
Performance of a Helical Microswimmer Traversing a Discrete Viscoelastic Network with Dynamic Remodeling


Abstract
:1. Introduction
2. Methods
2.1. The Computational Framework
2.2. The Network
2.3. The Swimmer
2.4. Repulsion and Anchoring Forces
2.5. Regularized Stokeslet Solution
2.6. Instantaneous Kinematics
2.7. Temporal Evolution
2.8. Remodeling
3. Simulation Results
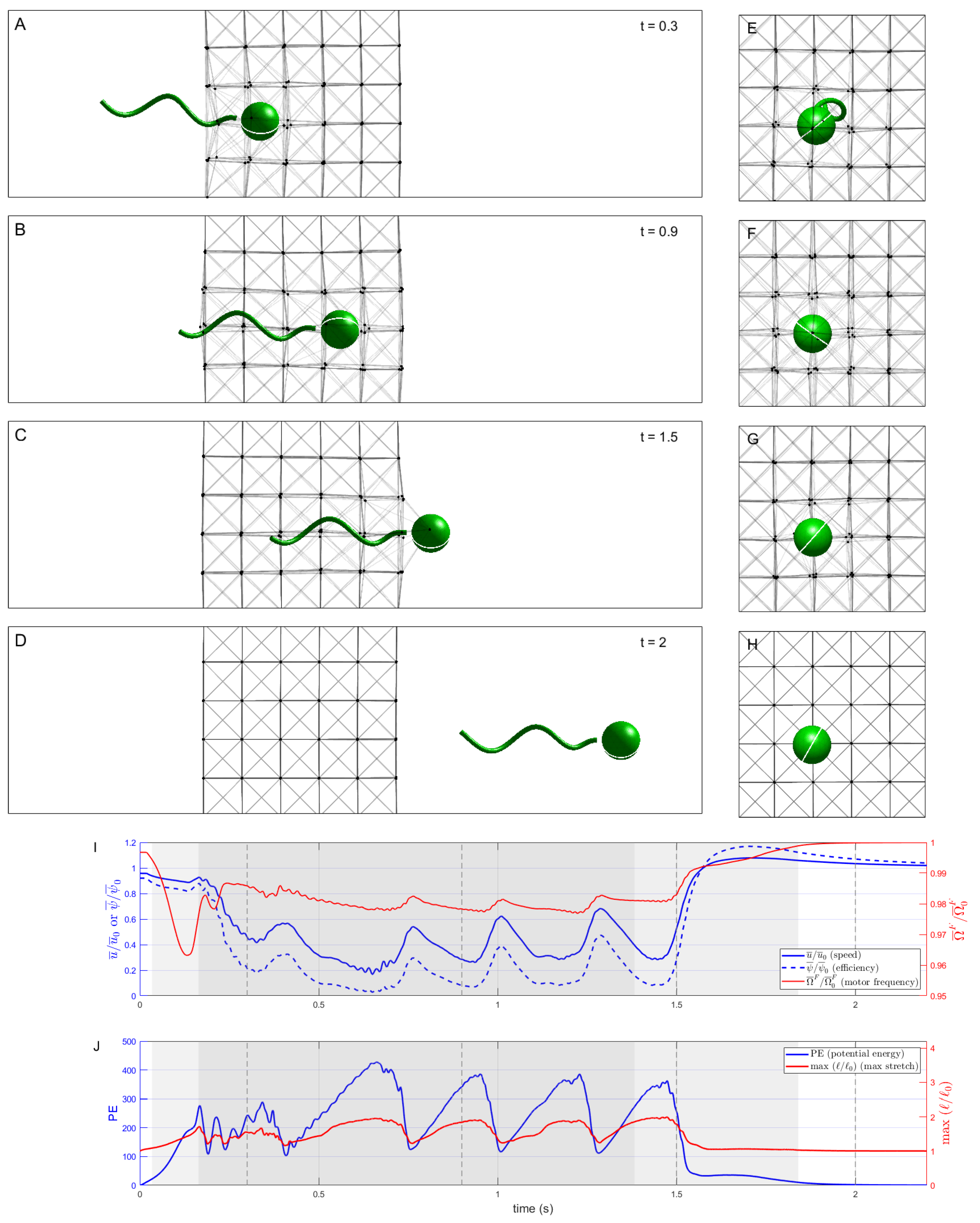
3.1. No Remodeling of Network
3.2. Remodeling of Network Due to Dissolution
3.3. Effects of Varying E,
4. Discussion
5. Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fauci, L.; Dillon, R. Biofluidmechanics of Reproduction. Annu. Rev. Fluid. Mech. 2006, 38, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Guasto, J.S.; Rusconi, R.; Stocker, R. Fluid Mechanics of Planktonic Microorganisms. Ann. Rev. Fluid Mech. 2012, 44, 373–400. [Google Scholar] [CrossRef] [Green Version]
- Lauga, E.; Powers, T. The hydrodynamics of swimming microorganisms. Rep. Prog. Phys. 2009, 72, 096601. [Google Scholar] [CrossRef]
- Lauga, E. Bacterial hydrodynamics. Annu. Rev. Fluid Mech. 2016, 48, 105–130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Abbott, J.; Dong, L.; Peyer, K.; Kratochvil, B.; Zhang, H.; Bergeles, C.; Nelson, B. Characterizing the swimming properties of artificial bacterial flagella. Nano Lett. 2009, 9, 3663–3667. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, E.; Gadehla, H.; Smith, D.; Blake, J.; Kirkman-Brown, J. Mammalian Sperm Motility: Observation and Theory. Ann. Rev. Fluid Mech. 2011, 43, 501–528. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Chen, Y.; Mukasa, D.; Pak, O.S.; Gao, W. Medical micro/nanorobots in complex media. Chem. Soc. Rev. 2020, 49, 8088–8112. [Google Scholar] [CrossRef]
- Primakoff, P.; Myles, D.G. Penetration, Adhesion, and Fusion in Mammalian Sperm-Egg Interaction. Science 2002, 296, 2183–2185. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Yamashita, M.; Kimura, M.; Honda, A.; Kashiwabara, S.I.; Baba, T. Sperm penetration through cumulus mass and zona pellucida. Int. J. Dev. Biol. 2008, 52, 677–682. [Google Scholar] [CrossRef]
- Schwartz, P.; Hinney, B.; Nayudu, P.; Michelmann, H. Oocyte-sperm interaction in the course of IVF: A scanning electron microscopy analysis. Reprod. Biomed. Online 2003, 7, 205–210. [Google Scholar] [CrossRef]
- Harman, M.W.; Dunham-Ems, S.M.; Caimano, M.J.; Belperron, A.A.; Bockenstedt, L.K.; Fu, H.C.; Radolf, J.D.; Wolgemuth, C.W. The heterogeneous motility of the Lyme disease spirochete in gelatin mimics dissemination through tissue. Proc. Natl. Acad. Sci. USA 2012, 109, 3059–3064. [Google Scholar] [CrossRef] [Green Version]
- Crossland, N.A.; Alvarez, X.; Embers, M.E. Late Disseminated Lyme Disease: Associated Pathology and Spirochete Persistence Posttreatment in Rhesus Macaques. Am. J. Pathol. 2018, 188, 672–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa-Morales, N.; Dominguez-Rubio, L.; Ott, T.L.; Aranson, I.S. Mechanical shear controls bacterial penetration in mucus. Sci. Rep. 2019, 9, 9713. [Google Scholar] [CrossRef] [Green Version]
- Bakshani, C.R.; Morales-Garcia, A.L.; Althaus, M.; Wilcox, M.D.; Pearson, J.P.; Bythell, J.C.; Burgess, J.G. Evolutionary conservation of the antimicrobial function of mucus: A first defence against infection. npj Biofilms Microbiomes 2018, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celli, J.P.; Turner, B.S.; Afdhal, N.H.; Keates, S.; Ghiran, I.; Kelly, C.P.; Ewoldt, R.H.; McKinley, G.H.; So, P.; Erramilli, S.; et al. Helicobacter pylori moves through mucus by reducing mucin viscoelasticity. Proc. Natl. Acad. Sci. USA 2009, 106, 14321–14326. [Google Scholar] [CrossRef] [Green Version]
- Mirbagheri, S.; Fu, H. Helicobacter pylori couples motility and diffusion to actively create a heterogeneous complex medium in gastric mucus. Phys. Rev. Lett. 2016, 116, 198101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Du, X.; Jin, D.; Zhang, L. Real-time ultrasound doppler tracking and autonomous navigation of a miniature helical robot for accelerating thrombolysis in dynamic blood flow. ACS Nano 2022, 16, 604–616. [Google Scholar] [CrossRef]
- Schamel, D.; Mark, A.G.; Gibbs, J.G.; Miksch, C.; Morozov, K.I.; Leshansky, A.M.; Fischer, P. Nanopropellers and Their Actuation in Complex Viscoelastic Media. ACS Nano 2014, 8, 8794–8801. [Google Scholar] [CrossRef]
- Walker, D.; Käsdorf, B.T.; Jeong, H.H.; Lieleg, O.; Fischer, P. Enzymatically active biomimetic micropropellers for the penetration of mucin gels. Sci. Adv. 2015, 1, e1500501. [Google Scholar] [CrossRef] [Green Version]
- Lauga, E. Propulsion in a viscoelastic fluid. Phys. Fluids 2007, 19, 0989104. [Google Scholar] [CrossRef] [Green Version]
- Spagnolie, S.E.; Liu, B.; Powers, T.R. Locomotion of Helical Bodies in Viscoelastic Fluids: Enhanced Swimming at Large Helical Amplitudes. Phys. Rev. Lett. 2013, 111, 068101. [Google Scholar] [CrossRef] [Green Version]
- Ho, N.; Olson, S.; Leiderman, K. Swimming speeds of filaments in viscous fluids with resistance. Phys. Rev. E 2016, 93, 043108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teran, J.; Fauci, L.; Shelley, M. Viscoelastic Fluid Response Can Increase the Speed and Efficiency of a Free Swimmer. Phys. Rev. Lett. 2010, 104, 038101. [Google Scholar] [CrossRef]
- Elfring, G.; Goyal, G. The effect of gait on swimming in viscoelastic fluids. J. Non-Newton. Fluid Mech. 2016, 234, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Thomases, B.; Guy, R.D. Mechanisms of Elastic Enhancement and Hindrance for Finite-Length Undulatory Swimmers in Viscoelastic Fluids. Phys. Rev. Lett. 2014, 113, 098102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomases, B.; Guy, R.D. The role of body flexibility in stroke enhancements for finite-length undulatory swimmers in viscoelastic fluids. J. Fluid Mech. 2017, 825, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Wróbel, J.K.; Cortez, R.; Fauci, L. Modeling viscoelastic networks in Stokes flow. Phys. Fluids 2014, 26, 388–391. [Google Scholar] [CrossRef] [Green Version]
- Starr, M.P.; Stolp, H.; Trüper, H.G.; Balows, A.; Schlegel, H.G. (Eds.). The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- Curtis, M.; Kirkman-Brown, J.; Connolly, T.; Gaffney, E. Modelling a tethered mammalian sperm cell undergoing hyperactivation. J. Theor. Biol. 2012, 309, 1–10. [Google Scholar] [CrossRef]
- Higdon, J.J.L. The hydrodynamics of flagellar propulsion: Helical waves. J. Fluid. Mech. 1979, 94, 331–351. [Google Scholar] [CrossRef] [Green Version]
- Pak, O.; Spagnolie, S.; Lauga, E. Hydrodynamics of the double-wave structure of insect spermatozoa flagella. J. R. Soc. Interface 2012, 9, 1908–1924. [Google Scholar] [CrossRef]
- Jung, S.; Mareck, K.; Fauci, L.; Shelley, M. Rotational dynamics of a superhelix towed in a Stokes fluid. Phys. Fluids 2007, 19, 103105. [Google Scholar] [CrossRef] [Green Version]
- Bottino, D.C. Modeling Viscoelastic Networks and Cell Deformation in the Context of the Immersed Boundary Method. J. Comput. Phys. 1998, 147, 86–113. [Google Scholar] [CrossRef]
- Fai, T.; Leo-Macias, A.; Stokes, D.; Peskin, C. Image-based model of the spectrin cytoskeleton for red blood cell simulation. PLOS Comput. Biol. 2017, 13, e1005790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wróbel, J.K.; Lynch, S.; Barrett, A.; Fauci, L.; Cortez, R. Enhanced flagellar swimming through a compliant viscoelastic network in Stokes flow. J. Fluid Mech. 2016, 792, 775–797. [Google Scholar] [CrossRef]
- Cortez, R. The method of regularized Stokeslets. SIAM J. Sci. Comput. 2001, 23, 1204–1225. [Google Scholar] [CrossRef]
- Cortez, R.; Fauci, L.; Medovikov, A. The method of regularized Stokeslets in three dimensions: Analysis, validation, and application to helical swimming. Phys. Fluids 2005, 17, 031504. [Google Scholar] [CrossRef]
- Shum, H.; Gaffney, E.A.; Smith, D.J. Modelling bacterial behaviour close to a no-slip plane boundary: The influence of bacterial geometry. Proc. R. Soc. A Math. Phys. Eng. Sci. 2010, 466, 1725–1748. [Google Scholar] [CrossRef] [Green Version]
- Schuech, R.; Hoehfurtner, T.; Smith, D.J.; Humphries, S. Motile curved bacteria are Pareto-optimal. Proc. Natl. Acad. Sci. USA 2019, 116, 14440–14447. [Google Scholar] [CrossRef] [Green Version]
- Ribes, A.; Caremoli, C. Salome platform component model for numerical simulation. In Proceedings of the Computer Software and Applications Conference, COMPSAC 2007, 31st Annual International, Beijing, China, 24–27 July 2007; Volume 2, pp. 553–564. [Google Scholar] [CrossRef]
- Schöberl, J. NETGEN An advancing front 2D/3D-mesh generator based on abstract rules. Comput. Vis. Sci. 1997, 1, 41–52. [Google Scholar] [CrossRef]
- Walker, B.J.; Wheeler, R.J.; Ishimoto, K.; Gaffney, E.A. Boundary behaviours of Leishmania mexicana: A hydrodynamic simulation study. J. Theor. Biol. 2019, 462, 311–320. [Google Scholar] [CrossRef]
- Ishimoto, K.; Gaffney, E. Mechanical tuning of mammalian sperm behaviour by hyperactivation, rheology and substrate adhesion: A numerical exploration. J. R. Soc. Interface 2016, 13, 20160633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimoto, K.; Cosson, J.; Gaffney, E. A simulation study of sperm motility hydrodynamics near fish eggs and spheres. J. Theor. Biol. 2016, 389, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Elgeti, J.; Kaupp, U.; Gompper, G. Hydrodynamics of sperm cells near surfaces. Biophys. J. 2010, 99, 1018–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genz, A.; Cools, R. An adaptive numerical cubature algorithm for simplices. ACM Trans. Math. Softw. (TOMS) 2003, 29, 297–308. [Google Scholar] [CrossRef]
- Pozrikidis, C. A Practical Guide to Boundary Element Methods with the Software Library BEMLIB; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Angeles, V.; Godínez, F.A.; Puente-Velazquez, J.A.; Mendez-Rojano, R.; Lauga, E.; Zenit, R. Front-back asymmetry controls the impact of viscoelasticity on helical swimming. Phys. Rev. Fluids 2021, 6, 043102. [Google Scholar] [CrossRef]
- Rutllant, J.; López-Béjar, M.; López-Gatius, F. Ultrastructural and rheological properties of bovine vaginal fluid and its relation to sperm motility and fertilization: A review. Reprod. Domest. Anim. 2005, 40, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Talbot, P.; Geiske, C.; Knoll, M. Oocyte Pickup by the Mammalian Oviduct. Mol. Biol. Cell 1999, 10, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Button, B.; Cai, L.H.; Ehre, C.; Kesimer, M.; Hill, D.B.; Sheehan, J.K.; Boucher, R.C.; Rubinstein, M. Periciliary Brush Promotes the Lung Health by Separating the Mucus Layer from Airway Epithelia. Science 2012, 337, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T. Suspension biomechanics of swimming microbes. J. R. Soc. Interface 2009, 6, rsif20090223. [Google Scholar] [CrossRef] [Green Version]
- Sretenovic, S.; Stojković, B.; Dogsa, I.; Kostanjšek, R.; Poberaj, I.; Stopar, D. An early mechanical coupling of planktonic bacteria in dilute suspensions. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Parameter | Value |
|---|---|---|
| Swimmer | ||
| Body | radius, | 0.6204 m |
| Flagellum | diameter | 0.16 m |
| amplitude, a | 0.4 m | |
| wavelength, | 2.9 m | |
| # wavelengths | 1.5 | |
| exponential decay constant, | 1.543 m | |
| Network | ||
| length, width, height | 6.2 m, 13.6 m, 13.6 m | |
| # layers in swimming direction | 6 | |
| connectivity | 18 links per node (6 along x, y, z, 12 diagonal) | |
| spacing between layers, min | 0.6204 m | |
| spring stiffness, E | 0–100 mPa | |
| dashpot constant, | 0.1–10, ∞ mPa s | |
| fluid viscosity, | 1 mPa s | |
| Numerical | ||
| Swimmer | total number of nodes, | 1780 |
| Body | ||
| regularization parameter, | m | |
| number of nodes and triangular elements | 454 and 226 | |
| Flagellum | ||
| regularization parameter, | m | |
| number of nodes and triangular elements | 1326 and 662 | |
| applied motor torque, | 1000 fN m | |
| Network | ||
| regularization parameter, | 0.0796 m | |
| total number of nodes () and links | 864 and 6396 | |
| number of anchored nodes, | 264 | |
| dissolution distance | m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuech, R.; Cortez, R.; Fauci, L. Performance of a Helical Microswimmer Traversing a Discrete Viscoelastic Network with Dynamic Remodeling. Fluids 2022, 7, 257. https://doi.org/10.3390/fluids7080257
Schuech R, Cortez R, Fauci L. Performance of a Helical Microswimmer Traversing a Discrete Viscoelastic Network with Dynamic Remodeling. Fluids. 2022; 7(8):257. https://doi.org/10.3390/fluids7080257
Chicago/Turabian StyleSchuech, Rudi, Ricardo Cortez, and Lisa Fauci. 2022. "Performance of a Helical Microswimmer Traversing a Discrete Viscoelastic Network with Dynamic Remodeling" Fluids 7, no. 8: 257. https://doi.org/10.3390/fluids7080257