
Pro-Inflammatory Cytokines Promote the Transcription of Circular RNAs in Human Pancreatic β Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell and Islet Culture
2.2. circRNA Microarray Labeling and Hybridization
2.3. Differential Expression Analysis
2.4. Real-Time qPCR Validation of circRNAs, Host Genes, miRNA Targets, and miRNAs
2.5. RNA Sequencing and Proteomics Data in EndoC-βH1 Cells
2.6. Prediction of circRNA–miRNA and circRNA–RNA-Binding Protein Interactions
2.7. Pathway Analysis
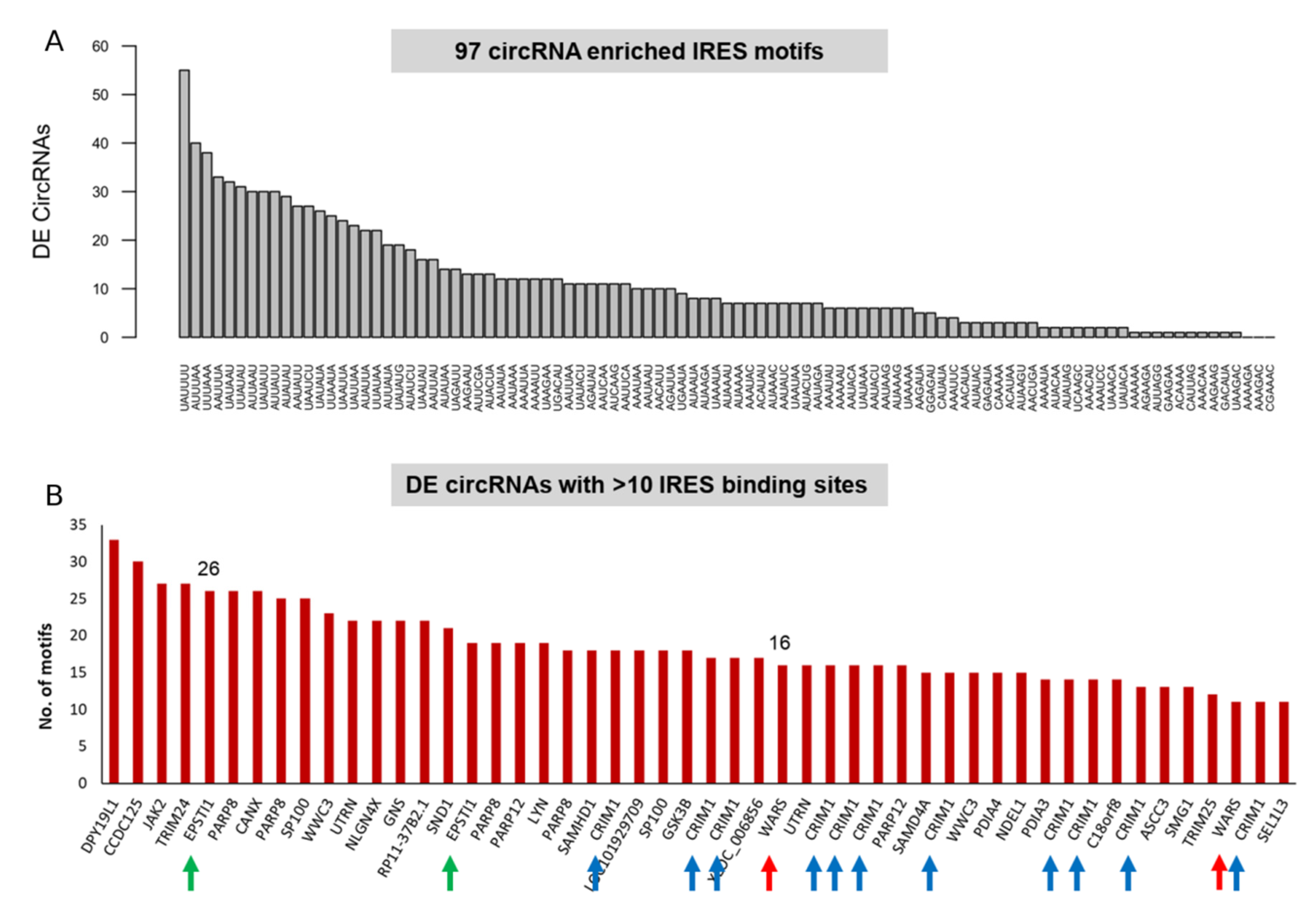
2.8. circRNAs Enriched in IRES and IRE Sites
2.9. Evolutionarily Conserved circRNAs
3. Results
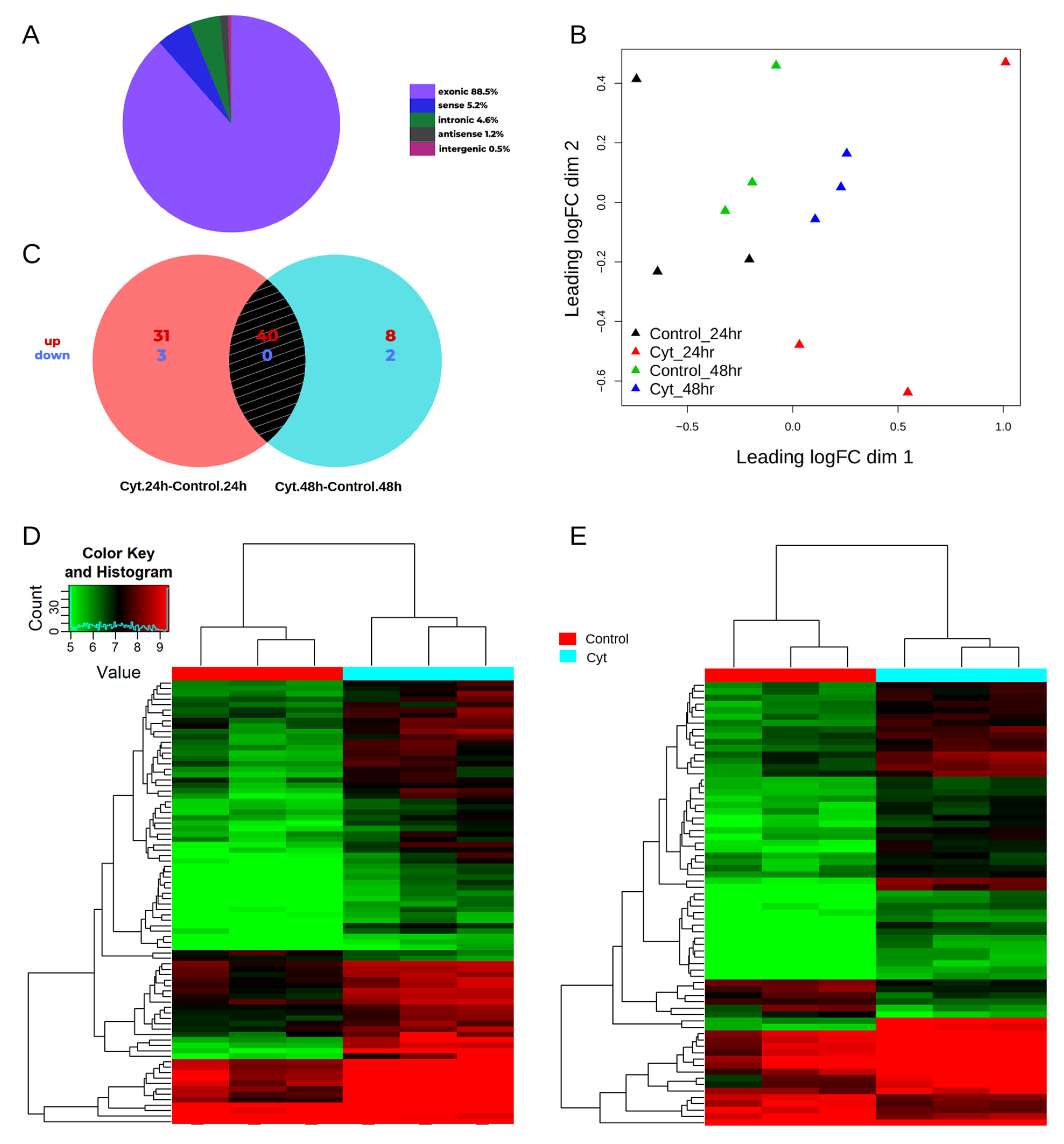
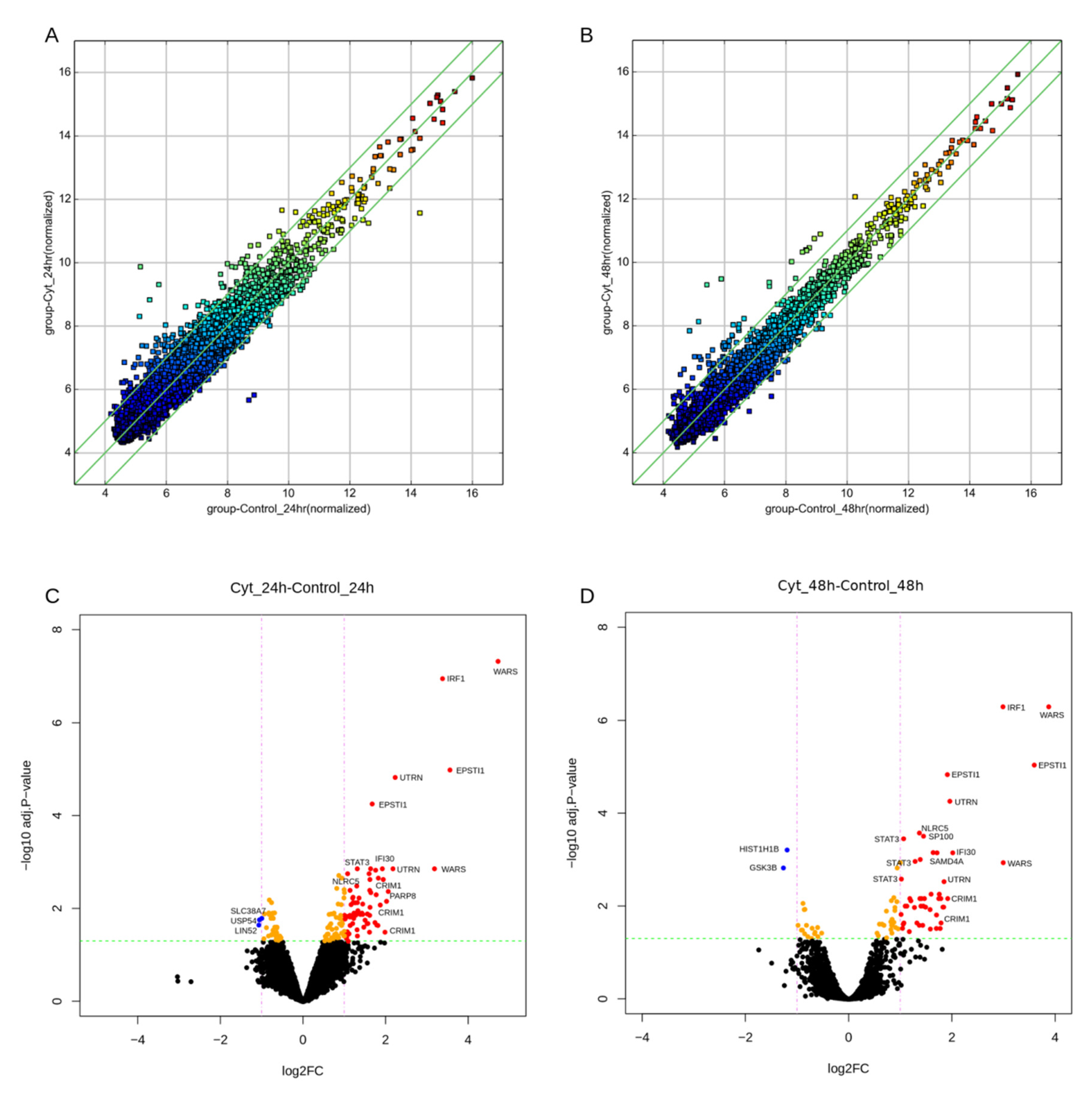
3.1. circRNAs Are Differentially Expressed in the EndoC-βH1 β-Cell Line
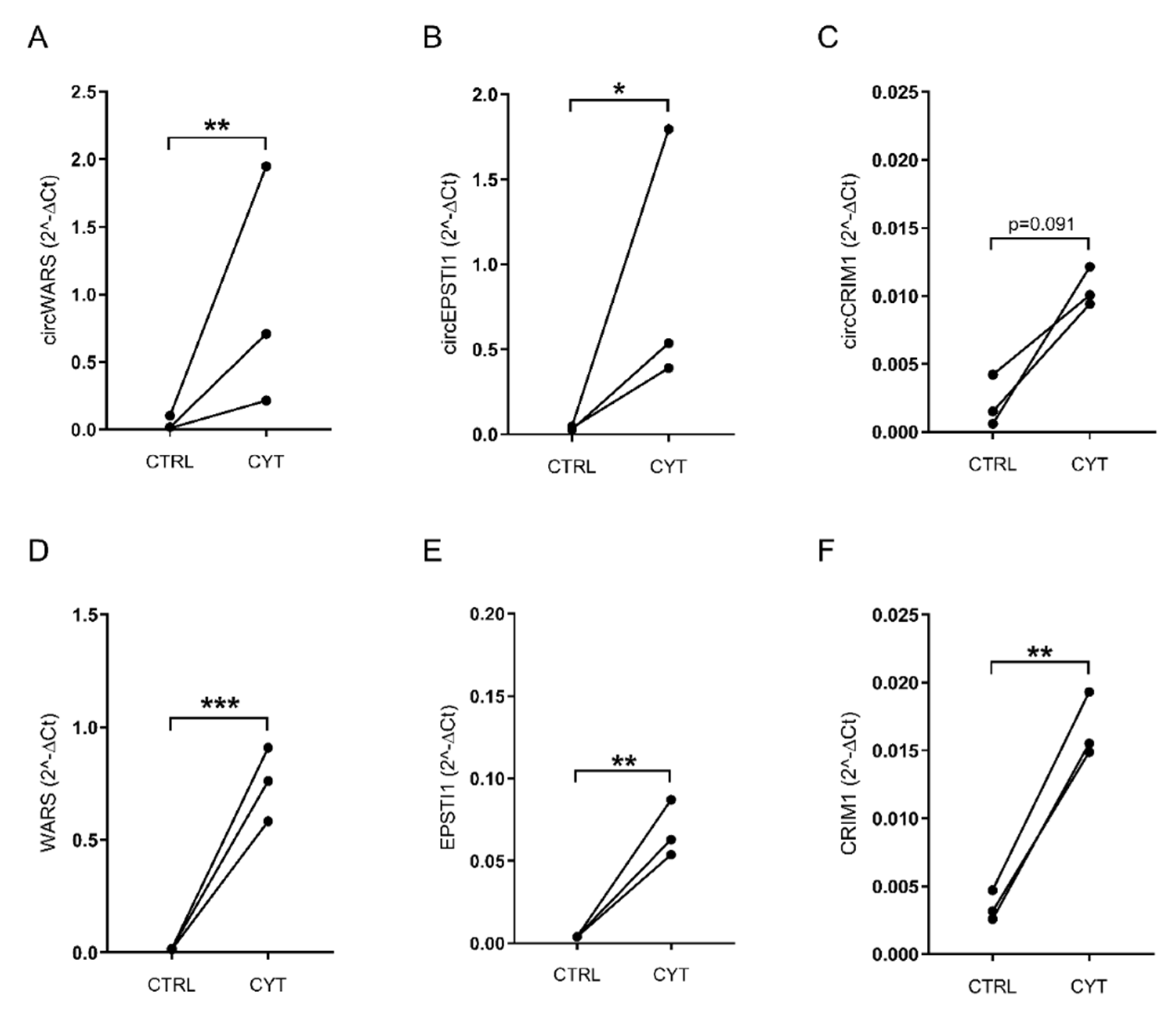
3.2. Validation of circWARS, circEPSTI1, and circCRIM1 in EndoC-βH1 Cells and Human Pancreatic Islets
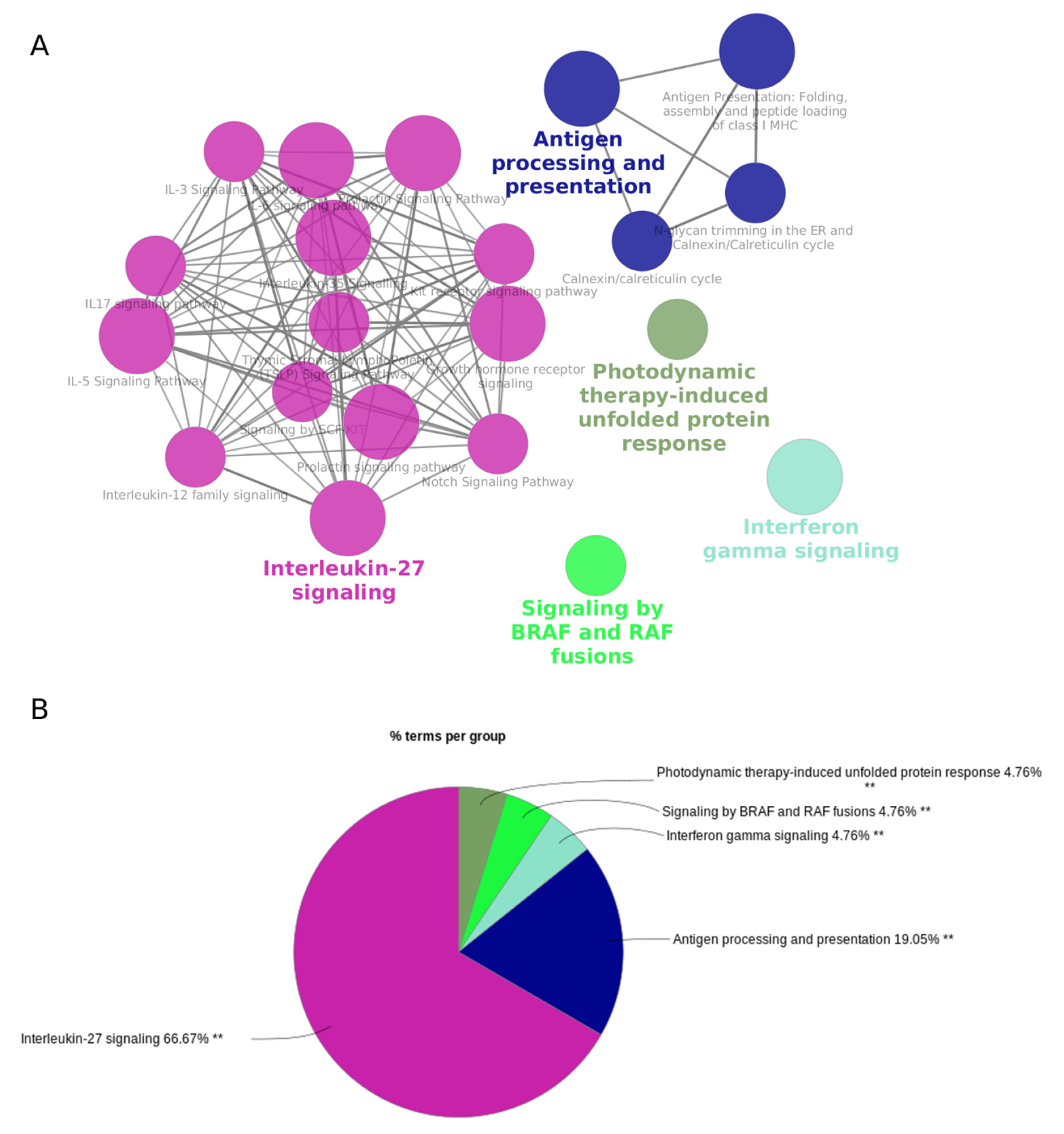
3.3. Host Genes of the Cytokine-Regulated circRNAs Are Enriched in Interferon and Interleukin Signaling Pathways
3.4. circRNA Translation Is Driven by IRES-Like Short Elements
3.5. Multi-Species Conserved Sequences (MCS) Analysis
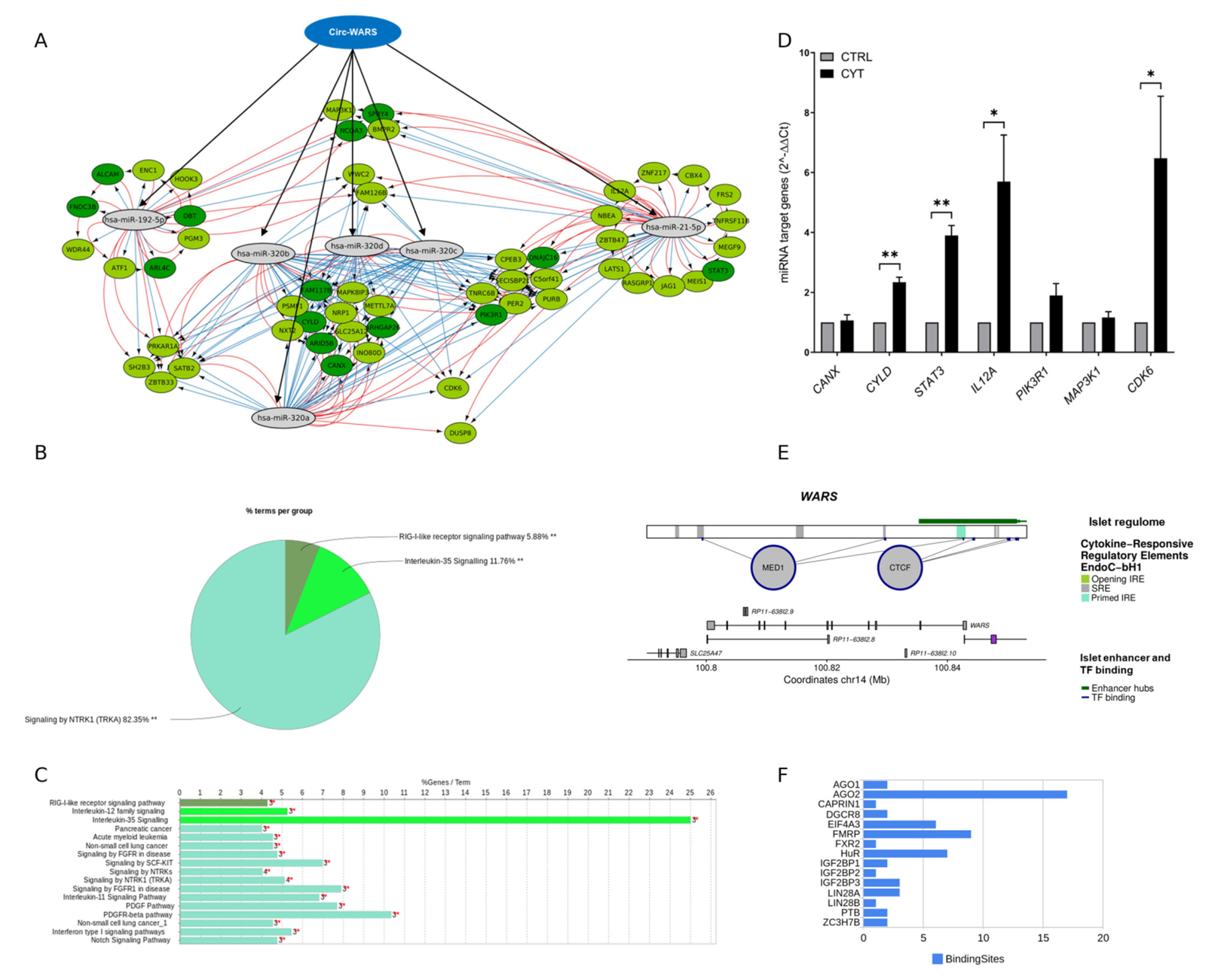
3.6. Cytokine-Induced Regulatory Elements in circRNAs
3.7. RNA-Binding Protein (RBP) and miRNA Binding Sites within circWARS, circEPSTI1, and circCRIM1
3.8. circWARS–miRNA–mRNA Interaction Network in EndoC-βH1 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortis, F.; Naamane, N.; Flamez, D.; Ladrière, L.; Moore, F.; Cunha, D.A.; Colli, M.L.; Thykjaer, T.; Thorsen, K.; Orntoft, T.F.; et al. Cytokines Interleukin-1beta and Tumor Necrosis Factor-Alpha Regulate Different Transcriptional and Alternative Splicing Networks in Primary Beta-Cells. Diabetes 2010, 59, 358–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berchtold, L.A.; Prause, M.; Størling, J.; Mandrup-Poulsen, T. Cytokines and Pancreatic β-Cell Apoptosis. Adv. Clin. Chem. 2016, 75, 99–158. [Google Scholar] [CrossRef] [PubMed]
- Eizirik, D.L.; Pasquali, L.; Cnop, M. Pancreatic β-Cells in Type 1 and Type 2 Diabetes Mellitus: Different Pathways to Failure. Nat. Rev. Endocrinol. 2020, 16, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-Coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in Human Diseases and Potential Use as Biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef]
- Özcan, S. MicroRNAs in Pancreatic β-Cell Physiology. Adv. Exp. Med. Biol. 2015, 887, 101–117. [Google Scholar] [CrossRef]
- Eliasson, L. The Small RNA MiR-375—A Pancreatic Islet Abundant MiRNA with Multiple Roles in Endocrine Beta Cell Function. Mol. Cell Endocrinol. 2017, 456, 95–101. [Google Scholar] [CrossRef]
- Eliasson, L.; Esguerra, J.L.S. Role of Non-Coding RNAs in Pancreatic Beta-Cell Development and Physiology. Acta Physiol. 2014, 211, 273–284. [Google Scholar] [CrossRef]
- Mirza, A.H.; Kaur, S.; Pociot, F. Long Non-Coding RNAs as Novel Players in β Cell Function and Type 1 Diabetes. Hum. Genom. 2017, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and Characterizing Circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs Are Abundant, Conserved, and Associated with ALU Repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Barrett, S.P.; Wang, P.L.; Salzman, J. Circular RNA Biogenesis Can Proceed through an Exon-Containing Lariat Precursor. eLife 2015, 4, e07540. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Zang, J.; Lu, D.; Xu, A. The Interaction of CircRNAs and RNA Binding Proteins: An Important Part of CircRNA Maintenance and Function. J. Neurosci. Res. 2020, 98, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Kulcheski, F.R.; Christoff, A.P.; Margis, R. Circular RNAs Are MiRNA Sponges and Can Be Used as a New Class of Biomarker. J. Biotechnol. 2016, 238, 42–51. [Google Scholar] [CrossRef]
- Stoll, L.; Sobel, J.; Rodriguez-Trejo, A.; Guay, C.; Lee, K.; Venø, M.T.; Kjems, J.; Laybutt, D.R.; Regazzi, R. Circular RNAs as Novel Regulators of β-Cell Functions in Normal and Disease Conditions. Mol. Metab. 2018, 9, 69–83. [Google Scholar] [CrossRef]
- Zaiou, M. CircRNAs Signature as Potential Diagnostic and Prognostic Biomarker for Diabetes Mellitus and Related Cardiovascular Complications. Cells 2020, 9, 659. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, Y.; Zhao, M.; Zou, J.; Zhu, Y.; Yuan, X.; Liu, Q.; Cai, H.; Chu, C.-Q.; Liu, Y. Differential Profile of Plasma Circular RNAs in Type 1 Diabetes Mellitus. Diabetes Metab. J. 2020, 44, 854–865. [Google Scholar] [CrossRef]
- Luo, S.; Deng, M.; Xie, Z.; Li, X.; Huang, G.; Zhou, Z. Circulating Circular RNAs Profiles Associated with Type 1 Diabetes. Diabetes Metab. Res. Rev. 2021, 37, e3394. [Google Scholar] [CrossRef]
- Yang, L.; Han, X.; Zhang, C.; Sun, C.; Huang, S.; Xiao, W.; Gao, Y.; Liang, Q.; Luo, F.; Lu, W.; et al. Hsa_circ_0060450 Negatively Regulates Type I Interferon-Induced Inflammation by Serving as MiR-199a-5p Sponge in Type 1 Diabetes Mellitus. Front. Immunol. 2020, 11, 576903. [Google Scholar] [CrossRef]
- Zhang, C.; Han, X.; Yang, L.; Fu, J.; Sun, C.; Huang, S.; Xiao, W.; Gao, Y.; Liang, Q.; Wang, X.; et al. Circular RNA CircPPM1F Modulates M1 Macrophage Activation and Pancreatic Islet Inflammation in Type 1 Diabetes Mellitus. Theranostics 2020, 10, 10908–10924. [Google Scholar] [CrossRef]
- Wang, Z.; Deng, C.; Zheng, Y. Involvement of CircRNAs in Proinflammatory Cytokines-Mediated β-Cell Dysfunction. Mediat. Inflamm. 2021, 2021, 5566453. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Mirza, A.H.; Pociot, F. Cell Type-Selective Expression of Circular RNAs in Human Pancreatic Islets. Noncoding RNA 2018, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, S.; Ames, R.M.; Moore, K.; Lee, B.P.; Jeffery, N.; Harries, L.W. Islet-expressed circular RNAs are associated with type 2 diabetes status in human primary islets and in peripheral blood. BMC Med. Genom. 2020, 13, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The Circular RNA Cdr1as, via MiR-7 and Its Targets, Regulates Insulin Transcription and Secretion in Islet Cells. Sci. Rep. 2015, 5, 12453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravassard, P.; Hazhouz, Y.; Pechberty, S.; Bricout-Neveu, E.; Armanet, M.; Czernichow, P.; Scharfmann, R. A Genetically Engineered Human Pancreatic β Cell Line Exhibiting Glucose-Inducible Insulin Secretion. J. Clin. Investig. 2011, 121, 3589–3597. [Google Scholar] [CrossRef]
- Brozzi, F.; Nardelli, T.R.; Lopes, M.; Millard, I.; Barthson, J.; Igoillo-Esteve, M.; Grieco, F.A.; Villate, O.; Oliveira, J.M.; Casimir, M.; et al. Cytokines Induce Endoplasmic Reticulum Stress in Human, Rat and Mouse Beta Cells via Different Mechanisms. Diabetologia 2015, 58, 2307–2316. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Rodríguez, M.; Raurell-Vila, H.; Colli, M.L.; Alvelos, M.I.; Subirana-Granés, M.; Juan-Mateu, J.; Norris, R.; Turatsinze, J.-V.; Nakayasu, E.S.; Webb-Robertson, B.-J.M.; et al. The Impact of Proinflammatory Cytokines on the β-Cell Regulatory Landscape Provides Insights into the Genetics of Type 1 Diabetes. Nat. Genet. 2019, 51, 1588–1595. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. CircBase: A Database for Circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-H.; Liu, S.; Zhou, H.; Qu, L.-H.; Yang, J.-H. StarBase v2.0: Decoding MiRNA-CeRNA, MiRNA-NcRNA and Protein-RNA Interaction Networks from Large-Scale CLIP-Seq Data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting Effective MicroRNA Target Sites in Mammalian MRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Chou, C.-H.; Chang, N.-W.; Shrestha, S.; Hsu, S.-D.; Lin, Y.-L.; Lee, W.-H.; Yang, C.-D.; Hong, H.-C.; Wei, T.-Y.; Tu, S.-J.; et al. MiRTarBase 2016: Updates to the Experimentally Validated MiRNA-Target Interactions Database. Nucleic Acids Res. 2016, 44, D239–D247. [Google Scholar] [CrossRef]
- Kutmon, M.; Ehrhart, F.; Willighagen, E.L.; Evelo, C.T.; Coort, S.L. CyTargetLinker App Update: A Flexible Solution for Network Extension in Cytoscape. F1000Research 2018, 7, ELIXIR-743. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- van de Bunt, M.; Gaulton, K.J.; Parts, L.; Moran, I.; Johnson, P.R.; Lindgren, C.M.; Ferrer, J.; Gloyn, A.L.; McCarthy, M.I. The MiRNA Profile of Human Pancreatic Islets and Beta-Cells and Relationship to Type 2 Diabetes Pathogenesis. PLoS ONE 2013, 8, e55272. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A Web Tool for Exploring Circular RNAs and Their Interacting Proteins and MicroRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape Plug-in to Decipher Functionally Grouped Gene Ontology and Pathway Annotation Networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Yang, Y.; Chen, C.; Wang, Z. Pervasive Translation of Circular RNAs Driven by Short IRES-like Elements. Nat. Commun. 2022, 13, 3751. [Google Scholar] [CrossRef]
- Wu, W.; Ji, P.; Zhao, F. CircAtlas: An Integrated Resource of One Million Highly Accurate Circular RNAs from 1070 Vertebrate Transcriptomes. Genome Biol. 2020, 21, 101. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Caudron-Herger, M.; Diederichs, S. Circ2GO: A Database Linking Circular RNAs to Gene Function. Cancers 2020, 12, 2975. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Matsumoto, K.; Nishihara, M.; Nakano, Y.; Shibata, A.; Maruyama, H.; Shuto, S.; Matsuda, A.; Yoshida, M.; Ito, Y.; et al. Rolling Circle Translation of Circular RNA in Living Human Cells. Sci. Rep. 2015, 5, 16435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Han, P.; Zhou, T.; Guo, X.; Song, X.; Li, Y. CircRNADb: A Comprehensive Database for Human Circular RNAs with Protein-Coding Annotations. Sci. Rep. 2016, 6, 34985. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA That Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Klein, D.; Misawa, R.; Bravo-Egana, V.; Vargas, N.; Rosero, S.; Piroso, J.; Ichii, H.; Umland, O.; Zhijie, J.; Tsinoremas, N.; et al. MicroRNA Expression in Alpha and Beta Cells of Human Pancreatic Islets. PLoS ONE 2013, 8, e55064. [Google Scholar] [CrossRef] [Green Version]
- Brozzi, F.; Regazzi, R. Circular RNAs as Novel Regulators of β-Cell Functions under Physiological and Pathological Conditions. Int. J. Mol. Sci. 2021, 22, 1503. [Google Scholar] [CrossRef]
- Stoll, L.; Rodríguez-Trejo, A.; Guay, C.; Brozzi, F.; Bayazit, M.B.; Gattesco, S.; Menoud, V.; Sobel, J.; Marques, A.C.; Venø, M.T.; et al. A Circular RNA Generated from an Intron of the Insulin Gene Controls Insulin Secretion. Nat. Commun. 2020, 11, 5611. [Google Scholar] [CrossRef]
- Das, D.; Das, A.; Sahu, M.; Mishra, S.S.; Khan, S.; Bejugam, P.R.; Rout, P.K.; Das, A.; Bano, S.; Mishra, G.P.; et al. Identification and Characterization of Circular Intronic RNAs Derived from Insulin Gene. Int. J. Mol. Sci. 2020, 21, 4302. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z. Efficient Backsplicing Produces Translatable Circular MRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.-L.; Wang, Y.; et al. Extensive Translation of Circular RNAs Driven by N6-Methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- Garikipati, V.N.S.; Uchida, S. Elucidating the Functions of Non-Coding RNAs from the Perspective of RNA Modifications. Noncoding RNA 2021, 7, 31. [Google Scholar] [CrossRef]
- Jin, M. Unique Roles of Tryptophanyl-TRNA Synthetase in Immune Control and Its Therapeutic Implications. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.T.; Yoon, H.K.; Kim, Y.T.; Choi, Y.H.; Lee, W.-K.; Jin, M. Tryptophanyl-TRNA Synthetase 1 Signals Activate TREM-1 via TLR2 and TLR4. Biomolecules 2020, 10, 1283. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Oh, S.-C.; Zhou, S.; Kim, T.-D. Tryptophanyl-TRNA Synthetase as a Potential Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 4523. [Google Scholar] [CrossRef]
- Du, H.; Yin, Z.; Zhao, Y.; Li, H.; Dai, B.; Fan, J.; He, M.; Nie, X.; Wang, C.-Y.; Wang, D.W.; et al. MiR-320a Induces Pancreatic β Cells Dysfunction in Diabetes by Inhibiting MafF. Mol. Ther. Nucleic Acids 2021, 26, 444–457. [Google Scholar] [CrossRef]
- Du, H.; Zhao, Y.; Yin, Z.; Wang, D.W.; Chen, C. The Role of MiR-320 in Glucose and Lipid Metabolism Disorder-Associated Diseases. Int. J. Biol. Sci. 2021, 17, 402–416. [Google Scholar] [CrossRef]
- He, M.; Wang, J.; Yin, Z.; Zhao, Y.; Hou, H.; Fan, J.; Li, H.; Wen, Z.; Tang, J.; Wang, Y.; et al. MiR-320a Induces Diabetic Nephropathy via Inhibiting MafB. Aging 2019, 11, 3055–3079. [Google Scholar] [CrossRef]
- Li, H.; Fan, J.; Chen, C.; Wang, D.W. Subcellular MicroRNAs in Diabetic Cardiomyopathy. Ann. Transl. Med. 2020, 8, 1602. [Google Scholar] [CrossRef]
- Bravo, R.; Parra, V.; Gatica, D.; Rodriguez, A.E.; Torrealba, N.; Paredes, F.; Wang, Z.V.; Zorzano, A.; Hill, J.A.; Jaimovich, E.; et al. Endoplasmic Reticulum and the Unfolded Protein Response: Dynamics and Metabolic Integration. Int. Rev. Cell Mol. Biol. 2013, 301, 215–290. [Google Scholar] [CrossRef] [Green Version]
- Coe, H.; Bedard, K.; Groenendyk, J.; Jung, J.; Michalak, M. Endoplasmic Reticulum Stress in the Absence of Calnexin. Cell Stress Chaperones 2008, 13, 497–507. [Google Scholar] [CrossRef] [Green Version]
- McCurdy, C.E.; Schenk, S.; Holliday, M.J.; Philp, A.; Houck, J.A.; Patsouris, D.; MacLean, P.S.; Majka, S.M.; Klemm, D.J.; Friedman, J.E. Attenuated Pik3r1 Expression Prevents Insulin Resistance and Adipose Tissue Macrophage Accumulation in Diet-Induced Obese Mice. Diabetes 2012, 61, 2495–2505. [Google Scholar] [CrossRef] [Green Version]
- Rajwani, A.; Ezzat, V.; Smith, J.; Yuldasheva, N.Y.; Duncan, E.R.; Gage, M.; Cubbon, R.M.; Kahn, M.B.; Imrie, H.; Abbas, A.; et al. Increasing Circulating IGFBP1 Levels Improves Insulin Sensitivity, Promotes Nitric Oxide Production, Lowers Blood Pressure, and Protects against Atherosclerosis. Diabetes 2012, 61, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Liu, X.; Zhang, S.; Chen, S.; Guan, X.; Li, Q.; Chen, X.; Zhao, Y. CircCRIM1 Promotes Ovarian Cancer Progression by Working as CeRNAs of CRIM1 and Targeting MiR-383-5p/ZEB2 Axis. Reprod. Biol. Endocrinol. 2021, 19, 176. [Google Scholar] [CrossRef]
- Kolle, G.; Georgas, K.; Holmes, G.P.; Little, M.H.; Yamada, T. CRIM1, a Novel Gene Encoding a Cysteine-Rich Repeat Protein, Is Developmentally Regulated and Implicated in Vertebrate CNS Development and Organogenesis. Mech. Dev. 2000, 90, 181–193. [Google Scholar] [CrossRef]
- Wilkinson, L.; Kolle, G.; Wen, D.; Piper, M.; Scott, J.; Little, M. CRIM1 Regulates the Rate of Processing and Delivery of Bone Morphogenetic Proteins to the Cell Surface. J. Biol. Chem. 2003, 278, 34181–34188. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Wang, H.; Ye, L.; Shi, X.; Yan, K.; Lin, K.; Huang, Q.; Li, B.; Lin, Q.; Zhu, L.; et al. Hsa_circ_0000479 as a Novel Diagnostic Biomarker of Systemic Lupus Erythematosus. Front. Immunol. 2019, 10, 2281. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, H.L.; Rønnov-Jessen, L.; Villadsen, R.; Petersen, O.W. Identification of EPSTI1, a Novel Gene Induced by Epithelial-Stromal Interaction in Human Breast Cancer. Genomics 2002, 79, 703–710. [Google Scholar] [CrossRef]
- Haque, S.; Harries, L.W. Circular RNAs (CircRNAs) in Health and Disease. Genes 2017, 8, 353. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, H.; Lu, Y.; Cheng, L. Regulatory Effects of Circular RNAs on Host Genes in Human Cancer. Front. Oncol. 2021, 10, 586163. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| circRNA | 24 h | 48 h | ||||
|---|---|---|---|---|---|---|
| Gene | circRNA | circBase ID | logFC | adj. p-Value | logFC | adj. p Value |
| WARS | hsa_circRNA_101439 | hsa_circ_0033184 | 4.72 | 4.7 × 10−8 | 3.87 | 5.0 × 10−7 |
| WARS | hsa_circRNA_033191 | hsa_circ_0033191 | 3.18 | 1.4 × 10−3 | 2.99 | 1.1 × 10−3 |
| EPSTI1 | hsa_circRNA_000479 | hsa_circ_0000479 | 3.55 | 1.0 × 10−5 | 3.59 | 9.0 × 10−6 |
| EPSTI1 | hsa_circRNA_405138 | 1.67 | 5.5 × 10−5 | 1.91 | 1.4 × 10−5 | |
| IRF1 | hsa_circRNA_001526 | hsa_circ_0001526 | 3.37 | 1.1 × 10−7 | 2.99 | 5.0 × 10−7 |
| UTRN | hsa_circRNA_001646 | hsa_circ_0001646 | 2.23 | 1.5 × 10−5 | 1.96 | 5.4 × 10−5 |
| UTRN | hsa_circRNA_001648 | hsa_circ_0001648 | 2.17 | 1.4 × 10−3 | 1.84 | 2.9 × 10−3 |
| CRIM1 | hsa_circRNA_102689 | hsa_circ_0054021 | 1.76 | 1.5 × 10−3 | 1.38 | 6.8 × 10−3 |
| CRIM1 | hsa_circRNA_102686 | hsa_circ_0053967 | 1.82 | 2.2 × 10−3 | 1.44 | 9.8 × 10−3 |
| CRIM1 | hsa_circRNA_007408 | hsa_circ_0007408 | 1.62 | 2.3 × 10−3 | 1.27 | 1.0 × 10−2 |
| CRIM1 | hsa_circRNA_102688 | hsa_circ_0006422 | 1.64 | 4.5 × 10−3 | 1.17 | 3.5 × 10−2 |
| CRIM1 | hsa_circRNA_102687 | hsa_circ_0002938 | 2.02 | 6.9 × 10−3 | 1.58 | 3.1 × 10−2 |
| CRIM1 | hsa_circRNA_102681 | hsa_circ_0053958 | 1.87 | 8.2 × 10−3 | 1.84 | 1.0 × 10−2 |
| CRIM1 | hsa_circRNA_005507 | hsa_circ_0005507 | 1.62 | 9.2 × 10−3 | 1.74 | 6.8 × 10−3 |
| CRIM1 | hsa_circRNA_405851 | 1.42 | 1.2 × 10−2 | 1.49 | 1.0 × 10−2 | |
| CRIM1 | hsa_circRNA_102682 | hsa_circ_0005442 | 1.56 | 1.3 × 10−2 | 1.78 | 6.8 × 10−3 |
| CRIM1 | hsa_circRNA_102679 | hsa_circ_0005579 | 1.55 | 1.3 × 10−2 | 1.43 | 2.5 × 10−2 |
| CRIM1 | hsa_circRNA_102677 | hsa_circ_0002017 | 1.44 | 1.3 × 10−2 | 1.75 | 5.4 × 10−3 |
| CRIM1 | hsa_circRNA_053955 | hsa_circ_0053955 | 1.61 | 1.4 × 10−2 | 1.91 | 6.8 × 10−3 |
| CRIM1 | hsa_circRNA_102680 | hsa_circ_0002348 | 1.74 | 2.0 × 10−2 | 1.79 | 2.3 × 10−2 |
| CRIM1 | hsa_circRNA_102678 | hsa_circ_0002346 | 1.57 | 2.1 × 10−2 | 1.82 | 1.0 × 10−2 |
| CRIM1 | hsa_circRNA_004182 | hsa_circ_0004182 | 1.55 | 2.1 × 10−2 | 1.7 | 1.5 × 10−2 |
| CRIM1 | hsa_circRNA_006294 | hsa_circ_0006294 | 1.78 | 2.2 × 10−2 | 1.77 | 3.0 × 10−2 |
| CRIM1 | hsa_circRNA_102685 | hsa_circ_0003578 | 1.61 | 3.2 × 10−2 | 1.69 | 3.0 × 10−2 |
| IFI30 | hsa_circRNA_102484 | hsa_circ_0005571 | 1.92 | 1.4 × 10−3 | 2.01 | 7.0 × 10−4 |
| STAT3 | hsa_circRNA_401803 | 1.77 | 4.9 × 10−3 | 1.36 | 2.5 × 10−2 | |
| STAT3 | hsa_circRNA_102073 | hsa_circ_0043812 | 1.31 | 1.4 × 10−3 | 1.28 | 1.1 × 10−3 |
| GRM4 | hsa_circRNA_104092 | hsa_circ_0076041 | 1.44 | 7.9 × 10−3 | 1.59 | 5.4 × 10−3 |
| GRM4 | hsa_circRNA_104091 | hsa_circ_0076040 | 1.31 | 2.8 × 10−2 | 1.57 | 1.2 × 10−2 |
| GRM4 | hsa_circRNA_076039 | hsa_circ_0076039 | 1.32 | 1.1 × 10−2 | 1.46 | 6.8 × 10−3 |
| SAMD4A | hsa_circRNA_101356 | hsa_circ_0004846 | 1.63 | 1.4 × 10−3 | 1.71 | 7.0 × 10−4 |
| SAMHD1 | hsa_circRNA_406106 | 1.6 | 1.7 × 10−3 | 1.07 | 2.3 × 10−2 | |
| LOC101928767 | hsa_circRNA_405273 | 1.61 | 4.0 × 10−3 | 1.4 | 9.8 × 10−3 | |
| LOC101929709 | hsa_circRNA_002576 | hsa_circ_0002576 | 1.21 | 5.6 × 10−3 | 1.18 | 6.8 × 10−3 |
| PARP12 | hsa_circRNA_082689 | hsa_circ_0082689 | 1.39 | 1.3 × 10−2 | 1.32 | 2.3 × 10−2 |
| PARP8 | hsa_circRNA_103835 | hsa_circ_0072431 | 1.35 | 5.8 × 10−3 | 1.04 | 3.0 × 10−2 |
| DENND3 | hsa_circRNA_407136 | 1.3 | 3.2 × 10−3 | 1.01 | 1.5 × 10−2 | |
| FRAS1 | hsa_circRNA_103671 | hsa_circ_0070098 | 1.2 | 5.8 × 10−3 | 1.63 | 6.9 × 10−4 |
| NLRC5 | hsa_circRNA_101819 | hsa_circ_0039522 | 1.08 | 1.7 × 10−3 | 1.37 | 2.6 × 10−4 |
| XLOC_006856 | hsa_circRNA_001200 | hsa_circ_0001812 | 1.16 | 8.1 × 10−3 | 1.2 | 7.6 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, S.; Frørup, C.; Mirza, A.H.; Fløyel, T.; Yarani, R.; Colli, M.L.; Johannesen, J.; Størling, J.; Eizirik, D.L.; Pociot, F. Pro-Inflammatory Cytokines Promote the Transcription of Circular RNAs in Human Pancreatic β Cells. Non-Coding RNA 2022, 8, 69. https://doi.org/10.3390/ncrna8050069
Kaur S, Frørup C, Mirza AH, Fløyel T, Yarani R, Colli ML, Johannesen J, Størling J, Eizirik DL, Pociot F. Pro-Inflammatory Cytokines Promote the Transcription of Circular RNAs in Human Pancreatic β Cells. Non-Coding RNA. 2022; 8(5):69. https://doi.org/10.3390/ncrna8050069
Chicago/Turabian StyleKaur, Simranjeet, Caroline Frørup, Aashiq H. Mirza, Tina Fløyel, Reza Yarani, Maikel L. Colli, Jesper Johannesen, Joachim Størling, Decio L. Eizirik, and Flemming Pociot. 2022. "Pro-Inflammatory Cytokines Promote the Transcription of Circular RNAs in Human Pancreatic β Cells" Non-Coding RNA 8, no. 5: 69. https://doi.org/10.3390/ncrna8050069