
Inverse Impact of Cancer Drugs on Circular and Linear RNAs in Breast Cancer Cell Lines
 , , ,
, , ,  and
and
Abstract
:
1. Introduction
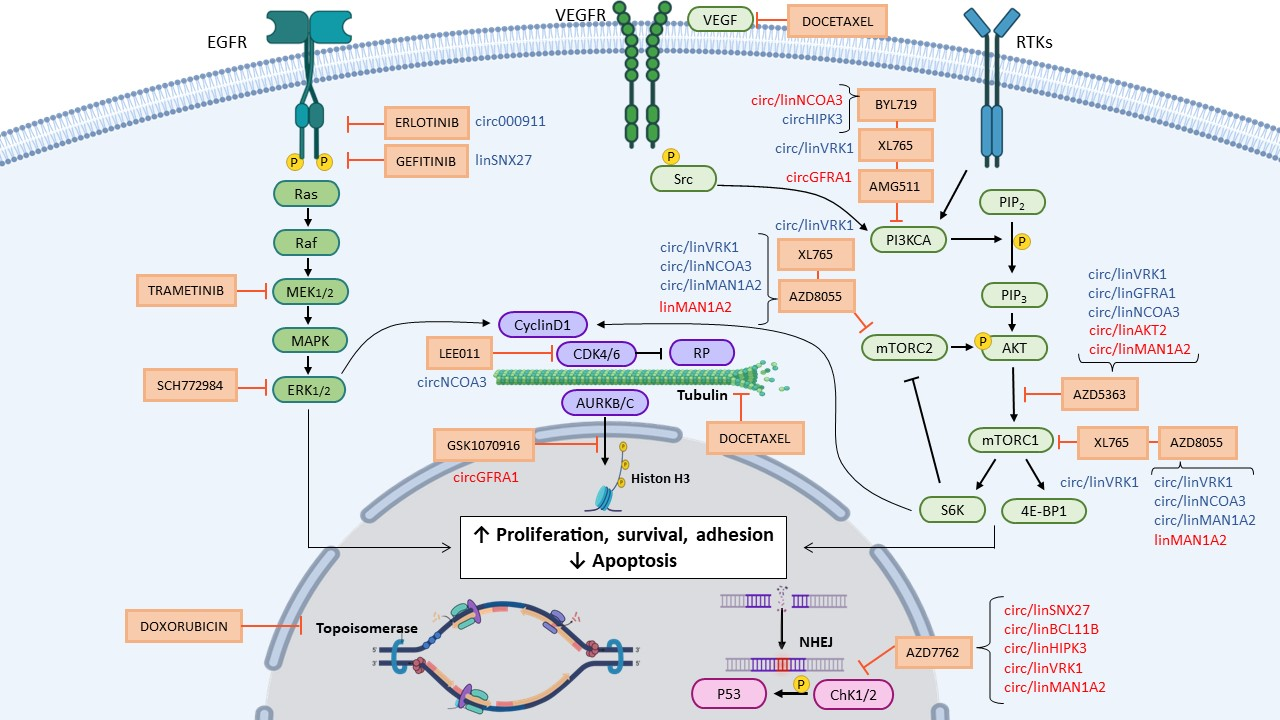
2. Results
2.1. Drugs Differentially Regulating the Expression of circRNAs and Cognate Linear Transcripts
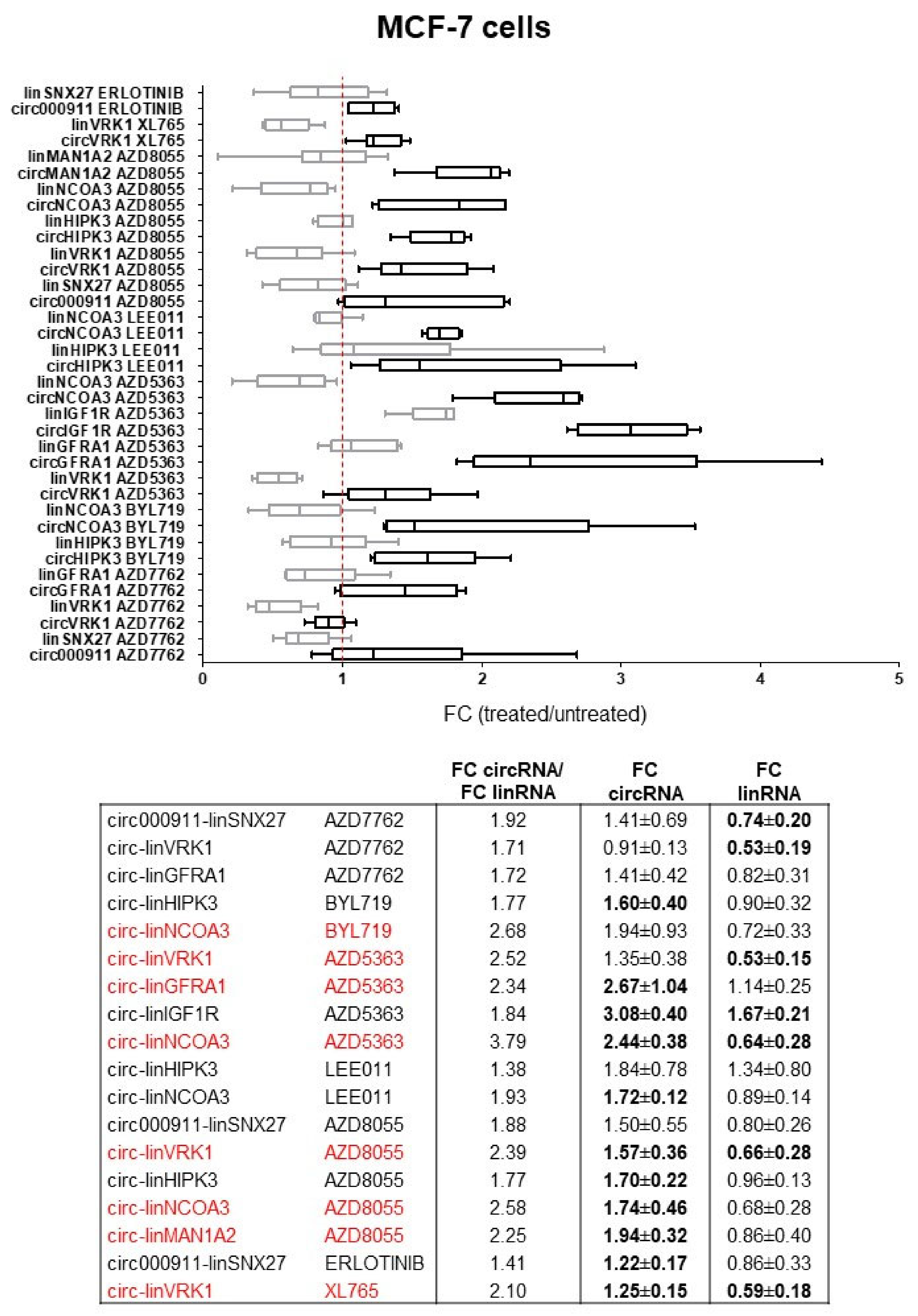
2.1.1. circRNAs and Cognate linRNAs Modulated by Drugs in MCF-7 Cells
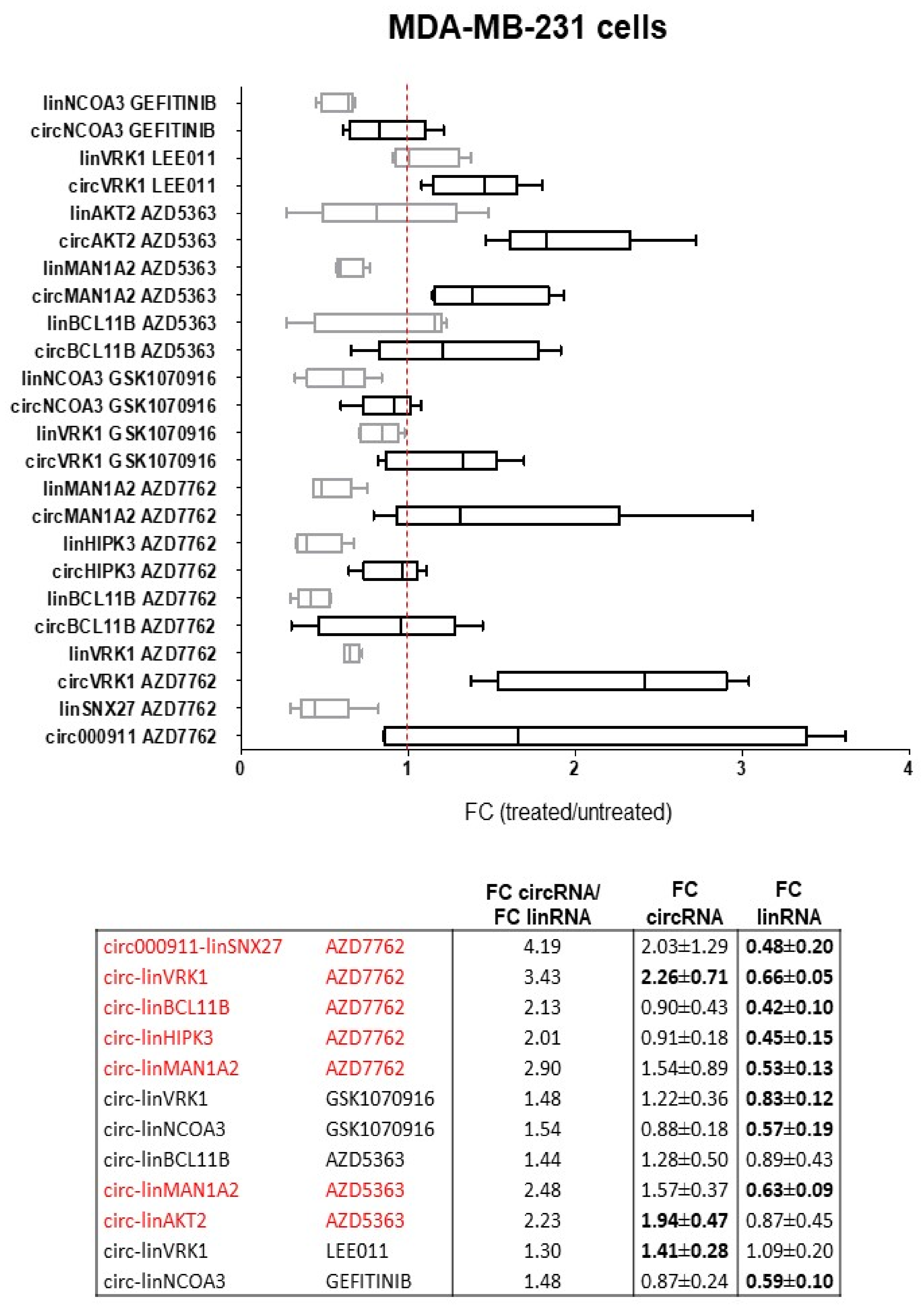
2.1.2. circRNAs and Cognate linRNAs Modulated by Drugs in MDA-MB-231 Cells
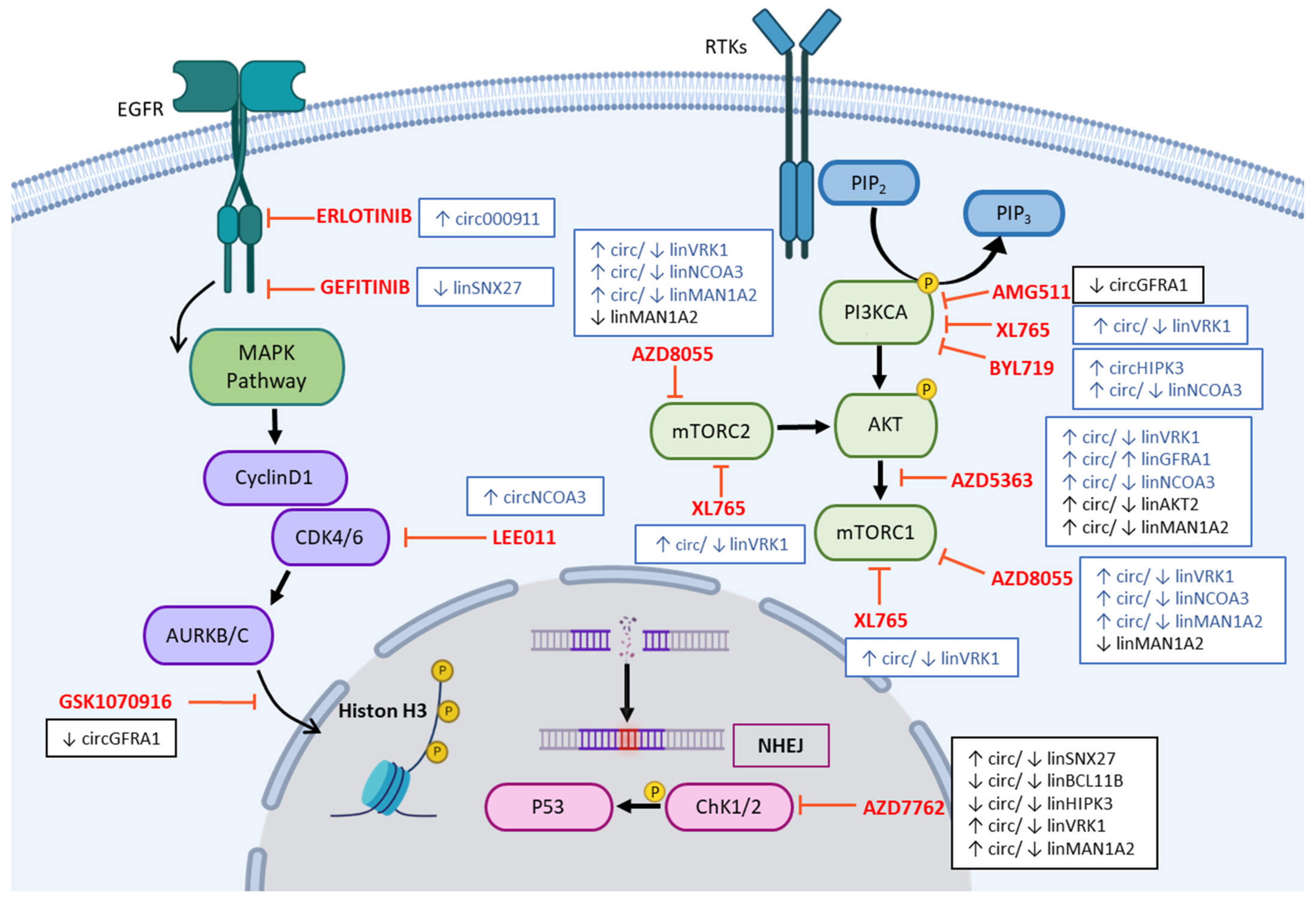
2.2. Dysregulation of Specific circRNAs versus Their Cognate linRNAs in BC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Anticancer Drugs Treatments
4.2. RT-qPCR Analysis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, X.; Wang, J.; Guo, Z.; Li, M.; Li, M.; Liu, S.; Liu, H.; Li, W.; Yin, X.; Tao, J.; et al. Emerging function and potential diagnostic value of circular RNAs in cancer. Mol. Cancer 2018, 17, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-Y.; Chen, W.; Yang, S.-J.; Xu, Z.-H.; Hu, J.-H.; Zhang, H.-D.; Zhong, S.-L.; Tang, J.-H. The emerging role of circular RNAs in breast cancer. Biosci. Rep. 2019, 39, BSR20190621. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, Y.; Liang, G.; Ling, Y.; Tan, W.; Tan, L.; Andrews, R.; Zhong, W.; Zhang, X.; Song, E.; et al. Circular RNA hsa_circ_001783 regulates breast cancer progression via sponging miR-200c-3p. Cell Death Dis. 2019, 10, 55–69. [Google Scholar] [CrossRef]
- Zhang, H.; Ge, Z.; Wang, Z.; Gao, Y.; Wang, Y.; Qu, X. Circular RNA RHOT1 promotes progression and inhibits ferroptosis via mir-106a-5p/STAT3 axis in breast cancer. Aging 2021, 13, 8115–8126. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, S.; Liao, X.; Li, M.; Chen, S.; Li, X.; Wu, X.; Yang, M.; Tang, M.; Hu, Y.; et al. Circular RNA circIKBKB promotes breast cancer bone metastasis through sustaining NF-κB/bone remodeling factors signaling. Mol. Cancer 2021, 20, 106–124. [Google Scholar] [CrossRef]
- Fu, B.; Liu, W.; Zhu, C.; Li, P.; Wang, L.; Pan, L.; Li, K.; Cai, P.; Meng, M.; Wang, Y.; et al. Circular RNA circBCBM1 promotes breast cancer brain metastasis by modulating miR-125a/BRD4 axis. Int. J. Biol. Sci. 2021, 17, 3104–3117. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Hussen, B.M.; Taheri, M.; Ayatollahi, S.A. Emerging role of circular RNAs in breast cancer. Pathol. Res. Pract. 2021, 223, 153496–153510. [Google Scholar] [CrossRef]
- Li, J.; Ma, M.; Yang, X.; Zhang, M.; Luo, J.; Zhou, H.; Huang, N.; Xiao, F.; Lai, B.; Lv, W.; et al. Circular HER2 RNA positive triple negative breast cancer is sensitive to Pertuzumab. Mol. Cancer 2020, 19, 142–160. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Li, Q.; Li, X.; Feng, X. A novel circular RNA confers trastuzumab resistance in human epidermal growth factor receptor 2-positive breast cancer through regulating ferroptosis. Environ. Toxicol. 2022, 37, 1597–1607. [Google Scholar] [CrossRef]
- López-Knowles, E.; O’Toole, S.A.; McNeil, C.M.; Millar, E.K.; Qiu, M.R.; Crea, P.; Daly, R.J.; Musgrove, E.A.; Sutherland, R.L. PI3K pathway activation in breast cancer is associated with the basal-like phenotype and cancer-specific mortality. Int. J. Cancer 2010, 126, 1121–1131. [Google Scholar] [CrossRef]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/mTOR pathway causes drug resistance in breast cancer. Front. Pharmacol. 2021, 12, 628690–628706. [Google Scholar] [CrossRef] [PubMed]
- Baldassari, F.; Zerbinati, C.; Galasso, M.; Corrà, F.; Minotti, L.; Agnoletto, C.; Previati, M.; Croce, C.M.; Volinia, S. Screen for MicroRNA and drug interactions in breast cancer cell lines points to miR-126 as a modulator of CDK4/6 and PIK3CA inhibitors. Front. Genet. 2018, 9, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.A.; Niu, N.; Tang, X.; Thompson, K.J.; Wang, L.; Kocher, J.-P.; Subramanian, S.; Kalari, K.R. Circular RNAs and their associations with breast cancer subtypes. Oncotarget 2016, 7, 80967–80979. [Google Scholar] [CrossRef]
- Tarrero, L.C.; Ferrero, G.; Miano, V.; De Intinis, C.; Ricci, L.; Arigoni, M.; Riccardo, F.; Annaratone, L.; Castellano, I.; Calogero, R.A.; et al. Luminal breast cancer-specific circular RNAs uncovered by a novel tool for data analysis. Oncotarget 2018, 9, 14580–14596. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Zhou, L.; Zhang, L.; Yin, K.; Peng, J.; Sha, R.; Zhang, S.; Xu, Y.; Sheng, X.; Wang, Y.; et al. Identification and integrated analysis of key differentially expressed circular RNAs in ER-positive subtype breast cancer. Epigenomics 2019, 11, 297–321. [Google Scholar] [CrossRef]
- Tian, T.; Zhao, Y.; Zheng, J.; Jin, S.; Liu, Z.; Wang, T. Circular RNA: A potential diagnostic, prognostic, and therapeutic biomarker for human triple-negative breast cancer. Mol. Ther. Nucleic Acids 2021, 26, 63–80. [Google Scholar] [CrossRef]
- Yan, N.; Xu, H.; Zhang, J.; Xu, L.; Zhang, Y.; Zhang, L.; Xu, Y.; Zhang, F. Circular RNA profile indicates circular RNA VRK1 is negatively related with breast cancer stem cells. Oncotarget 2017, 8, 95704–95718. [Google Scholar] [CrossRef]
- Wang, H.; Xiao, Y.; Wu, L.; Ma, D. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-000911/miR-449a pathway in breast carcinogenesis. Int. J. Oncol. 2018, 52, 743–754. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.-M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The landscape of circular RNA in cancer. Cell 2019, 176, 869–881.e13. [Google Scholar] [CrossRef]
- Li, Y.; Li, H. Circular RNA VRK1 correlates with favourable prognosis, inhibits cell proliferation but promotes apoptosis in breast cancer. J. Clin. Lab. Anal. 2019, 34, e22980–e22988. [Google Scholar] [CrossRef]
- He, R.; Liu, P.; Xie, X.; Zhou, Y.; Liao, Q.; Xiong, W.; Li, X.; Li, G.; Zeng, Z.; Tang, H. circGFRA1 and GFRA1 act as ceRNAs in triple negative breast cancer by regulating miR-34a. J. Exp. Clin. Cancer Res. 2017, 36, 137–145. [Google Scholar] [CrossRef] [PubMed]
- De Palma, F.D.E.; Salvatore, F.; Pol, J.G.; Kroemer, G.; Maiuri, M.C. Circular RNAs as potential biomarkers in breast cancer. Biomedicines 2022, 10, 725. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Niu, X.; Yan, S.; Liu, Y.; Dong, R.; Li, Y. Circular RNA profiling facilitates the diagnosis and prognostic monitoring of breast cancer: A pair-wise meta-analysis. J. Clin. Lab. Anal. 2020, 35, e23575–e23589. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, C.; Wang, Y. Exosome-mediated transfer of circHIPK3 promotes trastuzumab chemoresistance in breast cancer. J. Drug Target. 2021, 29, 1004–1015. [Google Scholar] [CrossRef]
- Ni, J.; Xi, X.; Xiao, S.; Xiao, X. Silencing of circHIPK3 sensitizes paclitaxel-resistant breast cancer cells to chemotherapy by regulating HK2 through targeting miR-1286. Cancer Manag. Res. 2021, 13, 5573–5585. [Google Scholar] [CrossRef]
- Sharma, P.; Parveen, S.; Shah, L.V.; Mukherjee, M.; Kalaidzidis, Y.; Kozielski, A.J.; Rosato, R.; Chang, J.C.; Datta, S. SNX27–retromer assembly recycles MT1-MMP to invadopodia and promotes breast cancer metastasis. J. Cell Biol. 2020, 219, e201812098. [Google Scholar] [CrossRef]
- Zhang, J.; Li, K.; Zhang, Y.; Lu, R.; Wu, S.; Tang, J.; Xia, Y.; Sun, J. Deletion of sorting nexin 27 suppresses proliferation in highly aggressive breast cancer MDA-MB-231 cells in vitro and in vivo. BMC Cancer 2019, 19, 555. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, J.; Tang, Y.; Zhao, J.; He, S.; Xiong, F.; Guo, C.; Xiang, B.; Zhou, M.; Li, X.; et al. circ MAN 1A2 could serve as a novel serum biomarker for malignant tumors. Cancer Sci. 2019, 110, 2180–2188. [Google Scholar] [CrossRef]
- Liu, L.; Sang, M.; Shi, J.; Zheng, Y.; Meng, L.; Gu, L.; Li, Z.; Liu, F.; Bu, J.; Duan, X.; et al. CircRNA mannosidase alpha class 1A member 2 promotes esophageal squamous cell carcinoma progression by regulating C–C chemokine ligand 5. Biochem. Biophys. Res. Commun. 2023, 645, 61–70. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, X.; Wang, Y.; Chi, N.; Yu, J.; Fu, X. circRNA mannosidase alpha class 1A member 2 contributes to the proliferation and motility of papillary thyroid cancer cells through upregulating metadherin via absorbing microRNA-449a. Anti-Cancer Drugs 2023, 34, 44–56. [Google Scholar] [CrossRef]
- Mon, A.M.; MacKinnon, A.C., Jr.; Traktman, P. Overexpression of the VRK1 kinase, which is associated with breast cancer, induces a mesenchymal to epithelial transition in mammary epithelial cells. PLoS ONE 2018, 13, e0203397. [Google Scholar] [CrossRef] [PubMed]
- Burwinkel, B.; Wirtenberger, M.; Klaes, R.; Schmutzler, R.K.; Grzybowska, E.; Försti, A.; Frank, B.; Bermejo, J.L.; Bugert, P.; Wappenschmidt, B.; et al. Association of NCOA3 polymorphisms with breast cancer risk. Clin. Cancer Res. 2005, 11, 2169–2174. [Google Scholar] [CrossRef]
- Yao, J.; Xu, G.; Zhu, L.; Zheng, H. circGFRA1 enhances NSCLC progression by sponging miR-188-3p. OncoTargets Ther. 2020, 13, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Zhang, J.; Yan, L.; Li, D. CircHIPK3/miR-381-3p axis modulates proliferation, migration, and glycolysis of lung cancer cells by regulating the AKT/mTOR signaling pathway. Open Life Sci. 2020, 15, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Guo, F.; Zhu, H.; Tan, X.; Zhu, X.; Liu, X.; Zhang, W.; Yang, Q.; Jiang, Y. Circular RNA-0001283 suppresses breast cancer proliferation and invasion via MiR-187/HIPK3 axis. Experiment 2020, 26, e921502. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Liu, S.; Li, X.; Hu, Y.; Zhang, K. Circular RNA circHIPK3 promotes breast cancer progression via sponging MiR-326. Cell Cycle 2021, 20, 1320–1333. [Google Scholar] [CrossRef]
- Yin, H.; Cui, X. Knockdown of circHIPK3 facilitates temozolomide sensitivity in glioma by regulating cellular behaviors through miR-524-5p/KIF2A-Mediated PI3K/AKT Pathway. Cancer Biother. Radiopharm. 2021, 36, 556–567. [Google Scholar] [CrossRef]
- Chen, X.; Mao, R.; Su, W.; Yang, X.; Geng, Q.; Guo, C.; Wang, Z.; Wang, J.; Kresty, L.A.; Beer, D.G.; et al. Circular RNA circHIPK3 modulates autophagy via MIR124-3p-STAT3-PRKAA/AMPKα signaling in STK11 mutant lung cancer. Autophagy 2020, 16, 659–671. [Google Scholar] [CrossRef]
- Wei, Z.; Shi, Y.; Xue, C.; Li, M.; Wei, J.; Li, G.; Xiong, W.; Zhou, M. Understanding the dual roles of CircHIPK3 in tumorigenesis and tumor progression. J. Cancer 2022, 13, 3674–3686. [Google Scholar] [CrossRef]
- Qi, L.; Sun, B.; Yang, B.; Lu, S. circHIPK3 (hsa_circ_0000284) Promotes proliferation, migration and invasion of breast cancer cells via miR-326. OncoTargets Ther. 2021, 14, 3671–3685. [Google Scholar] [CrossRef]
- Pereira, L.; Horta, S.; Mateus, R.; Videira, M.A. Implications of Akt2/Twist crosstalk on breast cancer metastatic outcome. Drug Discov. Today 2015, 20, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Li, X.; Li, F.; Wu, X.; Zhang, M.; Zhou, H.; Huang, N.; Yang, X.; Xiao, F.; Liu, D.; et al. A novel tumor suppressor protein encoded by circular AKT3 RNA inhibits glioblastoma tumorigenicity by competing with active phosphoinositide-dependent Kinase-1. Mol. Cancer 2019, 18, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.R.; Greenwood, H.; Dudley, P.; Crafter, C.; Yu, D.-H.; Zhang, J.; Li, J.; Gao, B.; Ji, Q.; Maynard, J.; et al. Preclinical pharmacology of AZD5363, an inhibitor of AKT: Pharmacodynamics, antitumor activity, and correlation of monotherapy activity with genetic background. Mol. Cancer Ther. 2012, 11, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Borges, S.; Lazo, P.A. The human vaccinia-related kinase 1 (VRK1) phosphorylates threonine-18 within the mdm-2 binding site of the p53 tumour suppressor protein. Oncogene 2000, 19, 3656–3664. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, A.; Santos, C.R.; Barcia, R.; Vega, F.M.; Lazo, P. c-Jun phosphorylation by the human vaccinia-related kinase 1 (VRK1) and its cooperation with the N-terminal kinase of c-Jun (JNK). Oncogene 2004, 23, 8950–8958. [Google Scholar] [CrossRef]
- Nichols, R.J.; Wiebe, M.; Traktman, P. The vaccinia-related kinases phosphorylate the N′ terminus of BAF, regulating its interaction with DNA and its retention in the nucleus. Mol. Biol. Cell 2006, 17, 2451–2464. [Google Scholar] [CrossRef]
- Sevilla, A.; Santos, C.R.; Vega, F.M.; Lazo, P.A. Human vaccinia-related Kinase 1 (VRK1) activates the ATF2 transcriptional activity by novel phosphorylation on Thr-73 and Ser-62 and cooperates with JNK. J. Biol. Chem. 2004, 279, 27458–27465. [Google Scholar] [CrossRef]
- Campillo-Marcos, I.; García-González, R.; Navarro-Carrasco, E.; Lazo, P.A. The human VRK1 chromatin kinase in cancer biology. Cancer Lett. 2021, 503, 117–128. [Google Scholar] [CrossRef]
- García-González, R.; Morejón-García, P.; Campillo-Marcos, I.; Salzano, M.; Lazo, P.A. VRK1 phosphorylates Tip60/KAT5 and is required for H4K16 acetylation in response to DNA damage. Cancers 2020, 12, 2986. [Google Scholar] [CrossRef]
- Miller, D.H.; Jin, D.X.; Sokol, E.S.; Cabrera, J.R.; Superville, D.A.; Gorelov, R.A.; Kuperwasser, C.; Gupta, P.B. BCL11B drives human mammary stem cell self-renewal in vitro by inhibiting basal differentiation. Stem Cell Rep. 2018, 10, 1131–1145. [Google Scholar] [CrossRef]
- Nalabothula, N.; Al-Jumaily, T.; Eteleeb, A.M.; Flight, R.M.; Xiaorong, S.; Moseley, H.; Rouchka, E.C.; Fondufe-Mittendorf, Y.N. Genome-wide profiling of PARP1 reveals an interplay with gene regulatory regions and DNA methylation. PLoS ONE 2015, 10, e0135410. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.; Lai, S.-T.; Ni, Y.-B.; Shao, Y.; Poon, I.K.; Kwan, J.S.; Chow, C.; Shea, K.-H.; Tse, G.M. SETD2 alterations and histone H3K36 trimethylation in phyllodes tumor of breast. Breast Cancer Res. Treat. 2021, 187, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Wang, J.; Zhang, Q.; Xu, H.; Feng, J. CircPrimer: A software for annotating circRNAs and determining the specificity of circRNA primers. BMC Bioinform. 2018, 19, 292. [Google Scholar] [CrossRef] [PubMed]
- Gorji-Bahri, G.; Moradtabrizi, N.; Hashemi, A. Uncovering the stability status of the reputed reference genes in breast and hepatic cancer cell lines. PLoS ONE 2021, 16, e0259669. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| circRNA | Modulation | Functions/ Processes | Reference |
|---|---|---|---|
| circIGF1R | upregulated (MCF-7 vs. MDA-MB-231 cells) | Biogenesis by H3K36me3 | Coscujuela Tarrero et al., 2018 [14] |
| circESR1 | upregulated (MCF-7 vs. MDA-MB-231 cells) | Biogenesis by H3K36me3 | Coscujuela Tarrero et al., 2018 [14] |
| circHIPK3 | similar expression (MCF-7 vs. MDA-MB-231 cells) | Biogenesis by H3K36me3 | Coscujuela Tarrero et al., 2018 [14] |
| circNCOA3 | upregulated (MCF-7 vs. MDA-MB-231 cells) | Biogenesis by H3K36me3 | Coscujuela Tarrero et al., 2018 [14] |
| circMAN1A2 | upregulated (MCF-7 vs. MDA-MB-231 cells) | Biogenesis by H3K36me3 | Coscujuela Tarrero et al., 2018 [14] |
| circVRK1 | downregulated | Inversely correlated with stemness of CSC 1, cell proliferation | Yan et al., 2017 [17] Li et al., 2020 [20] |
| circBCL11B | downregulated | Stemness of CSC 1 | Yan et al., 2017 [17] |
| circ000911 | downregulated | Cell proliferation, migration and invasion, acting as a sponge for miR-449a, targeting NF-κB signaling | Wang et al., 2018 [18] |
| circGFRA1 | upregulated in triple-negative BC | Induction of cell proliferation, acting as ceRNAs 2 through the regulation of miR-34a, poor survival of patients | He et al., 2017 [21] |
| circAKT2 | frequently expressed in tumors | Akt/PI3K signaling | Vo et al., 2019 [19] |
| circPIK3CB | frequently expressed in tumors | Akt/PI3K signaling | Vo et al., 2019 [19] |
| circAKT3 | frequently expressed in tumors | Akt/PI3K signaling | Vo et al., 2019 [19] |
| Drug | Commercial Name/Company | Mechanism of Action/ Affected Pathway | Application |
|---|---|---|---|
| AZD5363 | CAPIVASERTIB (AstraZeneca) | It binds all AKT isoforms inhibiting the substrates’ phosphorylation | Phase 3 study in combination with paclitaxel to treat advanced or metastatic triple negative BCs |
| AZD7762 | ATP-competitive CHK1/2 inhibitor, affecting non-homologous end-joining (NHEJ) pathway that repairs double-strand breaks in DNA | Phase 1 study, also with gemcitabine in advanced solid tumors | |
| AZD8055 | ATP-competitive mTORC1/C2 inhibitor | Phase 1 study, patients with gliomas that have not responded to conventional treatments | |
| BYL719 | ALPELISIB (Novartis) | PI3K α-isoform (PIK3CA) specific inhibitor | Authorized by European Medicines Agency (EMA) in combination with Fulvestrant to improve survival in patients with advanced HR+/HER2− BCs with PIK3CA gene mutation |
| ERLOTINIB | TARCEVA | It inhibits the activity of mutated and wildtype Epidermal Growth Factor Receptor (EGFR) tyrosine kinases | Approved by EMA to cure locally advanced or metastatic Non-Small Cell Lung cancer and in patients with metastatic pancreatic cancer, in combination with Gemcitabine |
| GEFITINIB | IRESSA | It inhibits the activity of mutated EGFR tyrosine kinases | Approved by EMA for the exclusive use in NSCL cancer with EGFR mutations |
| GSK1070916 | ATP competitive inhibitor of Aurora B/C kinases, inhibiting the Histone H3 phosphorylation | Phase 1 study for the treatment of adult solid tumors | |
| GSK1120212 (TRAMETINIB) | MEKINIST | Inhibitor blocking MEK 1/2 activated by mutated BRAF protein, preventing cell proliferation | Approved for medical use by EMA to treat melanoma and non-small cell lung cancer with BRAFV600 mutation |
| LEE011 | RIBOCICLIB (KISQALI) | Inhibitor of Cyclin-dependent kinase 4 and 6 (CDK 4/6), activated upon binding to D cyclins | Phase 3 study in Combination with Letrozole HR+/HER2− advanced BCs with no prior hormonal therapy |
| SCH772984 | orally bioavailable as MK-8353 (Merck) | ATP competitive inhibitor of selective ERK1/2 suppressing MAPK pathway signaling through induction of an inactive conformation of the phosphate-binding loop and a shift of the α-C helix. It caused G1 arrest and induced apoptosis | Phase I clinical trial in patients with advanced solid tumors, in particular melanoma and colorectal cancer with BRAFV600 or NRASQ61 somatic mutations. |
| DOXORUBICIN | Adriamycin®, as well as Rubex® | DNA intercalant, TOPOiso II inhibitor | It was used to treat different types of cancers, including BCs. |
| DOCETAXEL | TAXOTERE | It binds tubulin, promoting the assembly and blocking the disassembly of microtubules. It causes cell-cycle arrest at the G2/M phase and inhibits Vascular endothelial growth factor (VEGF) | Phase III trial of DOCETAXEL plus cisplatin in patients with stage IV NSCL cancer, and of BC, prostate, and stomach cancer |
| AMG511 | orally bioavailable as AOBIOUS | Selective inhibitor of pan-class I phosphatidylinositol-3 kinases (PI3Kα, β, δ and γ), selective over mTOR hVPS34, DNAPK, and a broad panel of other protein kinases. It effectively inhibited pAKT | It is currently in clinical trials for cancer treatment. In animal studies it efficaciously inhibited tumor growth in PTEN-null, KRAS mutant, and HER2 amplified xenograft. |
| XL765 | VOXATALISIB | Reversible ATP-competitive inhibitor of pan-Class I PI3K (α, β, γ, and δ) and mTORC1/mTORC2 | In clinical trials for the treatment of Glioblastoma in combination therapy with Temozolomide, prostate cancer, non-Hodgkin lymphoma or chronic lymphocytic leukaemia |
| MCF-7 Cells | MDA-MB-231 Cells | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| circRNAs | linRNAs | circRNAs | linRNAs | |||||||
| DRUGS | Drug Sensitivity (IC50 μM) | FC | Adj p-Value | FC | Adj p-Value | Drug Sensitivity (IC50 μM) | FC | Adj p-Value | FC | Adj p-Value |
| AZD5363 | 41.4 | 0.69 | 0.008 | 2.39 | - | 111.7 | 0.05 | 0.026 | 0.22 | - |
| AZD7762 | 6.8 | 0.54 | - | 2.01 | 0.039 | 1.5 | 0.06 | - | 0.14 | 1.0 × 10−7 |
| AZD8055 | 0.5 | 0.65 | 0.027 | 2.78 | - | 0.5 | 0.04 | - | 0.25 | - |
| BYL719 | 9.5 | 0.67 | - | 2.74 | - | 81.9 | 0.04 | - | 0.27 | - |
| ERLOTINIB | 95.6 | 0.60 | - | 3.05 | - | 36.2 | 0.05 | - | 0.24 | 0.042 |
| GEFITINIB | 99.6 | 0.34 | - | 2.01 | 0.039 | 46.7 | 0.04 | - | 0.22 | 4.2 × 10−4 |
| GSK1070916 | - | 0.36 | - | 2.34 | - | - | 0.03 | - | 0.21 | 2.3 × 10−5 |
| TRAMETINIB | 48.2 | 0.45 | - | 2.81 | - | 0.3 | 0.05 | - | 0.26 | - |
| LEE011 | 30.1 | 0.66 | - | 3.35 | - | 38.4 | 0.05 | - | 0.27 | - |
| SCH772984 | 203.2 | 0.38 | - | 2.71 | - | 2.6 | 0.05 | - | 0.31 | - |
| DOXORUBICIN | 0.1 | 0.71 | - | 3.97 | 0.001 | 0.1 | 0.07 | - | 0.46 | 5.0 × 10−6 |
| DOCETAXEL | 0.01 | 0.25 | 0.039 | 2.50 | - | 0.01 | 0.04 | - | 0.26 | - |
| AMG511 | - | 0.35 | - | 2.79 | - | - | 0.04 | - | 0.26 | - |
| XL765 | 91.4 | 0.54 | 0.015 | 3.41 | 0.008 | 75.2 | 0.04 | - | 0.26 | - |
| Drug | circRNA | FC (2−ΔΔCT) | p-Value | Adj p-Value | linRNA | FC (2−ΔΔCT) | p-Value | Adj p-Value |
|---|---|---|---|---|---|---|---|---|
| AZD5363 | circESR1 | 1.93 ± 0.72 | 0.046 | - | - | - | - | - |
| AZD5363 | circGFRA1 | 2.67 ± 1.04 | 0.02 | - | - | - | - | - |
| AZD5363 | circHIPK3 | 2.02 ± 0.79 | 0.045 | - | - | - | - | - |
| AZD5363 | circIGF1R | 3.08 ± 0.40 | 0.002 | - | linIGF1R | 1.67 ± 0.21 | 0.002 | - |
| AZD5363 | circMAN1A2 | 2.08 ± 0.58 | 0.01 | - | - | - | - | - |
| AZD5363 | circNCOA3 | 2.44 ± 0.38 | 0.001 | - | linNCOA3 | 0.64 ± 0.28 | 0.047 | - |
| AZD5363 | - | - | - | - | linVRK1 | 0.53 ± 0.15 | 0.002 | - |
| AZD7762 | - | - | - | - | linBCL11B | 0.43 ± 0.26 | 0.003 | - |
| AZD7762 | - | - | - | - | linESR1 | 0.63 ± 0.16 | 0.007 | - |
| AZD7762 | - | - | - | - | linSNX27 | 0.74 ± 0.20 | 0.02 | - |
| AZD7762 | - | - | - | - | linVRK1 | 0.53 ± 0.19 | 0.01 | - |
| AZD8055 | - | - | - | - | linESR1 | 1.93 ± 0.39 | 0.006 | - |
| AZD8055 | - | - | - | - | linGFRA1 | 1.30 ± 0.19 | 0.03 | - |
| AZD8055 | - | - | - | - | linIGF1R | 1.59 ± 0.38 | 0.03 | - |
| AZD8055 | circHIPK3 | 1.70 ± 0.22 | 0.002 | - | - | - | - | - |
| AZD8055 | circMAN1A2 | 1.94 ± 0.32 | 0.003 | - | - | - | - | - |
| AZD8055 | circNCOA3 | 1.74 ± 0.46 | 0.02 | - | - | - | - | - |
| AZD8055 | circVRK1 | 1.57 ± 0.36 | 0.006 | - | linVRK1 | 0.66 ± 0.28 | 0.03 | - |
| BYL719 | - | - | - | - | linESR1 | 1.57 ± 0.22 | 0.004 | - |
| BYL719 | circHIPK3 | 1.60 ± 0.40 | 0.03 | - | - | - | - | - |
| BYL719 | - | - | - | - | linVRK1 | 0.74 ± 0.18 | 0.02 | - |
| DOCETAXEL | circESR1 | 0.79 ± 0.15 | 0.03 | - | - | - | - | - |
| DOCETAXEL | circHIPK3 | 0.65 ± 0.22 | 0.03 | - | - | - | - | - |
| DOCETAXEL | circIGF1R | 0.72 ± 0.17 | 0.02 | - | - | - | - | - |
| DOCETAXEL | circMAN1A2 | 0.66 ± 0.13 | 0.004 | - | - | - | - | - |
| DOCETAXEL | circNCOA3 | 0.71 ± 0.14 | 0.009 | - | - | - | - | - |
| DOXORUBICIN | - | - | - | - | linESR1 | 1.71 ± 0.40 | 0.02 | - |
| DOXORUBICIN | circGFRA1 | 1.86 ± 0.67 | 0.046 | - | linGFRA1 | 1.47 ± 0.30 | 0.02 | - |
| DOXORUBICIN | circHIPK3 | 1.65 ± 0.27 | 0.006 | - | - | - | - | - |
| DOXORUBICIN | - | - | - | - | linIGF1R | 1.89 ± 0.49 | 0.02 | - |
| DOXORUBICIN | circMAN1A2 | 1.73 ± 0.51 | 0.03 | - | - | - | - | - |
| DOXORUBICIN | circNCOA3 | 1.98 ± 0.53 | 0.01 | - | linNCOA3 | 1.67 ± 0.35 | 0.01 | - |
| DOXORUBICIN | - | - | - | - | linSNX27 | 1.60 ± 0.49 | 0.03 | - |
| ERLOTINIB | circ000911 | 1.22 ± 0.17 | 0.045 | - | - | - | - | - |
| GEFITINIB | - | - | - | - | linSNX27 | 0.74 ± 0.25 | 0.0495 | - |
| LEE011 | circNCOA3 | 1.72 ± 0.12 | 0.0002 | 0.04 | - | - | - | - |
| XL765 | circ000911 | 1.61 ± 0.51 | 0.03 | - | - | - | - | - |
| XL765 | circESR1 | 1.55 ± 0.33 | 0.02 | - | linESR1 | 2.07 ± 0.45 | 0.006 | - |
| XL765 | circGFRA1 | 2.01 ± 0.66 | 0.03 | - | linGFRA1 | 1.51 ± 0.15 | 0.002 | - |
| XL765 | circHIPK3 | 1.65 ± 0.43 | 0.03 | - | - | - | - | - |
| XL765 | circIGF1R | 2.04 ± 0.62 | 0.02 | - | linIGF1R | 2.18 ± 0.78 | 0.03 | - |
| XL765 | circMAN1A2 | 1.73 ± 0.32 | 0.007 | - | - | - | - | - |
| XL765 | circNCOA3 | 1.71 ± 0.37 | 0.01 | - | - | - | - | - |
| XL765 | - | - | - | - | linPIK3CB | 1.40 ± 0.23 | 0.02 | - |
| XL765 | circVRK1 | 1.25 ± 0.15 | 0.005 | - | linVRK1 | 0.59 ± 0.18 | 0.01 | - |
| Drug | circRNA | FC (2−ΔΔCT) | p-Value | Adj p-Value | linRNA | FC (2−ΔΔCT) | p-Value | Adj p-Value |
|---|---|---|---|---|---|---|---|---|
| AMG511 | circGFRA1 | 0.71 ± 0.12 | 0.01 | - | - | - | - | - |
| AZD5363 | circAKT2 | 1.94 ± 0.47 | 0.01 | - | - | - | - | - |
| AZD5363 | circAKT3 | 2.37 ± 0.95 | 0.03 | - | linAKT3 | 1.33 ± 0.19 | 0.02 | - |
| AZD5363 | - | - | - | - | linMAN1A2 | 0.63 ± 0.09 | 0.004 | - |
| AZD5363 | - | - | - | - | linPIK3CB | 0.74 ± 0.16 | 0.02 | - |
| AZD5363 | - | - | - | - | linVRK1 | 0.68 ± 0.03 | 0.0003 | 0.016 |
| AZD7762 | - | - | - | - | linBCL11B | 0.42 ± 0.10 | 0.00003 | 0.006 |
| AZD7762 | - | - | - | - | linGFRA1 | 0.23 ± 0.13 | 0.0002 | 0.016 |
| AZD7762 | - | - | - | - | linHIPK3 | 0.45 ± 0.15 | 0.001 | 0.033 |
| AZD7762 | - | - | - | - | linIGF1R | 0.47 ± 0.11 | 0.0004 | 0.017 |
| AZD7762 | - | - | - | - | linMAN1A2 | 0.53 ± 0.13 | 0.0014 | 0.036 |
| AZD7762 | circNCOA3 | 0.74 ± 0.19 | 0.04 | - | linNCOA3 | 0.57 ± 0.19 | 0.01 | - |
| AZD7762 | - | - | - | - | linSNX27 | 0.48 ± 0.20 | 0.004 | - |
| AZD7762 | circVRK1 | 2.26 ± 0.71 | 0.02 | - | linVRK1 | 0.66 ± 0.05 | 0.0001 | 0.011 |
| AZD8055 | - | - | - | - | linMAN1A2 | 0.86 ± 0.03 | 0.0007 | 0.025 |
| DOXORUBICIN | - | - | - | - | linGFRA1 | 1.66 ± 0.52 | 0.046 | - |
| DOXORUBICIN | - | - | - | - | linHIPK3 | 1.58 ± 0.36 | 0.02 | - |
| DOXORUBICIN | circIGF1R | 1.65 ± 0.47 | 0.04 | - | - | - | - | - |
| DOXORUBICIN | circMAN1A2 | 1.59 ± 0.20 | 0.003 | - | linMAN1A2 | 1.71 ± 0.53 | 0.04 | - |
| DOXORUBICIN | circNCOA3 | 1.65 ± 0.42 | 0.03 | - | linNCOA3 | 2.08 ± 0.65 | 0.02 | - |
| DOXORUBICIN | - | - | - | - | linSNX27 | 2.01 ± 0.55 | 0.02 | - |
| DOXORUBICIN | - | - | - | - | linVRK1 | 1.85 ± 0.17 | 0.0004 | 0.017 |
| ERLOTINIB | - | - | - | - | linHIPK3 | 0.72 ± 0.17 | 0.02 | - |
| ERLOTINIB | - | - | - | - | linIGF1R | 0.72 ± 0.17 | 0.02 | - |
| ERLOTINIB | - | - | - | - | linMAN1A2 | 0.79 ± 0.08 | 0.0046 | - |
| GEFITINIB | - | - | - | - | linNCOA3 | 0.59 ± 0.10 | 0.0008 | 0.026 |
| GEFITINIB | - | - | - | - | linVRK1 | 0.79 ± 0.12 | 0.02 | - |
| GSK1070916 | - | - | - | - | linBCL11B | 0.67 ± 0.20 | 0.01 | - |
| GSK1070916 | circGFRA1 | 0.52 ± 0.22 | 0.01 | - | - | - | - | - |
| GSK1070916 | - | - | - | - | linHIPK3 | 0.65 ± 0.22 | 0.02 | - |
| GSK1070916 | - | - | - | - | linIGF1R | 0.74 ± 0.19 | 0.04 | - |
| GSK1070916 | - | - | - | - | linNCOA3 | 0.57 ± 0.19 | 0.008 | - |
| GSK1070916 | - | - | - | - | linVRK1 | 0.83 ± 0.12 | 0.03 | - |
| LEE011 | circ000911 | 1.48 ± 0.35 | 0.04 | - | - | - | - | - |
| LEE011 | circVRK1 | 1.41 ± 0.28 | 0.03 | - | - | - | - | - |
| XL765 | - | - | - | - | linGFRA1 | 0.78 ± 0.15 | 0.03 | - |
| XL765 | - | - | - | - | linNCOA3 | 0.89 ± 0.06 | 0.01 | - |
| XL765 | - | - | - | - | linVRK1 | 1.20 ± 0.12 | 0.02 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terrazzan, A.; Crudele, F.; Corrà, F.; Ancona, P.; Palatini, J.; Bianchi, N.; Volinia, S. Inverse Impact of Cancer Drugs on Circular and Linear RNAs in Breast Cancer Cell Lines. Non-Coding RNA 2023, 9, 32. https://doi.org/10.3390/ncrna9030032
Terrazzan A, Crudele F, Corrà F, Ancona P, Palatini J, Bianchi N, Volinia S. Inverse Impact of Cancer Drugs on Circular and Linear RNAs in Breast Cancer Cell Lines. Non-Coding RNA. 2023; 9(3):32. https://doi.org/10.3390/ncrna9030032
Chicago/Turabian StyleTerrazzan, Anna, Francesca Crudele, Fabio Corrà, Pietro Ancona, Jeffrey Palatini, Nicoletta Bianchi, and Stefano Volinia. 2023. "Inverse Impact of Cancer Drugs on Circular and Linear RNAs in Breast Cancer Cell Lines" Non-Coding RNA 9, no. 3: 32. https://doi.org/10.3390/ncrna9030032