
Targeting miRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update
 , and
, and
Abstract
:1. Introduction
2. Canonical miRNA Biogenesis
3. Dysregulation of Mediators of miRNA Biogenesis
4. The Role of miRNAs in Cancer Therapy Resistance
Approaches to Overcoming Anatomical and Physiological Barriers to Delivery

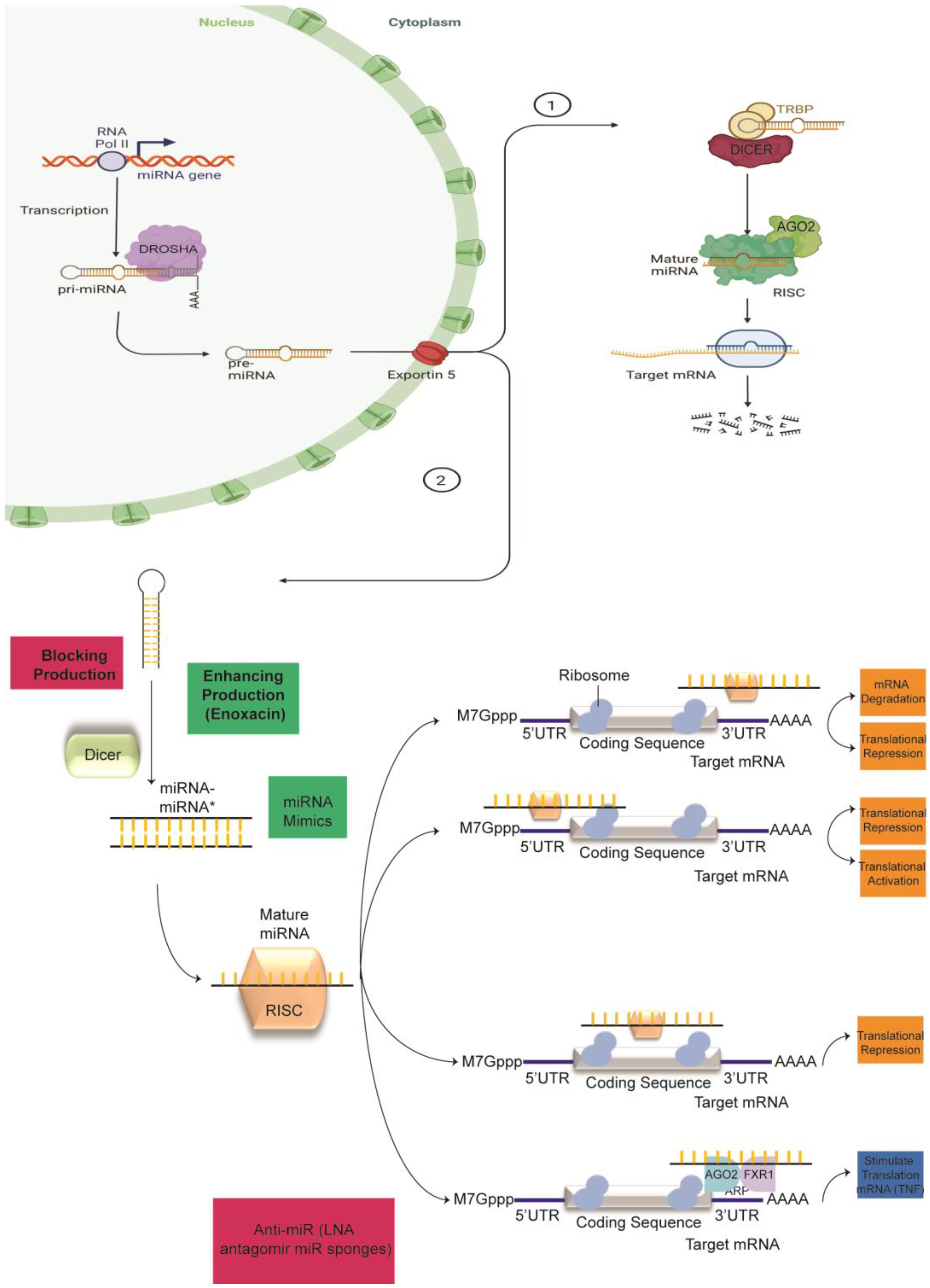{kind=link}
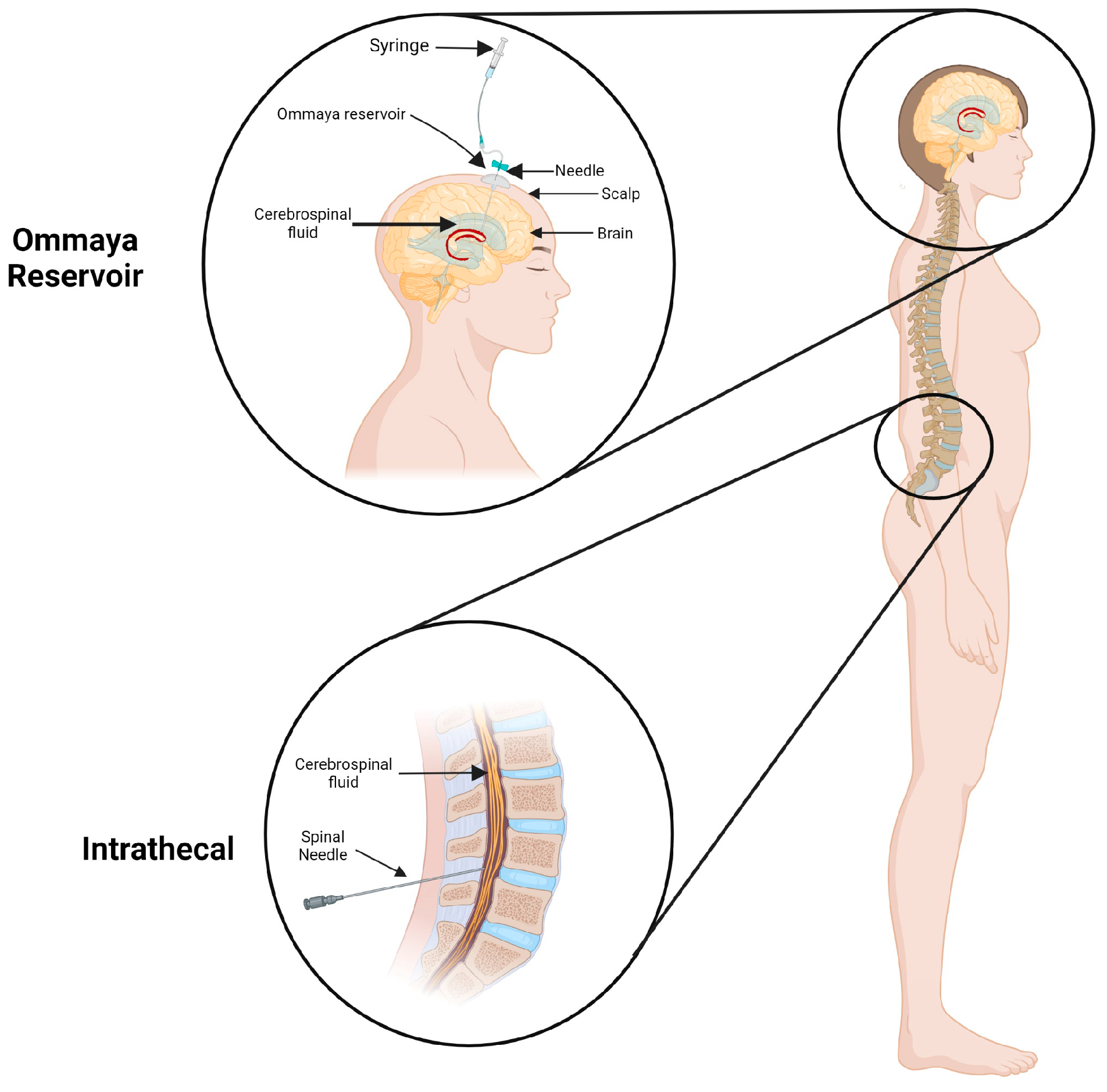{kind=link}
| miRNA | Effect on Resistance 1 | Drug | Cancer Type | Ref. |
|---|---|---|---|---|
| Let7b | Decreases MDR gene expressions, increases sensitivity | PTX | EOC | [86] |
| miR-101-5P | Inhibits growth, increases sensitivity | Trastuzumab and lapatinib | BC | [87] |
| miR-206 | Mediates chemoresistance, proliferation, and apoptosis | 5-FU | CRC | [91] |
| miR-26b | Downregulates P-gp, increases sensitivity | 5-FU | CRC | [92] |
| miR-195 | Inhibits growth, increases sensitivity | Microtubule-targeting agents (MTAs) | NSCLC | [93] |
| miR-493-5p | Increases chemoresistance | Platinum/PARPi | BRCA2-mutated carcinomas | [95,108] |
| miR-506-3p | Decreases β-catenin, decreases sensitivity | PARPi | OC | [96] |
| miR-155 | Induces chemoresistance | Cisplatin, doxorubicin | LC, CLL, ALL | [98] |
| miR-503 | Downregulates Bcl-2, increases sensitivity | Cisplatin | NSCLC | [100] |
| miR-17-92 cluster | Increases chemoresistance | Sorafenib | HCC | [104] |
| miR-19a-3p | Increases metastasis, modulates PTEN expression, increases chemoresistance | Sorafenib | HCC | [105] |
| miR-493-5p | Suppresses METTL3, increases sensitivity | Arabinocytosine | AML | [108] |
| miR-15a-5p | Targets pro-apoptotic genes, induces chemoresistance | Cytarabine and/or daunorubicin | AML | [109] |
| miR-21-5p | Targets pro-apoptotic genes, induces chemoresistance | Cytarabine and/or daunorubicin | AML | [109] |
| miR-138 | Targets EZH2, increases sensitivity | Bortezomib | MM | [112] |
| miR-21 | Induces chemoresistance | Tamoxifen | BC | [115] |
| miR-146a | Increases sensitivity | Tamoxifen | BC | [115] |
| miR-148a | Increases sensitivity | Tamoxifen | BC | [115] |
| miR-34a | Increases sensitivity | Tamoxifen | BC | [115] |
| miR-27a | Increases sensitivity | Tamoxifen | BC | [115] |
| miR-663b | Increases cell proliferation, induces chemoresistance | Tamoxifen | BC | [116] |
5. Systemic Delivery and Recent Developments Using miRNAs as Targets for Anticancer Drug Development
5.1. Lipid-Based miRNA Carrier Systems
5.1.1. Liposomes
5.1.2. Solid Lipid Nanoparticles
5.1.3. Nanostructured Lipid Carriers
6. lncRNAs and Their Opportunities as Targets for Anticancer Therapy
7. The Role of lncRNAs in Cancer Therapy Resistance
8. Clinical Experience
8.1. miRNA Mimics in Clinical Trials
8.2. miRNA Inhibitors in Clinical Trials
9. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattick, J.S. The genetic signatures of noncoding RNAs. PLoS Genet. 2009, 5, e1000459. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhou, Y.; Li, H. LncRNA, miRNA and lncRNA-miRNA interaction in viral infection. Virus Res. 2018, 257, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Katsarou, K.; Rao, A.L.; Tsagris, M.; Kalantidis, K. Infectious long non-coding RNAs. Biochimie 2015, 117, 37–47. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Cipolla, G.A.; de Oliveira, J.C.; Salviano-Silva, A.; Lobo-Alves, S.C.; Lemos, D.S.; Oliveira, L.C.; Jucoski, T.S.; Mathias, C.; Pedroso, G.A.; Zambalde, E.P.; et al. Long Non-Coding RNAs in Multifactorial Diseases: Another Layer of Complexity. Noncoding RNA 2018, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Tong, A.W.; Nemunaitis, J. Modulation of miRNA activity in human cancer: A new paradigm for cancer gene therapy? Cancer Gene Ther. 2008, 15, 341–355. [Google Scholar] [CrossRef]
- Almeida, M.I.; Reis, R.M.; Calin, G.A. MicroRNA history: Discovery, recent applications, and next frontiers. Mutat. Res. 2011, 717, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Song, E.; Lee, S.K.; Wang, J.; Ince, N.; Ouyang, N.; Min, J.; Chen, J.; Shankar, P.; Lieberman, J. RNA interference targeting Fas protects mice from fulminant hepatitis. Nat. Med. 2003, 9, 347–351. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I.; et al. MiR-15a and miR-16-1 cluster functions in human leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef] [Green Version]
- Klein, U.; Lia, M.; Crespo, M.; Siegel, R.; Shen, Q.; Mo, T.; Ambesi-Impiombato, A.; Califano, A.; Migliazza, A.; Bhagat, G.; et al. The DLEU2/miR-15a/16-1 cluster controls B cell proliferation and its deletion leads to chronic lymphocytic leukemia. Cancer Cell 2010, 17, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Calin, G.A.; Lopez-Berestein, G.; Sood, A.K. miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov. 2016, 6, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurha, P. MicroRNAs in cardiovascular disease. Curr. Opin. Cardiol. 2016, 31, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Emine Bayraktar, C.R.-A. MicroRNA Therapeutics: Basic Principles, Barriers and Perspectives. BAOJ Cancer Res. Ther. 2015, 1, 1–5. [Google Scholar]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, E.J.; Yu, A.M.; Lucks, J.B. High-throughput determination of RNA structures. Nat. Rev. Genet. 2018, 19, 615–634. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Cieslik, M.; Chinnaiyan, A.M. Cancer transcriptome profiling at the juncture of clinical translation. Nat. Rev. Genet. 2018, 19, 93–109. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hezroni, H.; Koppstein, D.; Schwartz, M.G.; Avrutin, A.; Bartel, D.P.; Ulitsky, I. Principles of long noncoding RNA evolution derived from direct comparison of transcriptomes in 17 species. Cell Rep. 2015, 11, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Liu, C.G.; Ferracin, M.; Hyslop, T.; Spizzo, R.; Sevignani, C.; Fabbri, M.; Cimmino, A.; Lee, E.J.; Wojcik, S.E.; et al. Ultraconserved regions encoding ncRNAs are altered in human leukemias and carcinomas. Cancer Cell 2007, 12, 215–229. [Google Scholar] [CrossRef]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Z.; Wang, D.; Qiu, C.; Liu, M.; Chen, X.; Zhang, Q.; Yan, G.; Cui, Q. LncRNADisease: A database for long-non-coding RNA-associated diseases. Nucleic Acids Res. 2013, 41, D983–D986. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Wassenegger, M. The role of the RNAi machinery in heterochromatin formation. Cell 2005, 122, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, A.; Sachidanandam, R.; Hannon, G.J.; Carmell, M.A. A germline-specific class of small RNAs binds mammalian Piwi proteins. Nature 2006, 442, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Desset, S.; Buchon, N.; Meignin, C.; Coiffet, M.; Vaury, C. In Drosophila melanogaster the COM locus directs the somatic silencing of two retrotransposons through both Piwi-dependent and -independent pathways. PLoS ONE 2008, 3, e1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, F.; Vaucheret, H.; Rajagopalan, R.; Lepers, C.; Gasciolli, V.; Mallory, A.C.; Hilbert, J.L.; Bartel, D.P.; Crete, P. Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol. Cell 2004, 16, 69–79. [Google Scholar] [CrossRef]
- Brantl, S. Antisense-RNA regulation and RNA interference. Biochim. Biophys. Acta 2002, 1575, 15–25. [Google Scholar] [CrossRef]
- Mochizuki, K.; Gorovsky, M.A. A Dicer-like protein in Tetrahymena has distinct functions in genome rearrangement, chromosome segregation, and meiotic prophase. Genes Dev. 2005, 19, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Calin, G.A. Non-coding RNAs and cancer: New paradigms in oncology. Discov. Med. 2011, 11, 245–254. [Google Scholar]
- Cheah, M.T.; Wachter, A.; Sudarsan, N.; Breaker, R.R. Control of alternative RNA splicing and gene expression by eukaryotic riboswitches. Nature 2007, 447, 497–500. [Google Scholar] [CrossRef]
- Mandal, M.; Boese, B.; Barrick, J.E.; Winkler, W.C.; Breaker, R.R. Riboswitches control fundamental biochemical pathways in Bacillus subtilis and other bacteria. Cell 2003, 113, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Kiss, T. Small nucleolar RNAs: An abundant group of noncoding RNAs with diverse cellular functions. Cell 2002, 109, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.J.; Glazov, E.A.; Cloonan, N.; Simons, C.; Stephen, S.; Faulkner, G.J.; Lassmann, T.; Forrest, A.R.; Grimmond, S.M.; Schroder, K. Tiny RNAs associated with transcription start sites in animals. Nat. Genet. 2009, 41, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Langenberger, D.; Bermudez-Santana, C.; Hertel, J.; Hoffmann, S.; Khaitovich, P.; Stadler, P.F. Evidence for human microRNA-offset RNAs in small RNA sequencing data. Bioinformatics 2009, 25, 2298–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafà, R.; Song, J.; Guo, Z. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [Green Version]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.; Plasterk, R.H.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landthaler, M.; Yalcin, A.; Tuschl, T. The human DiGeorge syndrome critical region gene 8 and Its D. melanogaster homolog are required for miRNA biogenesis. Curr. Biol. 2004, 14, 2162–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnsack, M.T.; Czaplinski, K.; Gorlich, D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Lund, E.; Guttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear export of microRNA precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Ketting, R.F.; Fischer, S.E.; Bernstein, E.; Sijen, T.; Hannon, G.J.; Plasterk, R.H. Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Genes Dev. 2001, 15, 2654–2659. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef]
- Parker, G.S.; Eckert, D.M.; Bass, B.L. RDE-4 preferentially binds long dsRNA and its dimerization is necessary for cleavage of dsRNA to siRNA. RNA 2006, 12, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2018, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, W.D.; Priest, J.R.; Duchaine, T.F. DICER1: Mutations, microRNAs and mechanisms. Nat. Rev. Cancer 2014, 14, 662–672. [Google Scholar] [CrossRef] [PubMed]
- van den Beucken, T.; Koch, E.; Chu, K.; Rupaimoole, R.; Prickaerts, P.; Adriaens, M.; Voncken, J.W.; Harris, A.L.; Buffa, F.M.; Haider, S.; et al. Hypoxia promotes stem cell phenotypes and poor prognosis through epigenetic regulation of DICER. Nat. Commun. 2014, 5, 5203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, J.H.; Bisogna, M.; Hoang, L.N.; Olvera, N.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Sood, A.K.; Levine, D.A.; Jelinic, P. miR-200c-driven Mesenchymal-To-Epithelial Transition is a Therapeutic Target in Uterine Carcinosarcomas. Sci. Rep. 2017, 7, 3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.Y.; Rupaimoole, R.; Shen, F.; Pradeep, S.; Pecot, C.V.; Ivan, C.; Nagaraja, A.S.; Gharpure, K.M.; Pham, E.; Hatakeyama, H.; et al. A miR-192-EGR1-HOXB9 regulatory network controls the angiogenic switch in cancer. Nat. Commun. 2016, 7, 11169. [Google Scholar] [CrossRef] [Green Version]
- Landen, C.N., Jr.; Chavez-Reyes, A.; Bucana, C.; Schmandt, R.; Deavers, M.T.; Lopez-Berestein, G.; Sood, A.K. Therapeutic EphA2 gene targeting in vivo using neutral liposomal small interfering RNA delivery. Cancer Res. 2005, 65, 6910–6918. [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, R.; Ivan, C.; Bayraktar, E.; Kanlikilicer, P.; Kabil, N.; Kahraman, N.; Mokhlis, H.A.; Karakas, D.; Rodriguez-Aguayo, C.; Arslan, A.; et al. Dual suppressive effect of microRNA-34a on the FOXM1/eEF2-kinase axis regulates triple-negative breast cancer growth and invasion. Clin. Cancer Res. 2018, 24, 4225–4241. [Google Scholar] [CrossRef] [Green Version]
- Kanlikilicer, P.; Rashed, M.H.; Bayraktar, R.; Mitra, R.; Ivan, C.; Aslan, B.; Zhang, X.; Filant, J.; Silva, A.M.; Rodriguez-Aguayo, C.; et al. Ubiquitous Release of Exosomal Tumor Suppressor miR-6126 from Ovarian Cancer Cells. Cancer Res. 2016, 76, 7194–7207. [Google Scholar] [CrossRef] [Green Version]
- Rashed, M.H.; Kanlikilicer, P.; Rodriguez-Aguayo, C.; Pichler, M.; Bayraktar, R.; Bayraktar, E.; Ivan, C.; Filant, J.; Silva, A.; Aslan, B.; et al. Exosomal miR-940 maintains SRC-mediated oncogenic activity in cancer cells: A possible role for exosomal disposal of tumor suppressor miRNAs. Oncotarget 2017, 8, 20145–20164. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zhang, Y.; Meng, Y.P.; Bo, L.S.; Ke, W.B. miR-542-3p Appended Sorafenib/All-trans Retinoic Acid (ATRA)-Loaded Lipid Nanoparticles to Enhance the Anticancer Efficacy in Gastric Cancers. Pharm. Res. 2017, 34, 2710–2719. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lujambio, A.; Lowe, S.W. The microcosmos of cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. microRNAs in cancer management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Tan, C. Combination of microRNA therapeutics with small-molecule anticancer drugs: Mechanism of action and co-delivery nanocarriers. Adv. Drug Deliv. Rev. 2015, 81, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.S.; Tekade, R.K.; Chougule, M.B. Nanocarrier mediated delivery of siRNA/miRNA in combination with chemotherapeutic agents for cancer therapy: Current progress and advances. J. Control. Release 2014, 194, 238–256. [Google Scholar] [CrossRef] [Green Version]
- Gandham, S.K.; Rao, M.; Shah, A.; Trivedi, M.S.; Amiji, M.M. Combination microRNA-based cellular reprogramming with paclitaxel enhances therapeutic efficacy in a relapsed and multidrug-resistant model of epithelial ovarian cancer. Mol. Ther.-Oncolytics 2022, 25, 57–68. [Google Scholar] [CrossRef]
- Normann, L.S.; Aure, M.R.; Leivonen, S.-K.; Haugen, M.H.; Hongisto, V.; Kristensen, V.N.; Mælandsmo, G.M.; Sahlberg, K.K. MicroRNA in combination with HER2-targeting drugs reduces breast cancer cell viability in vitro. Sci. Rep. 2021, 11, 10893. [Google Scholar] [CrossRef]
- Norouzi-Barough, L.; Sarookhani, M.R.; Sharifi, M.; Moghbelinejad, S.; Jangjoo, S.; Salehi, R. Molecular mechanisms of drug resistance in ovarian cancer. J. Cell Physiol. 2018, 233, 4546–4562. [Google Scholar] [CrossRef]
- Mader, R.M.; Muller, M.; Steger, G.G. Resistance to 5-fluorouracil. Gen. Pharm. 1998, 31, 661–666. [Google Scholar] [CrossRef]
- Sobrero, A.F.; Aschele, C.; Bertino, J.R. Fluorouracil in colorectal cancer—A tale of two drugs: Implications for biochemical modulation. J. Clin. Oncol. 1997, 15, 368–381. [Google Scholar] [CrossRef]
- Meng, X.; Fu, R. miR-206 regulates 5-FU resistance by targeting Bcl-2 in colon cancer cells. Onco. Targets Ther. 2018, 11, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Pouya, F.D.; Gazouli, M.; Rasmi, Y.; Lampropoulou, D.I.; Nemati, M. MicroRNAs and drug resistance in colorectal cancer with special focus on 5-fluorouracil. Mol. Biol. Rep. 2022, 49, 5165–5178. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, Y.; Ma, X.; Pertsemlidis, A. miR-195 Potentiates the Efficacy of Microtubule-Targeting Agents in Non-Small Cell Lung Cancer. Cancer Lett. 2018, 427, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Mihanfar, A.; Fattahi, A.; Nejabati, H.R. MicroRNA-mediated drug resistance in ovarian cancer. J. Cell. Physiol. 2019, 234, 3180–3191. [Google Scholar] [CrossRef] [PubMed]
- Meghani, K.; Fuchs, W.; Detappe, A.; Drane, P.; Gogola, E.; Rottenberg, S.; Jonkers, J.; Matulonis, U.; Swisher, E.M.; Konstantinopoulos, P.A.; et al. Multifaceted Impact of MicroRNA 493-5p on Genome-Stabilizing Pathways Induces Platinum and PARP Inhibitor Resistance in BRCA2-Mutated Carcinomas. Cell Rep. 2018, 23, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wu, J.; Dong, X.; Zhang, J.; Meng, C.; Liu, G. MicroRNA-506-3p increases the response to PARP inhibitors and cisplatin by targeting EZH2/β-catenin in serous ovarian cancers. Transl. Oncol. 2021, 14, 100987. [Google Scholar] [CrossRef] [PubMed]
- Aas, T.; Borresen, A.L.; Geisler, S.; Smith-Sorensen, B.; Johnsen, H.; Varhaug, J.E.; Akslen, L.A.; Lonning, P.E. Specific P53 mutations are associated with de novo resistance to doxorubicin in breast cancer patients. Nat. Med. 1996, 2, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Van Roosbroeck, K.; Fanini, F.; Setoyama, T.; Ivan, C.; Rodriguez-Aguayo, C.; Fuentes-Mattei, E.; Xiao, L.; Vannini, I.; Redis, R.S.; D’Abundo, L.; et al. Combining Anti-Mir-155 with Chemotherapy for the Treatment of Lung Cancers. Clin. Cancer Res. 2017, 23, 2891–2904. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Lan, H.; Jiang, X.; Zeng, D.; Xiao, S. Bcl-2 family: Novel insight into individualized therapy for ovarian cancer. Int. J. Mol. Med. 2020, 46, 1255–1265. [Google Scholar] [CrossRef]
- Qiu, T.; Zhou, L.; Wang, T.; Xu, J.; Wang, J.; Chen, W.; Zhou, X.; Huang, Z.; Zhu, W.; Shu, Y.; et al. miR-503 regulates the resistance of non-small cell lung cancer cells to cisplatin by targeting Bcl-2. Int. J. Mol. Med. 2013, 32, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, H.V.; Meyer-Hermann, M. The molecular basis of synergism between carboplatin and ABT-737 therapy targeting ovarian carcinomas. Cancer Res. 2011, 71, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, M.; Croce, C.M. MicroRNAs as therapeutic targets in chemoresistance. Drug Resist. Updat. 2013, 16, 47–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, F.M.; Naz, A.; Obaid, A.; Ikram, A.; Ali, A.; Ahmad, J.; Naveed, A.K.; Janjua, H.A. MicroRNA pharmacogenomics based integrated model of miR-17-92 cluster in sorafenib resistant HCC cells reveals a strategy to forestall drug resistance. Sci. Rep. 2017, 7, 11448. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Wang, Y. The diagnostic and prognostic value of the miR-17-92 cluster in hepatocellular carcinoma: A meta-analysis. Front. Genet. 2022, 13, 2306. [Google Scholar] [CrossRef]
- Liu, B.; Ma, X.; Liu, Q.; Xiao, Y.; Pan, S.; Jia, L. Aberrant mannosylation profile and FTX/miR-342/ALG3-axis contribute to development of drug resistance in acute myeloid leukemia. Cell Death Dis. 2018, 9, 688. [Google Scholar] [CrossRef] [Green Version]
- Dohner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Chen, Y.; Shi, L.; Li, M.; Li, L.; Wang, S.; Wang, C. Tumor-suppressive MEG3 induces microRNA-493-5p expression to reduce arabinocytosine chemoresistance of acute myeloid leukemia cells by downregulating the METTL3/MYC axis. J. Transl. Med. 2022, 20, 288. [Google Scholar] [CrossRef]
- Vandewalle, V.; Essaghir, A.; Bollaert, E.; Lenglez, S.; Graux, C.; Schoemans, H.; Saussoy, P.; Michaux, L.; Valk, P.J.; Demoulin, J.B. miR-15a-5p and miR-21-5p contribute to chemoresistance in cytogenetically normal acute myeloid leukaemia by targeting PDCD4, ARL2 and BTG2. J. Cell. Mol. Med. 2021, 25, 575–585. [Google Scholar] [CrossRef]
- Moloudizargari, M.; Abdollahi, M.; Asghari, M.H.; Zimta, A.A.; Neagoe, I.B.; Nabavi, S.M. The emerging role of exosomes in multiple myeloma. Blood Rev. 2019, 38, 100595. [Google Scholar] [CrossRef]
- Solimando, A.G.; Malerba, E.; Leone, P.; Prete, M.; Terragna, C.; Cavo, M.; Racanelli, V. Drug resistance in multiple myeloma: Soldiers and weapons in the bone marrow niche. Front. Oncol. 2022, 12, 973836. [Google Scholar] [CrossRef] [PubMed]
- Rastgoo, N.; Pourabdollah, M.; Abdi, J.; Reece, D.; Chang, H. Dysregulation of EZH2/miR-138 axis contributes to drug resistance in multiple myeloma by downregulating RBPMS. Leukemia 2018, 32, 2471–2482. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, A.; Johnstone, K.; McCart Reed, A.; Simpson, P.; Lakhani, S. Hereditary breast cancer: Syndromes, tumour pathology and molecular testing. Histopathology 2023, 82, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Ribas, R.; Pancholi, S.; Rani, A.; Schuster, E.; Guest, S.K.; Nikitorowicz-Buniak, J.; Simigdala, N.; Thornhill, A.; Avogadri-Connors, F.; Cutler, R.E., Jr.; et al. Targeting tumour re-wiring by triple blockade of mTORC1, epidermal growth factor, and oestrogen receptor signalling pathways in endocrine-resistant breast cancer. Breast Cancer Res. 2018, 20, 44. [Google Scholar] [CrossRef]
- Ye, P.; Fang, C.; Zeng, H.; Shi, Y.; Pan, Z.; An, N.; He, K.; Zhang, L.; Long, X. Differential microRNA expression profiles in tamoxifen-resistant human breast cancer cell lines induced by two methods. Oncol. Lett. 2018, 15, 3532–3539. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Cheng, L.; Hu, P.; Liu, R. MicroRNA663b mediates TAM resistance in breast cancer by modulating TP73 expression. Mol. Med. Rep. 2018, 18, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Dowdy, S.F. Overcoming cellular barriers for RNA therapeutics. Nat. Biotechnol. 2017, 35, 222–229. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Mainini, F.; Eccles, M.R. Lipid and Polymer-Based Nanoparticle siRNA Delivery Systems for Cancer Therapy. Molecules 2020, 25, 2692. [Google Scholar] [CrossRef]
- Dalpke, A.; Helm, M. RNA mediated Toll-like receptor stimulation in health and disease. RNA Biol. 2012, 9, 828–842. [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, R.; Bertilaccio, M.T.S.; Calin, G.A. The Interaction Between Two Worlds: MicroRNAs and Toll-Like Receptors. Front. Immunol. 2019, 10, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirón-Barroso, S.; Correia, J.S.; Frampton, A.E.; Lythgoe, M.P.; Clark, J.; Tookman, L.; Ottaviani, S.; Castellano, L.; Porter, A.E.; Georgiou, T.K.; et al. Polymeric Carriers for Delivery of RNA Cancer Therapeutics. Noncoding RNA 2022, 8, 58. [Google Scholar] [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, S.M.; Aartsma-Rus, A.; Alves, S.; Borgos, S.E.; Buijsen, R.A.M.; Collin, R.W.J.; Covello, G.; Denti, M.A.; Desviat, L.R.; Echevarría, L.; et al. Delivery of oligonucleotide-based therapeutics: Challenges and opportunities. EMBO Mol. Med. 2021, 13, e13243. [Google Scholar] [CrossRef]
- Labatut, A.E.; Mattheolabakis, G. Non-viral based miR delivery and recent developments. Eur. J. Pharm. Biopharm. 2018, 128, 82–90. [Google Scholar] [CrossRef]
- Cristian Rodriguez-Aguayo, A.C.-R.; Gabriel Lopez-Berestein Anil, K. Sood. RNAi in cancer therapy. In Advanced Delivery and Therapeutic Applications of RNAi; Kun Cheng, R.I.M., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 271–307. [Google Scholar]
- Grimm, D. Small silencing RNAs: State-of-the-art. Adv. Drug Deliv. Rev. 2009, 61, 672–703. [Google Scholar] [CrossRef]
- Giacca, M.; Zacchigna, S. Virus-mediated gene delivery for human gene therapy. J. Control. Release 2012, 161, 377–388. [Google Scholar] [CrossRef]
- Chen, X.; Mangala, L.S.; Rodriguez-Aguayo, C.; Kong, X.; Lopez-Berestein, G.; Sood, A.K. RNA interference-based therapy and its delivery systems. Cancer Metastasis Rev. 2018, 37, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Vilaça, A.; de Windt, L.J.; Fernandes, H.; Ferreira, L. Strategies and challenges for non-viral delivery of non-coding RNAs to the heart. Trends Mol. Med. 2023, 29, 70–91. [Google Scholar] [CrossRef]
- Ewert, K.; Slack, N.L.; Ahmad, A.; Evans, H.M.; Lin, A.J.; Samuel, C.E.; Safinya, C.R. Cationic lipid-DNA complexes for gene therapy: Understanding the relationship between complex structure and gene delivery pathways at the molecular level. Curr. Med. Chem. 2004, 11, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wang, G.; He, B.; Li, L.; Li, C.; Lai, Y.; Xu, X.; Gu, Z. Cationic lipid-coated PEI/DNA polyplexes with improved efficiency and reduced cytotoxicity for gene delivery into mesenchymal stem cells. Int. J. Nanomed. 2012, 7, 4637–4648. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Gao, D.Y.; Huang, L. In vivo delivery of miRNAs for cancer therapy: Challenges and strategies. Adv. Drug Deliv. Rev. 2015, 81, 128–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raemdonck, K.; Vandenbroucke, R.E.; Demeester, J.; Sanders, N.N.; De Smedt, S.C. Maintaining the silence: Reflections on long-term RNAi. Drug Discov. Today 2008, 13, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Stylianopoulos, T.; Jain, R.K. Combining two strategies to improve perfusion and drug delivery in solid tumors. Proc. Natl. Acad. Sci. USA 2013, 110, 18632–18637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: Review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Mattheolabakis, G.; Rigas, B.; Constantinides, P.P. Nanodelivery strategies in cancer chemotherapy: Biological rationale and pharmaceutical perspectives. Nanomedicine 2012, 7, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.M.; Pedroso de Lima, M.C. MicroRNAs as Molecular Targets for Cancer Therapy: On the Modulation of MicroRNA Expression. Pharmaceuticals 2013, 6, 1195–1220. [Google Scholar] [CrossRef]
- Chapoy-Villanueva, H.; Martinez-Carlin, I.; Lopez-Berestein, G.; Chavez-Reyes, A. Therapeutic silencing of HPV 16 E7 by systemic administration of siRNA-neutral DOPC nanoliposome in a murine cervical cancer model with obesity. J. BUON 2015, 20, 1471–1479. [Google Scholar]
- Rupaimoole, R.; Ivan, C.; Yang, D.; Gharpure, K.M.; Wu, S.Y.; Pecot, C.V.; Previs, R.A.; Nagaraja, A.S.; Armaiz-Pena, G.N.; McGuire, M.; et al. Hypoxia-upregulated microRNA-630 targets Dicer, leading to increased tumor progression. Oncogene 2016, 35, 4312–4320. [Google Scholar] [CrossRef] [Green Version]
- Muller, R.H.; Mader, K.; Gohla, S. Solid lipid nanoparticles (SLN) for controlled drug delivery—A review of the state of the art. Eur. J. Pharm. Biopharm. 2000, 50, 161–177. [Google Scholar] [CrossRef]
- Shi, S.J.; Zhong, Z.R.; Liu, J.; Zhang, Z.R.; Sun, X.; Gong, T. Solid lipid nanoparticles loaded with anti-microRNA oligonucleotides (AMOs) for suppression of microRNA-21 functions in human lung cancer cells. Pharm. Res. 2012, 29, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.L.; Bendayan, R.; Rauth, A.M.; Li, Y.; Wu, X.Y. Chemotherapy with anticancer drugs encapsulated in solid lipid nanoparticles. Adv. Drug Deliv. Rev. 2007, 59, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.L.; Bendayan, R.; Rauth, A.M.; Wu, X.Y. Development of solid lipid nanoparticles containing ionically complexed chemotherapeutic drugs and chemosensitizers. J. Pharm. Sci. 2004, 93, 1993–2008. [Google Scholar] [CrossRef] [PubMed]
- Subedi, R.K.; Kang, K.W.; Choi, H.K. Preparation and characterization of solid lipid nanoparticles loaded with doxorubicin. Eur. J. Pharm. Sci. 2009, 37, 508–513. [Google Scholar] [CrossRef]
- Lobovkina, T.; Jacobson, G.B.; Gonzalez-Gonzalez, E.; Hickerson, R.P.; Leake, D.; Kaspar, R.L.; Contag, C.H.; Zare, R.N. In vivo sustained release of siRNA from solid lipid nanoparticles. ACS Nano 2011, 5, 9977–9983. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Meng, T.; Yuan, M.; Wen, L.; Cheng, B.; Liu, N.; Huang, X.; Hong, Y.; Yuan, H.; Hu, F. MicroRNA-200c delivered by solid lipid nanoparticles enhances the effect of paclitaxel on breast cancer stem cell. Int. J. Nanomed. 2016, 11, 6713–6725. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Han, L.; Deng, L.; Zhang, Y.; Shen, H.; Gong, T.; Zhang, Z.; Sun, X. Dual drugs (microRNA-34a and paclitaxel)-loaded functional solid lipid nanoparticles for synergistic cancer cell suppression. J. Control. Release 2014, 194, 228–237. [Google Scholar] [CrossRef]
- Chauhan, I.; Yasir, M.; Verma, M.; Singh, A.P. Nanostructured lipid carriers: A groundbreaking approach for transdermal drug delivery. Adv. Pharm. Bull. 2020, 10, 150. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Jia, L.; Chu, F.; Zhou, Y.; He, Z.; Guo, M.; Chen, C.; Xu, L. Nanostructured lipid carriers for MicroRNA delivery in tumor gene therapy. Cancer Cell Int. 2018, 18, 101. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, P.; Gidwani, B.; Vyas, A. Nanostructured lipid carriers and their current application in targeted drug delivery. Artif. Cells Nanomed. Biotechnol. 2016, 44, 27–40. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, X.; Zhang, X.; Liu, B.; Huang, L. Nanoparticles modified with tumor-targeting scFv deliver siRNA and miRNA for cancer therapy. Mol. Ther. 2010, 18, 1650–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, L.; Zhang, M.; Datta, J.; Xie, X.; Su, T.; Li, H.; Teknos, T.N.; Pan, Q. Lipid-based nanoparticle delivery of Pre-miR-107 inhibits the tumorigenicity of head and neck squamous cell carcinoma. Mol. Ther. 2012, 20, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Peng, F.; Zhou, T.; Huang, Y.; Zhang, L.; Ye, P.; Lu, M.; Yang, G.; Gai, Y.; Yang, T. Targeted delivery of chemically modified anti-miR-221 to hepatocellular carcinoma with negatively charged liposomes. Int. J. Nanomed. 2015, 10, 4825. [Google Scholar]
- Unfried, J.P.; Serrano, G.; Suárez, B.; Sangro, P.; Ferretti, V.; Prior, C.; Boix, L.; Bruix, J.; Sangro, B.; Segura, V. Identification of Coding and Long Noncoding RNAs Differentially Expressed in Tumors and Preferentially Expressed in Healthy TissuesTumors Upregulate Transcripts from Healthy Testis and Brain. Cancer Res. 2019, 79, 5167–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toden, S.; Zumwalt, T.J.; Goel, A. Non-coding RNAs and potential therapeutic targeting in cancer. Biochim. Biophys. Acta BBA-Rev. Cancer 2021, 1875, 188491. [Google Scholar] [CrossRef]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, K.; Zhou, H.; Wu, Y.; Li, C.; Liu, Y.; Liu, Z.; Xu, Q.; Liu, S.; Xiao, D. Role of non-coding RNAs and RNA modifiers in cancer therapy resistance. Mol. Cancer 2020, 19, 47. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.; Sadeghi, S.; Tabatabaeian, H. Battling chemoresistance in cancer: Root causes and strategies to uproot them. Int. J. Mol. Sci. 2021, 22, 9451. [Google Scholar] [CrossRef]
- Singh, D.; Assaraf, Y.G.; Gacche, R.N. Long non-coding RNA mediated drug resistance in breast cancer. Drug Resist. Updat. 2022, 63, 100851. [Google Scholar] [CrossRef]
- Chang, L.; Hu, Z.; Zhou, Z.; Zhang, H. Linc00518 contributes to multidrug resistance through regulating the MiR-199a/MRP1 axis in breast cancer. Cell. Physiol. Biochem. 2018, 48, 16–28. [Google Scholar] [CrossRef]
- Huang, P.; Li, F.; Li, L.; You, Y.; Luo, S.; Dong, Z.; Gao, Q.; Wu, S.; Brünner, N.; Stenvang, J. lncRNA profile study reveals the mRNAs and lncRNAs associated with docetaxel resistance in breast cancer cells. Sci. Rep. 2018, 8, 17970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Qin, Z.; Cai, S.; Yu, L.; Hu, H.; Zeng, S. The role of non-coding RNAs in ABC transporters regulation and their clinical implications of multidrug resistance in cancer. Expert Opin. Drug Metab. Toxicol. 2021, 17, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Wang, W.; Fu, B.; Pang, Y.; Lou, Y.; Li, H. Increased lncRNA HOTAIR expression promotes the chemoresistance of multiple myeloma to dexamethasone by regulating cell viability and apoptosis by mediating the JAK2/STAT3 signaling pathway. Mol. Med. Rep. 2019, 20, 3917–3923. [Google Scholar] [CrossRef]
- Yuan, P.; Cao, W.; Zang, Q.; Li, G.; Guo, X.; Fan, J. The HIF-2α-MALAT1-miR-216b axis regulates multi-drug resistance of hepatocellular carcinoma cells via modulating autophagy. Biochem. Biophys. Res. Commun. 2016, 478, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; He, J.; Zhang, D. Suppression of long non-coding RNA CCAT2 improves tamoxifen-resistant breast cancer cells’ response to tamoxifen. Mol. Biol. 2016, 50, 821–827. [Google Scholar] [CrossRef]
- Cai, Y.; Dong, Z.; Wang, J. LncRNA NNT-AS1 is a major mediator of cisplatin chemoresistance in non-small cell lung cancer through MAPK/Slug pathway. Eur. Rev. Med. Pharm. Sci. 2018, 22, 4879–4887. [Google Scholar]
- Du, P.; Hu, C.e.; Qin, Y.; Zhao, J.; Patel, R.; Fu, Y.; Zhu, M.; Zhang, W.; Huang, G. LncRNA PVT1 mediates antiapoptosis and 5-fluorouracil resistance via increasing Bcl2 expression in gastric cancer. J. Oncol. 2019, 2019, 9325407. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Han, X.; Hu, Z.; Chen, L. The PVT1/miR-216b/Beclin-1 regulates cisplatin sensitivity of NSCLC cells via modulating autophagy and apoptosis. Cancer Chemother. Pharmacol. 2019, 83, 921–931. [Google Scholar] [CrossRef]
- Han, Y.; Gao, X.; Wu, N.; Jin, Y.; Zhou, H.; Wang, W.; Liu, H.; Chu, Y.; Cao, J.; Jiang, M. Long noncoding RNA LINC00239 inhibits ferroptosis in colorectal cancer by binding to Keap1 to stabilize Nrf2. Cell Death Dis. 2022, 13, 742. [Google Scholar] [CrossRef]
- Xie, W.; Chu, M.; Song, G.; Zuo, Z.; Han, Z.; Chen, C.; Li, Y.; Wang, Z.-W. Emerging roles of long noncoding RNAs in chemoresistance of pancreatic cancer. Semin. Cancer Biol. 2022, 83, 303–318. [Google Scholar] [CrossRef]
- Chen, P.; Wang, M.; Wang, C. Qingyihuaji formula reverses gemcitabine resistant human pancreatic cancer through regulate lncRNA AB209630/miR-373/EphB2-NANOG signals. Biosci. Rep. 2019, 39, BSR20190610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Su, H.; Zou, C.; Liang, X.; Fei, Z. Ginsenoside Rg3 suppresses the growth of gemcitabine-resistant pancreatic cancer cells by upregulating lncRNA-CASC2 and activating PTEN signaling. J. Biochem. Mol. Toxicol. 2020, 34, e22480. [Google Scholar] [CrossRef]
- Liu, B.; Wu, S.; Ma, J.; Yan, S.; Xiao, Z.; Wan, L.; Zhang, F.; Shang, M.; Mao, A. lncRNA GAS5 reverses EMT and tumor stem cell-mediated gemcitabine resistance and metastasis by targeting miR-221/SOCS3 in pancreatic cancer. Mol. Ther.-Nucleic Acids 2018, 13, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Y.; Feng, H.-M. MEG3 suppresses human pancreatic neuroendocrine tumor cells growth and metastasis by down-regulation of Mir-183. Cell. Physiol. Biochem. 2017, 44, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, F.; Zhang, Q.; Dong, Z.; Hu, J.; Ma, Z. LncRNA HOTTIP participates in cisplatin resistance of tumor cells by regulating miR-137 expression in pancreatic cancer. OncoTargets Ther. 2020, 13, 2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-z.; Xu, F.; Zhou, T.; Zhao, X.; McDonald, J.M.; Chen, Y. The long non-coding RNA HOTAIR enhances pancreatic cancer resistance to TNF-related apoptosis-inducing ligand. J. Biol. Chem. 2017, 292, 10390–10397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Yi, C.; Yi, Y.; Qin, W.; Yan, Y.; Dong, X.; Zhang, X.; Huang, Y.; Zhang, R.; Wei, J. LncRNA PVT1 promotes gemcitabine resistance of pancreatic cancer via activating Wnt/β-catenin and autophagy pathway through modulating the miR-619-5p/Pygo2 and miR-619-5p/ATG14 axes. Mol. Cancer 2020, 19, 118. [Google Scholar] [CrossRef]
- Lin, C.; Zheng, M.; Yang, Y.; Chen, Y.; Zhang, X.; Zhu, L.; Zhang, H. Knockdown of lncRNA ACTA2-AS1 reverses cisplatin resistance of ovarian cancer cells via inhibition of miR-378a-3p-regulated Wnt5a. Bioengineered 2022, 13, 9829–9838. [Google Scholar] [CrossRef]
- Wang, D.Y.; Li, N.; Cui, Y.L. Long Non-coding RNA CCAT1 Sponges miR-454 to Promote Chemoresistance of Ovarian Cancer Cells to Cisplatin by Regulation of Surviving. Cancer Res. Treat. 2020, 52, 798–814. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther.-Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayraktar, R.; Van Roosbroeck, K. miR-155 in cancer drug resistance and as target for miRNA-based therapeutics. Cancer Metastasis Rev. 2018, 37, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, R.; Van Roosbroeck, K.; Calin, G.A. Cell-to-cell communication: microRNAs as hormones. Mol. Oncol. 2017, 11, 1673–1686. [Google Scholar] [CrossRef] [Green Version]
- Robb, T.; Reid, G.; Blenkiron, C. Exploiting microRNAs As Cancer Therapeutics. Target Oncol. 2017, 12, 163–178. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. The biological functions of miRNAs: Lessons from in vivo studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [Green Version]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Thakral, S.; Ghoshal, K. miR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr. Gene Ther. 2015, 15, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Coulouarn, C.; Factor, V.M.; Andersen, J.B.; Durkin, M.E.; Thorgeirsson, S.S. Loss of miR-122 expression in liver cancer correlates with suppression of the hepatic phenotype and gain of metastatic properties. Oncogene 2009, 28, 3526–3536. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, M.B.; Baumert, T.F. Clinical development of hepatitis C virus host-targeting agents. Lancet 2017, 389, 674–675. [Google Scholar] [CrossRef] [Green Version]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Ree, M.H.; de Vree, J.M.; Stelma, F.; Willemse, S.; van der Valk, M.; Rietdijk, S.; Molenkamp, R.; Schinkel, J.; van Nuenen, A.C.; Beuers, U.; et al. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: A phase 1B, double-blind, randomised controlled trial. Lancet 2017, 389, 709–717. [Google Scholar] [CrossRef]
- Wallace, J.A.; Kagele, D.A.; Eiring, A.M.; Kim, C.N.; Hu, R.; Runtsch, M.C.; Alexander, M.; Huffaker, T.B.; Lee, S.H.; Patel, A.B.; et al. miR-155 promotes FLT3-ITD-induced myeloproliferative disease through inhibition of the interferon response. Blood 2017, 129, 3074–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Yi, M.; Kim, C.H.; Deng, C.; Li, Y.; Medina, D.; Stephens, R.M.; Green, J.E. Integrated miRNA and mRNA expression profiling of mouse mammary tumor models identifies miRNA signatures associated with mammary tumor lineage. Genome Biol. 2011, 12, R77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, H.; Guo, J.; Li, S.; Jiang, D. miR-155 inhibitor reduces the proliferation and migration in osteosarcoma MG-63 cells. Exp. Ther. Med. 2014, 8, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Luo, X.; Gu, C.; Fei, J. Seed targeting with tiny anti-miR-155 inhibits malignant progression of multiple myeloma cells. J. Drug Target 2015, 23, 59–66. [Google Scholar] [CrossRef]
- Meng, W.; Jiang, L.; Lu, L.; Hu, H.; Yu, H.; Ding, D.; Xiao, K.; Zheng, W.; Guo, H.; Ma, W. Anti-miR-155 oligonucleotide enhances chemosensitivity of U251 cell to taxol by inducing apoptosis. Cell Biol. Int. 2012, 36, 653–659. [Google Scholar] [CrossRef]
- Choi, C.H.; Park, Y.A.; Choi, J.J.; Song, T.; Song, S.Y.; Lee, Y.Y.; Lee, J.W.; Kim, T.J.; Kim, B.G.; Bae, D.S. Angiotensin II type I receptor and miR-155 in endometrial cancers: Synergistic antiproliferative effects of anti-miR-155 and losartan on endometrial cancer cells. Gynecol. Oncol. 2012, 126, 124–131. [Google Scholar] [CrossRef]


| Class | Size (Nucleotides) | Functions | Mechanisms | Origin | Model Organisms | Ref. |
|---|---|---|---|---|---|---|
| siRNAs | 21–25 | Regulate gene expression, provide an antiviral response, and restrict transposons | Degrade RNA, restrict transposon | Intergenic regions, exons, and introns | Caenorhabditis elegans, Drosophila melanogaster, Schizosaccharomyces pombe, Arabidopsis thaliana, Oryza sativa (rice) | [14] |
| endo-siRNAs | 21–25 | Restrict transposons, regulate mRNAs and heterochromatin | Degrade RNA | Transposable elements, pseudogenes | D. melanogaster, mammals | [42] |
| miRNAs | 21–25 | Regulate gene expression at the post-transcriptional level | Block translation, degrade RNA | Intergenic regions, and introns | C. elegans, D. melanogaster, S. pombe, A. thaliana, O. sativa, mammals | [11] |
| piRNAs | 24–31 | Regulate germline development and integrity and silence selfish DNA | Unknown | Defective transposon sequences and other repeats | C. elegans, D. melanogaster, Danio rerio, | [43] |
| ra-siRNAs | 23–28 | Remodel chromatin and silence transcriptional gene | Unknown | Repeated sequence elements (subset of piRNAs) | C. elegans, D. melanogaster, S. pombe, Trypanosoma brucei, D. rerio, A. thaliana | [44] |
| ta-siRNAs | 21–22 | Cleave endogenous mRNAs in a trans-acting manner | Degrade RNA | Non-coding endogenous transcripts | D. melanogaster, S. pombe, A. thaliana, | [45] |
| natRNAs | 35–150 | Regulate gene expression at the post-transcriptional level | Degrade RNA | Opposite strand of the same DNA molecule | All organisms | [46] |
| scnRNAs | 26–30 | Regulate chromatin structure | Eliminate DNA | Meiotic micronuclei | Tetrahymena thermophila and Paramecium tetraurelia | [47] |
| lncRNAs | 200–100 | Possibly play roles in epigenetic mechanisms and gene regulation | Degrade RNA and modify epigenetic mechanisms | Intergenic regions, exons, introns | Drosophila melanogaster, mammals | [48] |
| Bacterial riboswitches | Variable | Regulate the gene’s activity in response to the concentration of its target molecule | Block translation | 5′-untranslated regions (UTRs) of messenger RNAs | Bacillus subtilis and Neurospora crassa | [49,50] |
| snoRNAs | 60–300 | Guide RNAs in the 2′-O-methylation and pseudouridylation of various classes of RNAs | Involved in nucleolytic processing of ribosomal RNAs to the synthesis of telomeric DNA | U3 species was the first snoRNA described | A broad variety of organisms | [51] |
| tiRNAs | 18 | Modulate local epigenetic structure | Not Found | Sequences immediately downstream of the RNA polymerase II transcription start site | Human, chicken, and Drosophila | [52] |
| moRNAs | 20 | Unknown | Not Found | Located immediately adjacent to miRNAs in the primary miRs | Ciona intestinalis | [53] |
| circRNAs | 1500 | Regulate gene expression in mammals. | Act as endogenous miRNA sponges and buffer microRNA activity | Originated in different ways (“head-to-tail”, “backsplicing” or reverse splicing) | Human, mouse, and C. elegans | [54,55] |
| lincRNAs | 200 | Regulate transcription and epigenetic mechanisms | Regulate chromatin topology by both cis and trans mechanisms, the scaffolding of proteins and other RNAs, act as protein and RNA decoys, regulate neighboring genes, and produce micro-peptides | Exonic sequences and promoter regions | Oryza sativa | [56] |
| T-UCRs | 200 | Unknown | Altered at the transcriptional level in human tumorigenesis; aberrant T-UCR expression profiles may discriminate between different human cancers | Intra- and intergenic regions | Human, mouse, and rat | [37,57] |
| lncRNA | Effect on Resistance 1 | Drug | Cancer Type | Ref. |
|---|---|---|---|---|
| Linc00518 | Absorbs miR-199a, upregulates MRP1, induces chemoresistance | Adriamycin, vincristine, Paclitaxel | BC | [161] |
| EPB41L4A-AS2 | Downregulates ABCB1, increases chemosensitivity | Docetaxel | BC | [162] |
| FOXC2-AS1 | Upregulates ABCB1, induces chemoresistance | Docetaxel | OS | [163] |
| MALAT1 | Upregulates ABC transporters, increases chemoresistance | PTX, DTX, 5-FU, mitomycin | NSCLC, HCC | [158,163,165] |
| CASC9 | Upregulates ABC transporters, increases chemoresistance | Doxorubicin, gemcitabine | BC, PC | [163,173] |
| MRUL | Upregulates ABC transporters, increases chemoresistance | Adriamycin, vincristine | GC | [163] |
| HOTAIR | Downregulates JAK2/STAT3 pathway, increases chemoresistance | Dexamethasone, gemcitabine | MM, PC | [164,177] |
| CCAT2 | Enhances proliferation, increases chemoresistance | Tamoxifen, cisplatin | BC, OC | [166,180] |
| NNT-AS1 | Targets MAPK/Slug pathway, increases chemoresistance | Cisplatin | NSCLC | [167] |
| PVT1 | Modulates Bcl-2 expression, regulates miR-216b and Beclin-1, decreases autophagy and apoptosis, increases chemoresistance | 5-FU, cisplatin, gemcitabine | GC, NSCLC, PC | [168,169,178] |
| AB209630 | Targets EphB2 and Nanog, reduces chemoresistance | Gemcitabine | PC | [171,172] |
| GAS5 | Suppresses cell growth and metastasis, increases chemosensitivity | Gemcitabine | PC | [174] |
| MEG3 | Suppresses cell growth and metastasis, increases chemosensitivity | Gemcitabine | PC | [175] |
| HOTTIP | Downregulates miR-137, increases chemoresistance | Cisplatin | PC | [176] |
| ACTA2-AS1 | Modulates of Bax, survivin, and Bcl-1, increases chemoresistance | Cisplatin | OC | [179] |
| ClinicalTrials.Gov Identifier | Therapeutic Agent | Target Diseases | Combined Drugs/Therapy | Status |
|---|---|---|---|---|
| NCT04406831 | Serum miRNAs | Pancreatic cancer | - | Recruiting |
| NCT04305366 | miRNA signatures | Squamous cell carcinoma of head and neck | - | Recruiting |
| NCT04435756 | miRNA 371 | Germ cell tumors | - | Recruiting |
| NCT03738319 | miRNA signatures | High-grade serous ovarian cancer | - | Unknown |
| NCT03779022 | miRNA biomarkers | Breast cancer | - | Unknown |
| NCT04972201 | Liquid biopsy miRNA | Multi-cancer | - | Recruiting |
| NCT05495685 | Blood miRNAs | Pancreatic cancer | - | Recruiting |
| NCT04427475 | Plasma miRNAs | Advanced non-small-cell lung cancer | Pabolizumab, nafulizumab | Unknown |
| NCT05556603 | Blood miRNAs | Pancreatic cancer | - | Active, not recruiting |
| NCT04903665 | Blood miRNAs | Gynecologic cancer | - | Active, not recruiting |
| NCT05224596 | Blood miRNAs | Gastric cancer | - | Recruiting |
| NCT03742856 | miRNA signatures | Epithelial ovarian cancer | - | Unknown |
| NCT05431621 | miRNA7 | Esophageal cancer, gastric cancer, colorectal cancer, hepatocellular carcinoma | - | Recruiting |
| NCT03742869 | miRNA signatures | Uterine cervical adenocarcinoma | - | Unknown |
| NCT04010487 | miRNA expression profiles | Endometrial carcinoma | - | Unknown |
| NCT04792437 | miRNA expression profiles | Glioma | - | Recruiting |
| NCT05417048 | Exosomal miRNA profiling | Breast cancer | - | Not yet recruiting |
| NCT03397355 | miRNA expression profiles | Lung cancer | - | Unknown |
| NCT01210495 | Blood miRNAs | Advanced hepatocellular carcinoma | Axitinib | Completed |
| NCT03236649 | miRNA biomarkers | Advanced hepatocellular carcinoma | Icaritin, sorafenib tosylate | Unknown |
| NCT03236636 | miRNA biomarkers | Advanced hepatocellular carcinoma | Icaritin, HUACHANSU PIAN | Unknown |
| NCT03108677 | Exosomal miRNA profiling | Primary high-grade osteosarcoma | - | Active, not recruiting |
| NCT03741829 | miRNA expression profiles | Small-cell lung cancer | - | Completed |
| NCT03694483 | Blood miRNAs | Prostate cancer | - | Suspended |
| NCT03526835 | miRNA biomarkers | Metastatic colorectal cancer | MCLA-158 | Unknown |
| NCT02509052 | miRNA biomarkers | Recurrent plasma cell myeloma, refractory plasma cell myeloma | Leflunomide | Active, not recruiting |
| NCT04720430 | miRNA signatures | Hepatocellular carcinoma | - | Recruiting |
| NCT03886571 | Exosomal miRNA profiling | Pancreatic cancer | - | Recruiting |
| NCT00926640 | miRNA profiling | Small-cell lung carcinoma, malignant epithelial neoplasms | Belinostat, cisplatin, etoposide | Completed |
| NCT04515004 | miR-19a | Early-stage lung cancer | Leucoselect phytosome | Not yet recruiting |
| NCT03451383 | Blood or saliva miRNAs | Breast cancer | - | Recruiting |
| NCT02724202 | miRNA profiling | Colon cancer | Curcumin, 5-FU | Unknown |
| NCT03770468 | miRNA profiling | Glioblastoma | - | Active, not recruiting |
| NCT04453046 | miRNA profiling | Squamous cell carcinoma of the head and neck | Pembrolizumab | Terminated |
| NCT04158635 | miRNA profiling | Pancreatic cancer | Bosentan, gemcitabine, Nab-paclitaxel | Recruiting |
| NCT02657005 | miRNA profiling | Ewing sarcoma | TK216 | Terminated |
| NCT01132586 | miR-181 | Acute myeloid leukemia | Lenalidomide, cytarabine, idarubicin | Completed |
| NCT03824327 | Hypoxia-inducible miRNAs | Non-small-cell lung carcinoma | Papaverine hydrochloride | Recruiting |
| NCT01999972 | Circulating miRNAs | Advanced solid tumors | Axitinib, crizotinib | Completed |
| NCT05275075 | miRNA expression | Resectable pancreatic adenocarcinoma | - | Recruiting |
| NCT02507765 | miRNA expression | Liver cancer | - | Completed |
| NCT03824145 | miRNA signatures | Breast cancer | - | Recruiting |
| NCT02642965 | miR-29b, miR-499 | Acute myeloid leukemia | Cytarabine, fludarabine phosphate | Active, not recruiting |
| NCT03953443 | miRNA expression | Head and neck squamous cell carcinoma | - | Active, not recruiting |
| NCT03443908 | miRNA expression | Lung cancer | - | Withdrawn |
| NCT01780662 | miRNA expression | Hodgkin lymphoma | Brentuximab vedotin, gemcitabine hydrochloride | Completed |
| NCT03233724 | miRNA expression profiles | Non-small-cell lung cancers, Esophageal carcinomas | Decitabine, tetrahydrouridine, pembrolizumab | Recruiting |
| NCT01050296 | miRNA expression | Pediatric solid tumors | - | Recruiting |
| NCT00681512 | Serum miRNA profiles | Non-small-cell lung cancer | Berry Powder | Terminated |
| NCT02594202 | miRNA sequencing | Prostate cancer | - | Recruiting |
| NCT01676805 | miRNA sequencing | Lymph cancer | - | Recruiting |
| NCT02983279 | miR-21 | Breast carcinoma, endometrial carcinoma, prostate carcinosarcoma | - | Completed |
| NCT04100811 | miRNA profiling | Prostate cancer | - | Recruiting |
| NCT04697576 | Circulating miRNAs | Stage I, II, and IV melanoma | Ipilimumab, nivolumab, pembrolizumab | Recruiting |
| NCT01555268 | miRNA expression | Acute myeloid leukemia | Trebananib, cytarabine | Completed |
| NCT05136846 | miRNA biomarkers | Stage II-III non-small-cell lung cancer | Carboplatin, durvalumab, paclitaxel | Recruiting |
| NCT00898092 | miRNA expression | Acute myeloid leukemia | - | Active, not recruiting |
| NCT01629498 | Blood miRNAs | Stage II-IIIB non-small-cell lung cancer | - | Recruiting |
| NCT01446809 | Plasma miRNAs | Soft tissue sarcoma | Doxorubicin hydrochloride, ifosfamide | Completed |
| NCT01583283 | microRNA expression profiles | Multiple myeloma | ACY-1215, lenalidomide, dexamethasone | Completed |
| NCT02323607 | miRNA expression | Acute myeloid leukemia | Pacritinib, cytarabine, daunorubicin hydrochloride | Completed |
| NCT03537599 | Exosomal miRNAs | Acute myeloid leukemia | Daratumumab | Completed |
| NCT01955499 | Serum miRNAs | B-cell non-Hodgkin lymphoma | Ibrutinib, lenalidomide | Active, not recruiting |
| NCT01598285 | miRNA signatures | Breast cancer | Bevacizumab | Terminated |
| NCT01612871 | miRNA signatures | Breast cancer | Tamoxifen or anti-aromatase | Completed |
| NCT02635087 | miR-21, miR-20a-5p, miR-103a-3p, miR-106b-5p, miR-143-5p and miR-215 | Colon cancer | - | Recruiting |
| NCT02247453 | 24 previously identified miRNAs | Lung cancer | - | Active, not recruiting |
| NCT02812680 | Circulating microRNAs | Esophageal adenocarcinoma | Multiregimen chemotherapy | Active, not recruiting |
| NCT02466113 | miR-21, miR-20a-5p, miR-103a-3p, miR-106b-5p, miR-143-5p and miR-215 | Colon cancer | Adjuvant chemotherapy | Not yet recruiting |
| NCT01722851 | Circulating microRNAs | Breast cancer | Adjuvant chemotherapy | Completed |
| NCT02656589 | Plasma miRNAs | Breast cancer | Capecitabine and trastuzumab | Unknown |
| NCT01231386 | miRNA signatures | Breast cancer | Neoadjuvant or adjuvant treatment | Completed |
| NCT03452514 | miRNA signatures | Lung cancer | - | Completed |
| NCT02253251 | KRAS-variant and microRNA binding site mutation testing | Breast cancer | - | Recruiting |
| NCT03293433 | miRNA signatures | Pulmonary cancer | - | Completed |
| NCT01964508 | miRNA signatures | Thyroid cancer | - | Completed |
| NCT02009852 | miR-29b | Oral squamous cell carcinoma | - | Unknown |
| NCT00806650 | Serum miRNA signatures | Kidney cancer | - | Completed |
| NCT01220427 | miRNA expression profiles | Prostate cancer | - | Terminated |
| NCT02366494 | Exosomal miRNA profiling | Prostate cancer | Bicalutamide, leuprolide, goserelin, triptorelin, docetaxel | Active, not recruiting |
| NCT00849914 | miRNA signatures | Epithelial skin cancer | - | Completed |
| NCT03432624 | MicroRNA-25 | Pancreatic cancer | - | Unknown |
| NCT02445924 | miRNA signatures | Non-small-cell lung cancer | - | Completed |
| NCT02065908 | Circulating microRNAs | Breast cancer | - | Completed |
| NCT02950207 | miRNA-100 | Breast cancer | - | Unknown |
| NCT02127073 | miRNA signatures | Breast cancer | Intranasal oxytocin | Suspended |
| NCT03051191 | Expression pattern of miRNA in blood | Cancer and cardiovascular diseases | - | Completed |
| NCT01143311 | miRNA expression profiles | Cutaneous squamous cell carcinoma | - | Terminated |
| NCT03309722 | miRNA expression profiles | Colorectal cancer | - | Recruiting |
| NCT01712958 | miRNA expression profiles | Colorectal carcinoma | - | Unknown |
| NCT02471469 | miRNA expression profiles | Metastatic castration-resistant prostate cancer | Enzalutamide | Completed |
| NCT01829971 | MRX34 | Primary liver cancer, Sclc, lymphoma, melanoma multiple myeloma, renal cell carcinoma, Nsclc | - | Terminated |
| NCT02758652 | miRNA expression profiles | Ovarian cancer | - | Active, not recruiting |
| NCT01541800 | Circulating microRNAs | Pediatric cancers: leukemia, lymphoma, central nervous system | - | Unknown |
| NCT02464930 | miR-192-5p, miR-215-5p miR-194-5p | Barrett’s esophagus, gastroesophageal reflux, esophageal adenocarcinoma | - | Unknown |
| NCT02964351 | Circulating microRNAs | Prostate carcinosarcoma | - | Unknown |
| NCT03338712 | miRNA expression profiles | Prostate cancer, radical prostatectomy | - | Withdrawn |
| NCT02791217 | miRNA expression profiles | Lymphoma, B-cell, follicular lymphoma, Hodgkin lymphoma, multiple myeloma | - | Unknown |
| NCT03074175 | Plasma miRNA profiles | Advanced non-small-cell lung cancer | Radiotherapy | Unknown |
| NCT02531607 | Blood miRNAs | Pancreatic neoplasms | - | Active, not recruiting |
| NCT01240369 | miR-326 | Non-small-cell lung cancer, esophagus squamous cell carcinoma | - | Unknown |
| NCT01119573 | miRNA profiling in tissues | Endometrial cancer | - | Unknown |
| NCT02634502 | miRNA expression profiles in serum | Pancreatic cancer with liver metastasis | Drug: S-1 | Unknown |
| NCT02268734 | Circulating miRNAs | Metastatic sporadic medullary thyroid cancer | Vandetanib | Completed |
| NCT01595139 | miRNA expression profiles | Glioma, neurofibromatosis Type 1 | - | Completed |
| NCT01572467 | miRNA expression profiles | Ovarian or testicular sex cord stromal tumors | - | Completed |
| NCT01391351 | miRNA expression profiles | Ovarian carcinoma, fallopian tube cancer, peritoneal, serous-type advanced stage | Taxol and carboplatin therapy and Taxol and carboplatin chemotherapy with avastin | Completed |
| NCT02364154 | Circulating miRNAs | Colorectal cancer | - | Completed |
| NCT01849952 | MicroRNA-10b | Gliomas | - | Recruiting |
| NCT01927354 | MicroRNA-29 family | Head and neck squamous cell carcinoma | - | Unknown |
| NCT02402036 | Serum miRNA profiles | Colorectal cancer | Regorafenib | Terminated |
| NCT03048266 | Serum miRNA profiles | Multiple endocrine neoplasia Type 1 | - | Recruiting |
| NCT00909350 | miRNA expression profiles | Barrett’s esophagus, esophageal adenocarcinoma | - | Completed |
| NCT01595126 | Saliva and serum miRNA profiles | Head and neck cancer | Dietary Supplement | Unknown |
| NCT01828918 | miRNA expression profiles | Colorectal cancer | - | Unknown |
| NCT01965522 | Serum miRNA profiles | Breast cancer | Vitamin D and melatonin | Completed |
| NCT01498250 | miRNA expression profiles | Basal cell carcinoma | - | Completed |
| NCT01500954 | miRNA expression profiles | Cutaneous squamous cell carcinoma | - | Completed |
| NCT01453465 | miRNA expression profiles | Brain and central nervous system tumors, kidney cancer | - | Withdrawn |
| NCT03253107 | miRNA expression profiles | Gastric cancer | XP (xeloda + cisplatin) or Xelox (xeloda + oxaliplatin) +/− Herceptin | Recruiting |
| NCT03202810 | Serum miRNA profiles | Oral cancer | - | Unknown |
| NCT03081988 | Circulating miRNAs | Esophageal cancer | Concomitant chemoradiotherapy | Recruiting |
| NCT01556178 | miRNA expression profiles | Hydrocephalus | - | Completed |
| NCT02807896 | miRNA expression profiles | Pancreatic cancer, bile duct cancer, stomach cancer, colon cancer | - | Completed |
| NCT03429530 | Circulating miRNAs | Hepatocellular carcinoma | - | Completed |
| NCT00581750 | miRNA expression profiles | Breast cancer, lobular carcinoma, invasive breast cancer | - | Completed |
| NCT01970696 | miRNA expression profiles | Ovarian stromal tumor, testicular stromal tumors, ovarian small-cell carcinoma, Dicer1 Syndrome | - | Recruiting |
| NCT03227510 | Circulating miRNAs | Hepatocellular carcinoma | - | Unknown |
| NCT02928627 | Hepatic and circulating miR-221 and miR-222 | Hepatocellular carcinoma | - | Unknown |
| NCT03255486 | miRNA expression profiles | Advanced breast cancer | - | Completed |
| NCT01229124 | miRNA expression profiles | Acute myeloid leukemia | - | Completed |
| NCT03362684 | miR-31-5p and miR-31-3p | Colorectal cancer | Cetuximab and FOLFOX | Completed |
| NCT03167476 | miRNA expression profiles | Lymphoma, reactive hyperplasia lymphoid | - | Unknown |
| NCT02507882 | miRNA expression profiles | Hepatocellular carcinoma | - | Unknown |
| NCT01957332 | miRNA expression profiles | Metastatic breast cancer | - | Active, not recruiting |
| NCT00897234 | miRNA expression profiles | Lung cancer | - | Completed |
| NCT01528956 | miRNA expression profiles | Pediatric adrenocortical tumors | - | Completed |
| NCT02862145 | MRX34 | Melanoma | Dexamethasone | Withdrawn |
| NCT01345760 | miRNA expression profiles | Basal cell carcinoma, squamous cell carcinoma | - | Completed |
| NCT00743054 | miRNA expression profiles | Renal cell carcinoma | - | Completed |
| NCT01444560 | miRNA expression profiles | Cutaneous melanoma | - | Completed |
| NCT01298414 | AML-specific miRs (miR-34a, miR-538e, miR-193e, and miR-198) | Acute myeloid leukemia | - | Completed |
| NCT01057199 | microRNA-34a and microRNA-194 | Acute myeloid leukemia | - | Completed |
| NCT02392377 | miRNA expression profiles | Esophageal adenocarcinoma | Paclitaxel, carboplatin, oxaliplatin, leucovorin, calcium, fluorouracil | Terminated |
| NCT02412579 | miRNA expression profiles | Hepatocellular carcinoma | - | Completed |
| NCT02369198 | miR-16 family | Malignant pleural mesothelioma, non-small-cell lung cancer | TargomiR | Completed |
| NCT01433809 | miRNA expression profiles | Thyroid cancer | - | Completed |
| NCT02448056 | miRNA expression profiles | Hepatocellular carcinoma | Sorafenib | Not yet recruiting |
| NCT03142633 | miRNA expression profiles | Polycystic ovary syndrome | - | Completed |
| NCT01482260 | miRNA expression profiles | Cutaneous malignant melanoma | - | Completed |
| NCT03000335 | miR-451, miR-151-5p and miR-1290 | Acute lymphoblastic leukemia | - | Unknown |
| NCT00536029 | miRNA expression profiles | Melanoma | - | Completed |
| NCT00862914 | miRNA expression profiles | Malignant melanoma | - | Completed |
| NCT03474614 | miRNA expression profiles | Cerebral cavernous malformations | Propranolol | Unknown |
| NCT01247506 | miRNA expression profiles | Hepatocellular carcinoma | - | Unknown |
| NCT01505699 | miRNA expression profiles | B-cell acute lymphoblastic leukemia | - | Completed |
| NCT01511575 | miRNA expression profiles | Down syndrome acute myeloid leukemia | - | Completed |
| NCT01606605 | miRNA expression profiles | Diffuse large B-cell lymphoma | Completed | |
| NCT00639054 | miRNA expression profiles | Multiple myeloma | - | Completed |
| NCT03416803 | miRNA expression profiles | Hepatocellular carcinoma | Radiotherapy | Unknown |
| NCT01560195 | miRNA expression profiles | Advanced non-small-cell lung cancer | Pegylated rhG-CSF | Unknown |
| NCT02580552 | miR-155 | Cutaneous T-cell lymphoma, mycosis fungoides, chronic lymphocytic leukemia, diffuse large B-cell lymphoma, ABC subtype adult T-cell leukemia/lymphoma | Cobomarsen (MRG-106) | Completed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayraktar, E.; Bayraktar, R.; Oztatlici, H.; Lopez-Berestein, G.; Amero, P.; Rodriguez-Aguayo, C. Targeting miRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update. Non-Coding RNA 2023, 9, 27. https://doi.org/10.3390/ncrna9020027
Bayraktar E, Bayraktar R, Oztatlici H, Lopez-Berestein G, Amero P, Rodriguez-Aguayo C. Targeting miRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update. Non-Coding RNA. 2023; 9(2):27. https://doi.org/10.3390/ncrna9020027
Chicago/Turabian StyleBayraktar, Emine, Recep Bayraktar, Hulya Oztatlici, Gabriel Lopez-Berestein, Paola Amero, and Cristian Rodriguez-Aguayo. 2023. "Targeting miRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update" Non-Coding RNA 9, no. 2: 27. https://doi.org/10.3390/ncrna9020027