
Identification of Two Exosomal miRNAs in Circulating Blood of Cancer Patients by Using Integrative Transcriptome and Network Analysis

 ,
,  , and
, and
Abstract
:1. Introduction
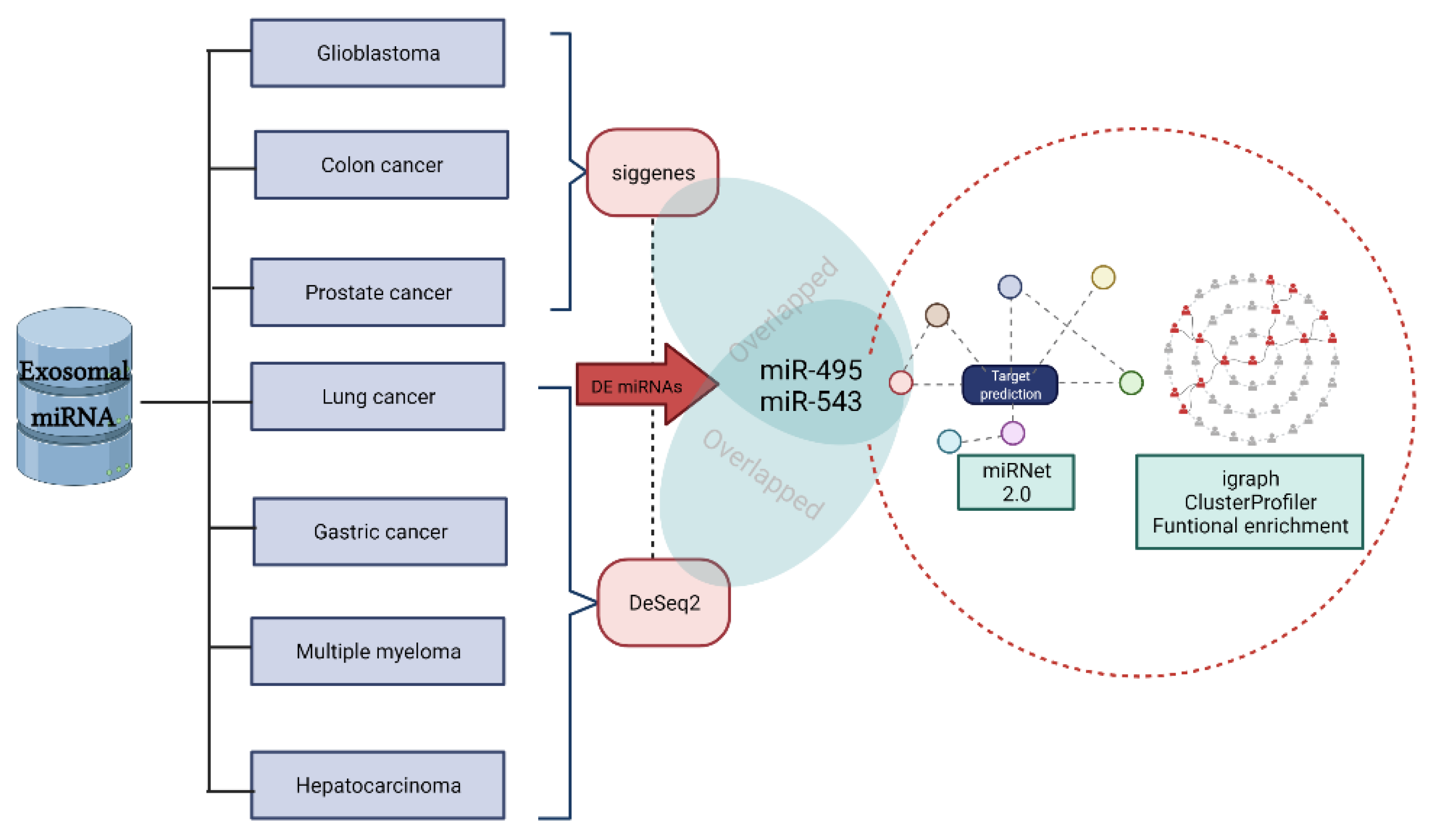
2. Results
2.1. miRNA Expression in Different Types of Cancer
2.2. External Validation of Differentially Expressed miRNAs
2.3. miRNA–mRNA Bipartite Interaction Network
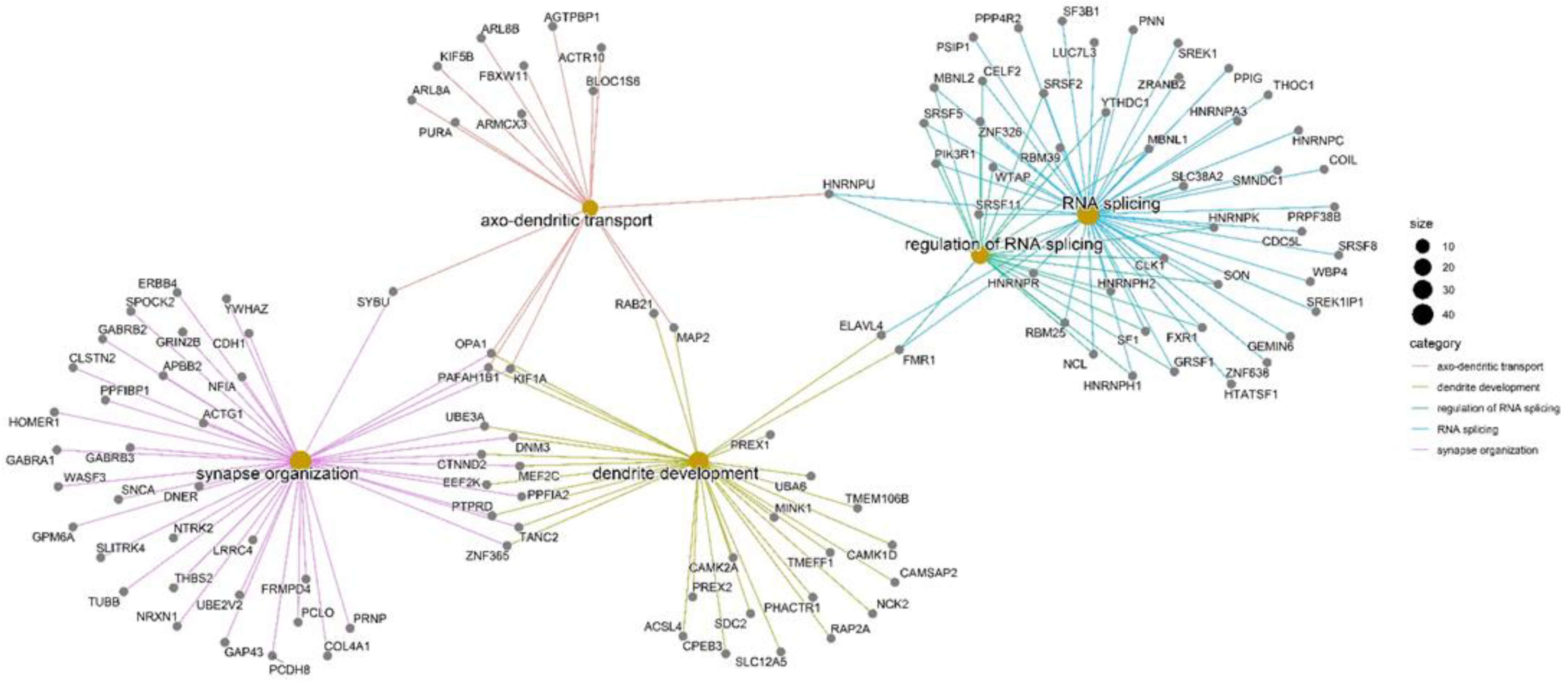
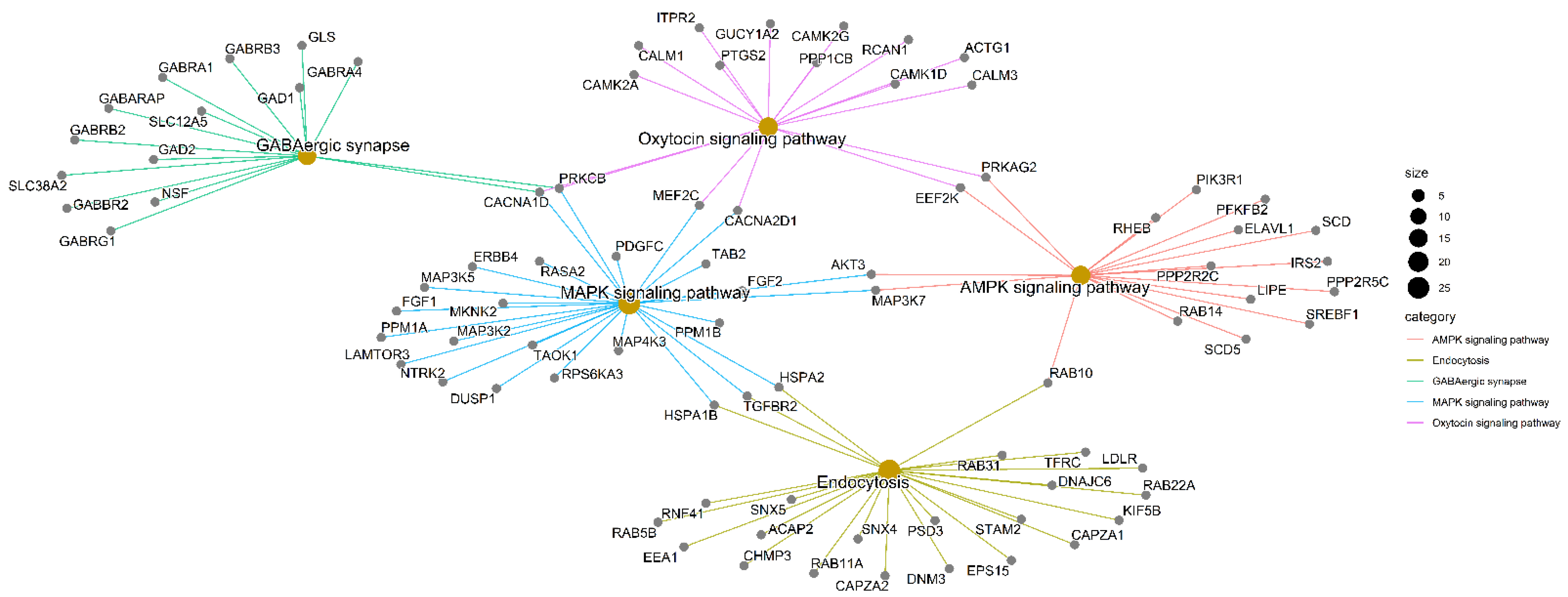
2.4. Community Detection and Functional Enrichment Analysis
3. Discussion
4. Material and Methods
4.1. High-Throughput Gene Expression Data Retrieval
4.2. Data Processing
4.3. Bipartite miRNA–Gene Network
4.4. Network Community Detection
4.5. Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3′ UTR | 3′ Untranslated region |
| AMPK | AMP-activated protein kinase |
| ceRNA | competitive endogenous RNA |
| circRNA | Circular RNA |
| CRC | Colorectal cancer |
| FDR | False Discovery Rate |
| GABA | Gamma-aminobutyric acid |
| GEA | Genomic Expression Archive |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes pathways |
| lncRNA | Long non-coding RNA |
| MAPK | Mitogen-activated protein kinase |
| miR | miRNA |
| ncRNA | non-coding RNA |
| RNA | Ribonucleic acid |
| SNARE | Soluble N-ethylmaleimide-sensitive-factor attachment protein receptor |
| sRNA | small RNA |
References
- Tsimberidou, A.M.; Fountzilas, E.; Bleris, L.; Kurzrock, R. Transcriptomics and solid tumors: The next frontier in precision cancer medicine. Semin. Cancer Biol. 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Beckers, M.; Mohorianu, I.; Stocks, M.; Applegate, C.; Dalmay, T.; Moulton, V. Comprehensive processing of high-throughput small RNA sequencing data including quality checking, normalization, and differential expression analysis using the UEA sRNA Workbench. RNA 2017, 23, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vykoukal, J.; Sun, N.; Aguilar-Bonavides, C.; Katayama, H.; Tanaka, I.; Fahrmann, J.F.; Capello, M.; Fujimoto, J.; Aguilar, M.; Wistuba, I.I.; et al. Plasma-derived extracellular vesicle proteins as a source of biomarkers for lung adenocarcinoma. Oncotarget 2017, 8, 95466–95480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semina, E.V.; Rysenkova, K.D.; Troyanovskiy, K.E.; Shmakova, A.A.; Rubina, K.A. MicroRNAs in Cancer: From Gene Expression Regulation to the Metastatic Niche Reprogramming. Biochemistry 2021, 86, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.G.; Ham, S.; Shin, H.; Chai, E.P.Z.; Lek, E.S.H.; Lobb, R.J.; Müller, A.F.; Mathivanan, S.; Yeo, B.; Choi, Y.; et al. Tumor microenvironmental cytokines bound to cancer exosomes determine uptake by cytokine receptor-expressing cells and biodistribution. Nat. Commun. 2021, 12, 3543. [Google Scholar] [CrossRef] [PubMed]
- Muluhngwi, P.; Klinge, C.M. Identification and Roles of miR-29b-1-3p and miR29a-3p-Regulated and Non-Regulated lncRNAs in Endocrine-Sensitive and Resistant Breast Cancer Cells. Cancers 2021, 13, 3530. [Google Scholar] [CrossRef]
- Romano, R.; Picca, A.; Eusebi, L.H.; Marzetti, E.; Calvani, R.; Moro, L.; Bucci, C.; Guerra, F. Extracellular Vesicles and Pancreatic Cancer: Insights on the Roles of miRNA, lncRNA, and Protein Cargos in Cancer Progression. Cells 2021, 10, 1361. [Google Scholar] [CrossRef]
- Angius, A.; Cossu-Rocca, P.; Arru, C.; Muroni, M.R.; Rallo, V.; Carru, C.; Uva, P.; Pira, G.; Orrù, S.; De Miglio, M.R. Modulatory Role of microRNAs in Triple Negative Breast Cancer with Basal-Like Phenotype. Cancers 2020, 12, 3298. [Google Scholar] [CrossRef]
- Torii, C.; Maishi, N.; Kawamoto, T.; Morimoto, M.; Akiyama, K.; Yoshioka, Y.; Minami, T.; Tsumita, T.; Alam, M.T.; Ochiya, T.; et al. miRNA-1246 in extracellular vesicles secreted from metastatic tumor induces drug resistance in tumor endothelial cells. Sci. Rep. 2021, 11, 13502. [Google Scholar] [CrossRef]
- Stocks, M.B.; Mohorianu, I.; Beckers, M.; Paicu, C.; Moxon, S.; Thody, J.; Dalmay, T.; Moulton, V. The UEA sRNA Workbench (version 4.4): A comprehensive suite of tools for analyzing miRNAs and sRNAs. Bioinformatics 2018, 34, 3382–3384. [Google Scholar] [CrossRef] [PubMed]
- Fallmann, J.; Videm, P.; Bagnacani, A.; Batut, B.; Doyle, M.A.; Klingstrom, T.; Eggenhofer, F.; Stadler, P.F.; Backofen, R.; Grüning, B. The RNA workbench 2.0: Next generation RNA data analysis. Nucleic Acids Res. 2019, 47, W511–W515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Racaud-Sultan, C.; Vergnolle, N. GSK3β, a Master Kinase in the Regulation of Adult Stem Cell Behavior. Cells 2021, 10, 225. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.-J.; Wildey, G.M.; Howe, P.H. Evidence That Ser87 of BimEL Is Phosphorylated by Akt and Regulates BimEL Apoptotic Function *. J. Biol. Chem. 2006, 281, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Jia, J.; Kumar, S.; Choi, S.W.; Gu, Y.; Mudd, M.; Dupont, N.; Jiang, S.; Peters, R.; Farzam, F.; et al. Dedicated SNAREs and specialized TRIM cargo receptors mediate secretory autophagy. EMBO J. 2017, 36, 42–60. [Google Scholar] [CrossRef]
- Yu, H.; Sun, T.; An, J.; Wen, L.; Liu, F.; Bu, Z.; Cui, Y.; Feng, J. Potential Roles of Exosomes in Parkinson’s Disease: From Pathogenesis, Diagnosis, and Treatment to Prognosis. Front. Cell Dev. Biol. 2020, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Shi, M.; Stewart, T.; Fernagut, P.-O.; Huang, Y.; Tian, C.; Dehay, B.; Atik, A.; Yang, D.; De Giorgi, F.; et al. Reduced oligodendrocyte exosome secretion in multiple system atrophy involves SNARE dysfunction. Brain 2020, 143, 1780–1797. [Google Scholar] [CrossRef]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef]
- Lee, J.E.; Kim, M.-Y. Cancer epigenetics: Past, present and future. Semin. Cancer Biol. 2021, 31, S1044-579X(21)00075-4. [Google Scholar] [CrossRef]
- Dragomir, M.; Mafra, A.C.P.; Dias, S.M.G.; Vasilescu, C.; Calin, G.A. Using microRNA Networks to Understand Cancer. Int. J. Mol. Sci. 2018, 19, 1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yang, J.; Chen, Y.; Jiang, F.; Liao, H.; Liu, X.; Wang, Y.; Kong, G.; Zhang, X.; Li, J.; et al. miRNAs derived from plasma small extracellular vesicles predict organo-tropic metastasis of gastric cancer. Gastric Cancer 2022, 25, 360–374. [Google Scholar] [CrossRef]
- Pisarska, J.; Baldy-Chudzik, K. MicroRNA-Based Fingerprinting of Cervical Lesions and Cancer. J. Clin. Med. 2020, 9, 3668. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.-S.; Zhang, E.-X.; Sun, Q.-F.; Ye, Z.-J.; Liu, J.-W.; Zhou, D.-H.; Tang, Y. Integrated analysis of lncRNA-miRNA-mRNA ceRNA network in squamous cell carcinoma of tongue. BMC Cancer 2019, 19, 779. [Google Scholar] [CrossRef] [PubMed]
- Barabási, A.-L.; Oltvai, Z.N. Network biology: Understanding the cell’s functional organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar] [CrossRef]
- Masoomy, H.; Askari, B.; Tajik, S.; Rizi, A.K.; Jafari, G.R. Topological analysis of interaction patterns in cancer-specific gene regulatory network: Persistent homology approach. Sci. Rep. 2021, 11, 16414. [Google Scholar] [CrossRef]
- Otálora-Otálora, B.A.; Florez, M.; López-Kleine, L.; Canas Arboleda, A.; Grajales Urrego, D.M.; Rojas, A. Joint Transcriptomic Analysis of Lung Cancer and Other Lung Diseases. Front. Genet. 2019, 10, 1260. [Google Scholar] [CrossRef] [Green Version]
- Capobianco, E. Next Generation Networks: Featuring the Potential Role of Emerging Applications in Translational Oncology. J. Clin. Med. 2019, 8, 664. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Hou, S.; Zou, M.; Ye, K.; Xiang, L. miR-543 impairs cell proliferation, migration, and invasion in breast cancer by suppressing VCAN. Biochem. Biophys. Res. Commun. 2021, 570, 191–198. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.-Y.; Cai, J.; Zhang, D.-J.; Zhao, L.; Luo, L.-H.; Xiong, Y.; Zhang, T.; Jin, M. MiR-543/Numb promotes proliferation, metastasis, and stem-like cell traits of prostate cancer cells. Am. J. Transl. Res. 2021, 13, 617–631. [Google Scholar] [PubMed]
- Zhou, C.; Zhao, X.; Duan, S. The role of miR-543 in human cancerous and noncancerous diseases. J. Cell. Physiol. 2021, 236, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Yang, M.; Xiao, Z.; Zhang, H.; Tan, S. circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis. Open Life Sci. 2021, 16, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Yang, T.; Xu, Y.; Ou, S.; Shi, P.; Cao, M.; Zuo, X.; Liu, Q.; Yao, J. SNHG10 Promotes Cell Proliferation and Migration in Gastric Cancer by Targeting miR-495-3p/CTNNB1 Axis. Dig. Dis. Sci. 2021, 66, 2627–2636. [Google Scholar] [CrossRef]
- Eun, J.W.; Kim, H.S.; Shen, Q.; Yang, H.D.; Kim, S.Y.; Yoon, J.H.; Park, W.S.; Lee, J.Y.; Nam, S.W. MicroRNA-495-3p functions as a tumor suppressor by regulating multiple epigenetic modifiers in gastric carcinogenesis. J. Pathol. 2018, 244, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhou, Y.; Han, H.; Li, P.; Wei, W.; Lin, N. lncRNA NEAT1 facilitates melanoma cell proliferation, migration, and invasion via regulating miR-495-3p and E2F3. J. Cell. Physiol. 2019, 234, 19592–19601. [Google Scholar] [CrossRef]
- Chen, F.; Liu, L.; Wang, S. Long non-coding RNA NORAD exhaustion represses prostate cancer progression through inhibiting TRIP13 expression via competitively binding to miR-495-3p. Cancer Cell Int. 2020, 20, 323. [Google Scholar] [CrossRef]
- Jia, Z.; Li, W.; Bian, P.; Liu, H.; Pan, D.; Dou, Z. LncRNA MCM3AP-AS1 Promotes Cell Proliferation and Invasion Through Regulating miR-543-3p/SLC39A10/PTEN Axis in Prostate Cancer. OncoTargets Ther. 2020, 13, 9365–9376. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Hallal, S.; Wei, H.; Lee, M.Y.T.; Young, P.E.; Satgunaseelan, L.; Beadnall, H.; Barnett, M.H.; Shivalingam, B.; et al. Deep sequencing of circulating exosomal microRNA allows non-invasive glioblastoma diagnosis. NPJ Precis. Oncol. 2018, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zhou, J.; Dong, M. Down-regulation of miR-543 expression increases the sensitivity of colorectal cancer cells to 5-Fluorouracil through the PTEN/PI3K/AKT pathway. Biosci. Rep. 2019, 39, BSR20190249. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhou, J.; Dong, M.; Sheng, W. Dysregulation of MicroRNA-543 expression in colorectal cancer promotes tumor migration and invasion. Mol. Carcinog. 2017, 56, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, G.; Wang, B.; Gao, W.; Yang, Q. miR-543 promotes gastric cancer cell proliferation by targeting SIRT1. Biochem. Biophys. Res. Commun. 2016, 469, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Manier, S.; Liu, C.-J.; Avet-Loiseau, H.; Park, J.; Shi, J.; Campigotto, F.; Salem, K.Z.; Huynh, D.; Glavey, S.V.; Rivotto, B.; et al. Prognostic role of circulating exosomal miRNAs in multiple myeloma. Blood 2017, 129, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Huang, H.; Li, Z.; He, C.; Li, Y.; Chen, P.; Gurbuxani, S.; Arnovitz, S.; Hong, G.-M.; Price, C.; et al. miR-495 is a tumor-suppressor microRNA down-regulated in MLL-rearranged leukemia. Proc. Natl. Acad. Sci. USA 2012, 109, 19397–19402. [Google Scholar] [CrossRef] [Green Version]
- Pelagalli, A.; Nardelli, A.; Fontanella, R.; Zannetti, A. Inhibition of AQP1 Hampers Osteosarcoma and Hepatocellular Carcinoma Progression Mediated by Bone Marrow-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2016, 17, 1102. [Google Scholar] [CrossRef] [Green Version]
- Vacca, A.; Frigeri, A.; Ribatti, D.; Nicchia, G.P.; Nico, B.; Ria, R.; Svelto, M.; Dammacco, F. Microvessel overexpression of aquaporin 1 parallels bone marrow angiogenesis in patients with active multiple myeloma. Br. J. Haematol. 2001, 113, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lin, Z.-W.; Wang, G.; Zhang, H.; Liu, B.; Xu, Q.-J. MicroRNA-495 downregulates AQP1 and facilitates proliferation and differentiation of osteoblasts in mice with tibial fracture through activation of p38 MAPK signaling pathway. Sci. Rep. 2019, 9, 16171. [Google Scholar] [CrossRef] [Green Version]
- Takehara, A.; Hosokawa, M.; Eguchi, H.; Ohigashi, H.; Ishikawa, O.; Nakamura, Y.; Nakagawa, H. γ-Aminobutyric Acid (GABA) Stimulates Pancreatic Cancer Growth through Overexpressing GABA-A Receptor π Subunit. Cancer Res. 2007, 67, 9704–9712. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; He, C.; Zhu, Y.; Wang, S.; Li, H.; Zhang, Z.; Jiang, X.; Liu, J. GABA(B)R-Induced EGFR Transactivation Promotes Migration of Human Prostate Cancer Cells. Mol. Pharmacol. 2017, 92, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.-H.; Hu, L.-P.; Wang, X.; Li, J.; Zhang, Z.-G. Neurotransmitters: Emerging targets in cancer. Oncogene 2020, 39, 503–515. [Google Scholar] [CrossRef]
- Tang, S.; Cheng, J.; Yao, Y.; Lou, C.; Wang, L.; Huang, X.; Zhang, Y. Combination of Four Serum Exosomal MiRNAs as Novel Diagnostic Biomarkers for Early-Stage Gastric Cancer. Front. Genet. 2020, 11, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, B.; Qu, S.; Hu, R.; Gao, W.; Jin, S.; Liu, M.; Zhao, Q. A panel of miRNAs derived from plasma extracellular vesicles as novel diagnostic biomarkers of lung adenocarcinoma. FEBS Open Bio 2019, 9, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Cao, J.; Chen, L.; Xi, X.; Wang, S.; Zhu, Y.; Yang, L.; Ma, L.; Wang, D.; Yin, J.; et al. Noncoding RNAs Serve as Diagnosis and Prognosis Biomarkers for Hepatocellular Carcinoma. Clin. Chem. 2019, 65, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Huang, X.; Woodcock, M.; Du, M.; Dittmar, R.; Wang, Y.; Tsai, S.; Kohli, M.; Boardman, L.; Patel, T.; et al. Plasma extracellular RNA profiles in healthy and cancer patients. Sci. Rep. 2016, 6, 19413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. miRNet 2.0: Network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-Y.; Lin, Y.-C.-D.; Li, J.; Huang, K.-Y.; Shrestha, S.; Hong, H.-C.; Tang, Y.; Chen, Y.-G.; Jin, C.-N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA–target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, U.N.; Albert, R.; Kumara, S. Near linear time algorithm to detect community structures in large-scale networks. Phys. Rev. E 2007, 76, 36106. [Google Scholar] [CrossRef] [Green Version]
- Pons, P.; Latapy, M. Computing Communities in Large Networks Using Random Walks BT—Computer and Information Sciences—ISCIS 2005; Yolum, P., Güngör, T., Gürgen, F., Özturan, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 284–293. [Google Scholar]
- Clauset, A.; Newman, M.E.J.; Moore, C. Finding community structure in very large networks. Phys. Rev. E 2004, 70, 66111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cancer | Number of Upregulated miRNAs | Number of Downregulated miRNAs |
|---|---|---|
| Prostate cancer | 6 | 25 |
| Gastric cancer | 7 | - |
| Colon cancer | 39 | - |
| Glioblastoma | 21 | |
| Multiple myeloma | 109 | 97 |
| Lung cancer | 3 | 10 |
| Liver cancer | 19 | 46 |
| Cancer | Upregulated miRNA (Fold Change) | Downregulated miRNA (Fold Change) |
|---|---|---|
| Prostate cancer | miR-543 + (1.16 × 10−6) | miR-543 (−5.07), miR-495-3p (−4.25) |
| Gastric cancer | miR-495-3p (1.86), miR-543 (1.47) miR-543 + (1.35 × 10−10) | |
| Colon cancer | miR-495-3p (4.62), miR-543 (5.7) miR-495-3p + (3.72 × 10−12) | |
| Glioblastoma | miR-543 (−4.64) miR-495-3p–(0.0120) | |
| Multiple myeloma | miR-495-3p (1.32) |
| Description | Adjusted p-Value | Gene ID |
|---|---|---|
| GABAergic synapse | 0.001 | GABRA1/NSF/GABRA4/GLS/SLC12A5/GAD1/SLC38A2/GAD2/GABBR2/GABRB2/CACNA1D/GABRG1/GABRB3/PRKCB/GABARAP |
| AMPK signaling pathway | 0.002 | SREBF1/RAB10/EEF2K/ELAVL1/PPP2R2C/PPP2R5C/LIPE/SCD/RHEB/PRKAG2/AKT3/RAB14/PFKFB2/MAP3K7/SCD5/PIK3R1/IRS2 |
| MAPK signaling pathway | 0.020 | HSPA1B/LDLR/TGFBR2/RNF41/RAB10/RAB31/TFRC/EPS15/SNX5/EEA1/RAB11A/RAB5B/ACAP2/SNX4/STAM2/CHMP3/CAPZA1/DNAJC6/RAB22A/HSPA2/PSD3/KIF5B/DNM3/CAPZA2 |
| Endocytosis | 0.0204017 | HSPA1B/LDLR/TGFBR2/RNF41/RAB10/RAB31/TFRC/EPS15/SNX5/EEA1/RAB11A/RAB5B/ACAP2/SNX4/STAM2/CHMP3/CAPZA1/DNAJC6/RAB22A/HSPA2/PSD3/KIF5B/DNM3/CAPZA2 |
| Oxytocin signaling pathway | 0.0204017 | EEF2K/CAMK2A/PTGS2/MEF2C/PRKAG2/ITPR2/CAMK2G/GUCY1A2/CACNA2D1/CACNA1D/RCAN1/CALM3/PRKCB/CAMK1D/ACTG1/CALM1/PPP1CB |
| Accession Number | Sample Type | Patient Features | Methodological Analysis |
|---|---|---|---|
| GSE130654 [51] | Extracellular vesicles derived from gastric cancer patients | 36 non-cardia adenocarcinoma patients (stages I and II) and 12 healthy individuals | DEseq2 |
| GSE111803 [52] | Extracellular vesicles derived from lung cancer patients | 5 patients with lung adenocarcinoma and 5 healthy controls | DEseq2 |
| GSE94564 [43] | Extracellular vesicles derived from multiple myeloma patients | 10 patients newly diagnosed with MM and 5 healthy individuals | DEseq2 |
| GSE123972 [53] | Extracellular vesicles derived from Hepatocellular carcinoma patients | 10 individuals with HCC pooled into 2 libraries and 10 healthy donors pooled into 2 libraries | DEseq2 |
| GSE71008 [54] | Extracellular vesicles derived from colon and prostate cancer patients | 100 colon cancer patients and 36 prostate cancer patients, and 50 healthy controls | SAM |
| GSE122488 [39] | Extracellular vesicles derived from glioblastoma patients | 12 patients with glioblastoma, 10 with glioma stages II–III, and 16 healthy controls | SAM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón-Riveros, A.; Rodríguez, J.A.; Villegas, V.E.; López-Kleine, L. Identification of Two Exosomal miRNAs in Circulating Blood of Cancer Patients by Using Integrative Transcriptome and Network Analysis. Non-Coding RNA 2022, 8, 33. https://doi.org/10.3390/ncrna8030033
Rincón-Riveros A, Rodríguez JA, Villegas VE, López-Kleine L. Identification of Two Exosomal miRNAs in Circulating Blood of Cancer Patients by Using Integrative Transcriptome and Network Analysis. Non-Coding RNA. 2022; 8(3):33. https://doi.org/10.3390/ncrna8030033
Chicago/Turabian StyleRincón-Riveros, Andrés, Josefa Antonia Rodríguez, Victoria E. Villegas, and Liliana López-Kleine. 2022. "Identification of Two Exosomal miRNAs in Circulating Blood of Cancer Patients by Using Integrative Transcriptome and Network Analysis" Non-Coding RNA 8, no. 3: 33. https://doi.org/10.3390/ncrna8030033