2015, 58, 1978–1992. [Google Scholar] [CrossRef]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef]
- Pardo, F.; Villalobos-Labra, R.; Sobrevia, B.; Toledo, F.; Sobrevia, L. Extracellular vesicles in obesity and diabetes mellitus. Mol. Asp. Med. 2018, 60, 81–91. [Google Scholar] [CrossRef]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Qasim, M.; Khan, K.; Kim, J.-H. Biogenesis, Membrane Trafficking, Functions, and Next Generation Nanotherapeutics Medicine of Extracellular Vesicles. Int. J. Nanomed. 2021, 16, 3357–3383. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal. 2021, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Kwan, H.Y.; Chen, M.; Xu, K.; Chen, B. The impact of obesity on adipocyte-derived extracellular vesicles. Cell Mol. Life Sci. 2021, 78, 7275–7288. [Google Scholar] [CrossRef] [PubMed]
- Miroshnikova, V.V.; Dracheva, K.V.; Kamyshinsky, R.A.; Yastremsky, E.V.; Garaeva, L.A.; Pobozheva, I.A.; Landa, S.B.; Anisimova, K.A.; Balandov, S.G.; Hamid, Z.M.; et al. Cryo-electron microscopy of adipose tissue extracellular vesicles in obesity and type 2 diabetes mellitus. PLoS ONE 2023, 18, e0279652. [Google Scholar] [CrossRef]
- Chandrasekera, D.; Katare, R. Exosomal microRNAs in diabetic heart disease. Cardiovasc. Diabetol. 2022, 21, 122. [Google Scholar] [CrossRef]
- Deng, Z.-B.; Poliakov, A.; Hardy, R.W.; Clements, R.; Liu, C.; Liu, Y.; Wang, J.; Xiang, X.; Zhang, S.; Zhuang, X.; et al. Adipose Tissue Exosome-Like Vesicles Mediate Activation of Macrophage-Induced Insulin Resistance. Diabetes 2009, 58, 2498–2505. [Google Scholar] [CrossRef]
- Fachim, H.A.; Loureiro, C.M.; Siddals, K.; Dalton, C.F.; Reynolds, G.P.; Gibson, J.M.; Chen, Z.B.; Heald, A.H. Circulating microRNA changes in patients with impaired glucose regulation. Adipocyte 2020, 9, 443–453. [Google Scholar] [CrossRef]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA Signatures as Future Biomarkers for Diagnosis of Diabetes States. Cells 2019, 8, 1533. [Google Scholar] [CrossRef]
- Fluitt, M.B.; Mohit, N.; Gambhir, K.K.; Nunlee-Bland, G. To the Future: The Role of Exosome-Derived microRNAs as Markers, Mediators, and Therapies for Endothelial Dysfunction in Type 2 Diabetes Mellitus. J. Diabetes Res. 2022, 2022, 5126968. [Google Scholar] [CrossRef]
- Melesse, D.Y.; Mutua, M.K.; Choudhury, A.; Wado, Y.D.; Faye, C.M.; Neal, S.; Boerma, T. Adolescent sexual and reproductive health in sub-Saharan Africa: Who is left behind? BMJ Glob. Health 2020, 5, e002231. [Google Scholar] [CrossRef]
- Katayama, M.; Wiklander, O.P.B.; Fritz, T.; Caidahl, K.; El-Andaloussi, S.; Zierath, J.R.; Krook, A. Circulating Exosomal miR-20b-5p Is Elevated in Type 2 Diabetes and Could Impair Insulin Action in Human Skeletal Muscle. Diabetes 2019, 68, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Hubal, M.J.; Nadler, E.P.; Ferrante, S.C.; Barberio, M.D.; Suh, J.; Wang, J.; Dohm, G.L.; Pories, W.J.; Mietus-Snyder, M.; Freishtat, R.J. Circulating adipocyte-derived exosomal MicroRNAs associated with decreased insulin resistance after gastric bypass. Obesity 2016, 25, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, S.C.; Nadler, E.P.; Pillai, D.K.; Hubal, M.J.; Wang, Z.; Wang, J.M.; Gordish-Dressman, H.; Koeck, E.; Sevilla, S.; Wiles, A.A.; et al. Adipocyte-derived exosomal miRNAs: A novel mechanism for obesity-related disease. Pediatr. Res. 2014, 77, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.J.; Patel, R.M.; McClintock, T.S.; Dupont-Versteegden, E.E.; Peterson, C.A.; McCarthy, J.J. Myonuclear transcription is responsive to mechanical load and DNA content but uncoupled from cell size during hypertrophy. Mol. Biol. Cell 2016, 27, 788–798. [Google Scholar] [CrossRef]
- Wang, L.; Shang, C.; Pan, H.; Yang, H.; Zhu, H.; Gong, F. MicroRNA Expression Profiles in the Subcutaneous Adipose Tissues of Morbidly Obese Chinese Women. Obes. Facts 2021, 14, 78–92. [Google Scholar] [CrossRef]
- Liu, G.; Lei, Y.; Luo, S.; Huang, Z.; Chen, C.; Wang, K.; Yang, P.; Huang, X. MicroRNA expression profile and identification of novel microRNA biomarkers for metabolic syndrome. Bioengineered 2021, 12, 3864–3872. [Google Scholar] [CrossRef]
- Samandari, N.; Mirza, A.H.; Kaur, S.; Hougaard, P.; Nielsen, L.B.; Fredheim, S.; Mortensen, H.B.; Pociot, F. Influence of Disease Duration on Circulating Levels of miRNAs in Children and Adolescents with New Onset Type 1 Diabetes. Non-Coding RNA 2018, 4, 35. [Google Scholar] [CrossRef]
- Masaki, T. Effect of the anti-diabetic drug metformin in hepatocellular carcinoma in vitro and in vivo. Int. J. Oncol. 2013, 45, 322–332. [Google Scholar] [CrossRef]
- Jin, Z.-Q. MicroRNA targets and biomarker validation for diabetes-associated cardiac fibrosis. Pharmacol. Res. 2021, 174, 105941. [Google Scholar] [CrossRef]
- Hou, J.; Liang, W.Y.; Xiong, S.; Long, P.; Yue, T.; Wen, X.; Wang, T.; Deng, H. Identification of hub genes and potential ceRNA networks of diabetic cardiomyopathy. Sci. Rep. 2023, 13, 10258. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, W.; Zhang, W.; Wang, W.; Liu, T.; Zhou, X. LncRNA DCRF regulates cardiomyocyte autophagy by targeting miR-551b-5p in diabetic cardiomyopathy. Theranostics 2019, 9, 4558–4566. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Zhang, K.; Wang, Y.-L.; Zhang, F.; Cao, J.; Zheng, J.-B.; Zhang, H.-J. MiR-551b-5p Contributes to Pathogenesis of Vein Graft Failure via Upregulating Early Growth Response-1 Expression. Chin. Med. J. 2017, 130, 1578–1585. [Google Scholar] [CrossRef]
- Khan, R.; Kadamkode, V.; Kesharwani, D.; Purkayastha, S.; Banerjee, G.; Datta, M. Circulatory miR-98-5p levels are deregulated during diabetes and it inhibits proliferation and promotes apoptosis by targeting PPP1R15B in keratinocytes. RNA Biol. 2019, 17, 188–201. [Google Scholar] [CrossRef]
- Guo, Z.; Cao, Y. An lncRNA-miRNA-mRNA ceRNA network for adipocyte differentiation from human adipose-derived stem cells. Mol. Med. Rep. 2019, 19, 4271–4287. [Google Scholar] [CrossRef] [PubMed]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.T.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA Profiles in Patients with Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271–E2276. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wei, Z.; Wu, X.; Yang, H. Screening of exosomal miRNAs derived from subcutaneous and visceral adipose tissues: Determination of targets for the treatment of obesity and associated metabolic disorders. Mol. Med. Rep. 2018, 18, 3314–3324. [Google Scholar] [CrossRef]
- Petersmann, A.; Nauck, M.; Müller-Wieland, D.; Kerner, W.; Müller, U.A.; Landgraf, R.; Freckmann, G.; Heinemann, L. Definition, Classification and Diagnosis of Diabetes Mellitus. Exp. Clin. Endocrinol. Diabetes 2018, 126, 406–410. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Sebestyén, E.; Nagy, A.; Marosvári, D.; Rajnai, H.; Kajtár, B.; Deák, B.; Matolcsy, A.; Brandner, S.; Storhoff, J.; Chen, N.; et al. Distinct miRNA Expression Signatures of Primary and Secondary Central Nervous System Lymphomas. J. Mol. Diagn. 2021, 24, 224–240. [Google Scholar] [CrossRef]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.-P.; Meese, E.; et al. miRTargetLink 2.0—Interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nature Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Guo, Q.; Nie, H.; Zhou, C.; Cai, Q.; Huang, J.; Meng, X. Linear-hairpin variable primer RT-qPCR for MicroRNA. Chem. Sci. 2018, 10, 2034–2043. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups/ Parameter | Obesity without T2DM n = 9 | Obesity with T2DM n = 15 | Control Group n = 7 | p |
|---|---|---|---|---|
| Age, years | 41.0 ± 14.7 | 47.9 ± 10.9 | 45.3 ± 4.9 | 10.215 20.781 30.527 |
| Sex (male/female) | 2/7 | 6/9 | 2/5 | |
| Weight (kg) | 118.6 ± 18.1 | 140.8 ± 21.4 | 66.0 ± 8.5 | 10.046 20.021 30.008 |
| Body mass index, kg/m2 | 41.9 ± 5.6 | 48.9 ± 6.1 | 24.9 ± 2.8 | 10.016 20.017 30.006 |
| Waist circumference, cm | 107.0 (99.0–143.0) | 145.0 (123.0–160.0) | nd | 10.002 |
| Hip, cm | 126.5 (100.0–141.0) | 135 (115.0–145.0) | nd | 10.159 |
| Waist-to-hip ratio | 0.9 ± 0.2 | 1.1 ± 0.1 | nd | 10.027 |
| Glucose, nmol/L | 5.4 (4.3–8.1) | 7.3 (5.5–12.7) | 4.9 (4.3–5.0) | 10.007 20.389 30.007 |
| Insulin, µIU/mL | 26.86 (9.98–41.4) | 28.1 (16.3–79.4) | nd | 10.907 |
| HOMA-IR index | 6.1 ± 2.7 | 10.7 ± 5.4 | nd | 10.165 |
| C-peptide (ng/mL) | 3.5 (1.8–4.6) | 3.8 (1.9–11.9) | nd | 10.587 |
| HbA1c, % | 5.6 ± 0.2 | 7.45 ± 1.7 | nd | 10.010 |
| Total cholesterol, mmol/L | 4.9 ± 1.4 | 5.1 ± 0.8 | nd | 11.000 |
| HDL cholesterol, mmol/L | 1.6 ± 0.3 | 1.2 ± 0.2 | nd | 10.182 |
| LDL cholesterol, mmol/L | 2.9 ± 1.4 | 2.7 ± 0.9 | nd | 10.570 |
| Triglyceride, mmol/L | 1.7 (1.2–2.9) | 2.2 (1.3–4.4) | nd | 10.101 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dracheva, K.V.; Pobozheva, I.A.; Anisimova, K.A.; Balandov, S.G.; Grunina, M.N.; Hamid, Z.M.; Vasilevsky, D.I.; Pchelina, S.N.; Miroshnikova, V.V. Downregulation of Exosomal hsa-miR-551b-3p in Obesity and Its Link to Type 2 Diabetes Mellitus. Non-Coding RNA 2023, 9, 67. https://doi.org/10.3390/ncrna9060067
Dracheva KV, Pobozheva IA, Anisimova KA, Balandov SG, Grunina MN, Hamid ZM, Vasilevsky DI, Pchelina SN, Miroshnikova VV. Downregulation of Exosomal hsa-miR-551b-3p in Obesity and Its Link to Type 2 Diabetes Mellitus. Non-Coding RNA. 2023; 9(6):67. https://doi.org/10.3390/ncrna9060067
Chicago/Turabian StyleDracheva, Kseniia V., Irina A. Pobozheva, Kristina A. Anisimova, Stanislav G. Balandov, Maria N. Grunina, Zarina M. Hamid, Dmitriy I. Vasilevsky, Sofya N. Pchelina, and Valentina V. Miroshnikova. 2023. "Downregulation of Exosomal hsa-miR-551b-3p in Obesity and Its Link to Type 2 Diabetes Mellitus" Non-Coding RNA 9, no. 6: 67. https://doi.org/10.3390/ncrna9060067