
Influence of Diameter and Cyclic Mechanical Stimulation on the Beating Frequency of Myocardial Cell-Laden Fibers
 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
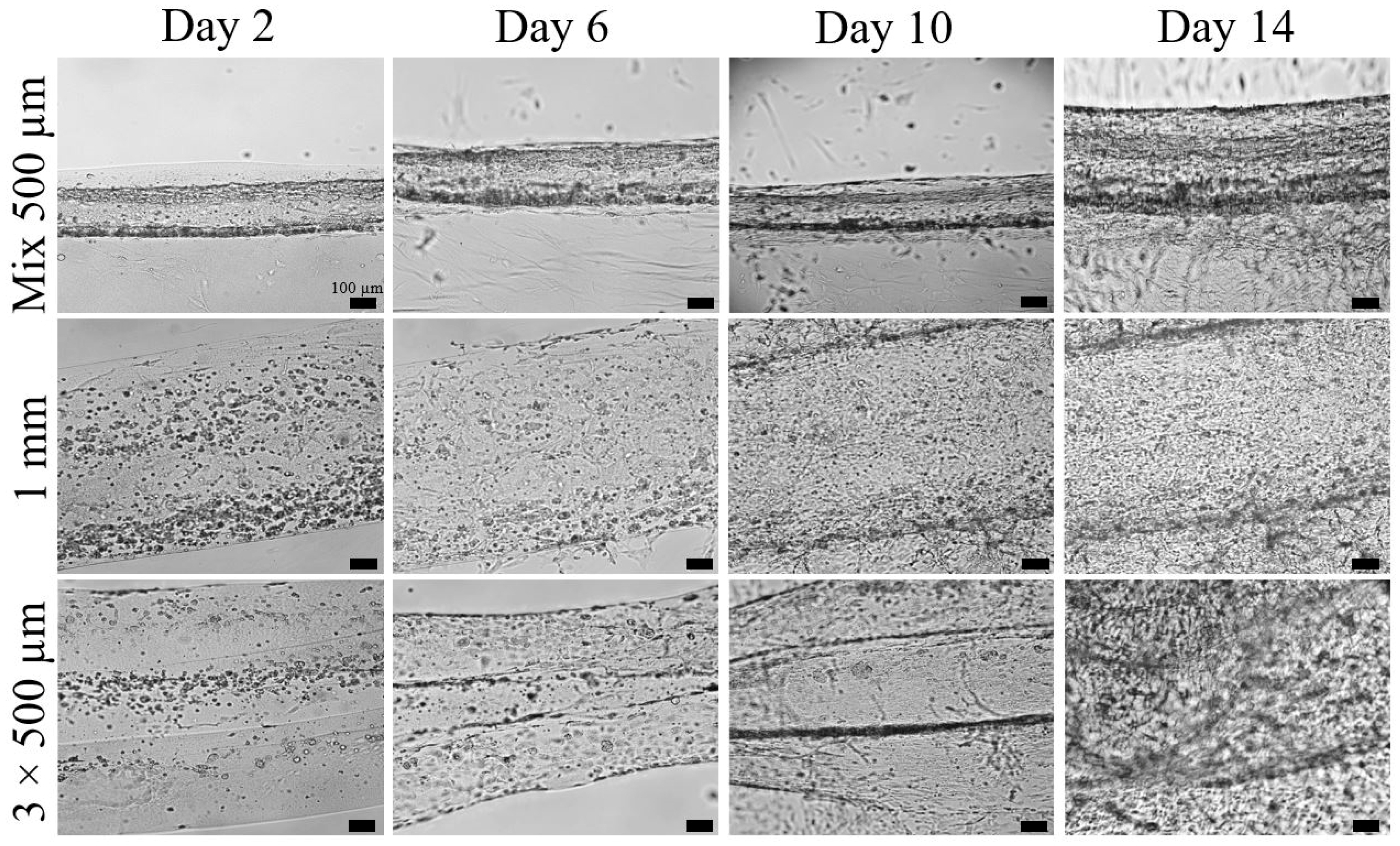
2.1. Cell Infiltration and Proliferation
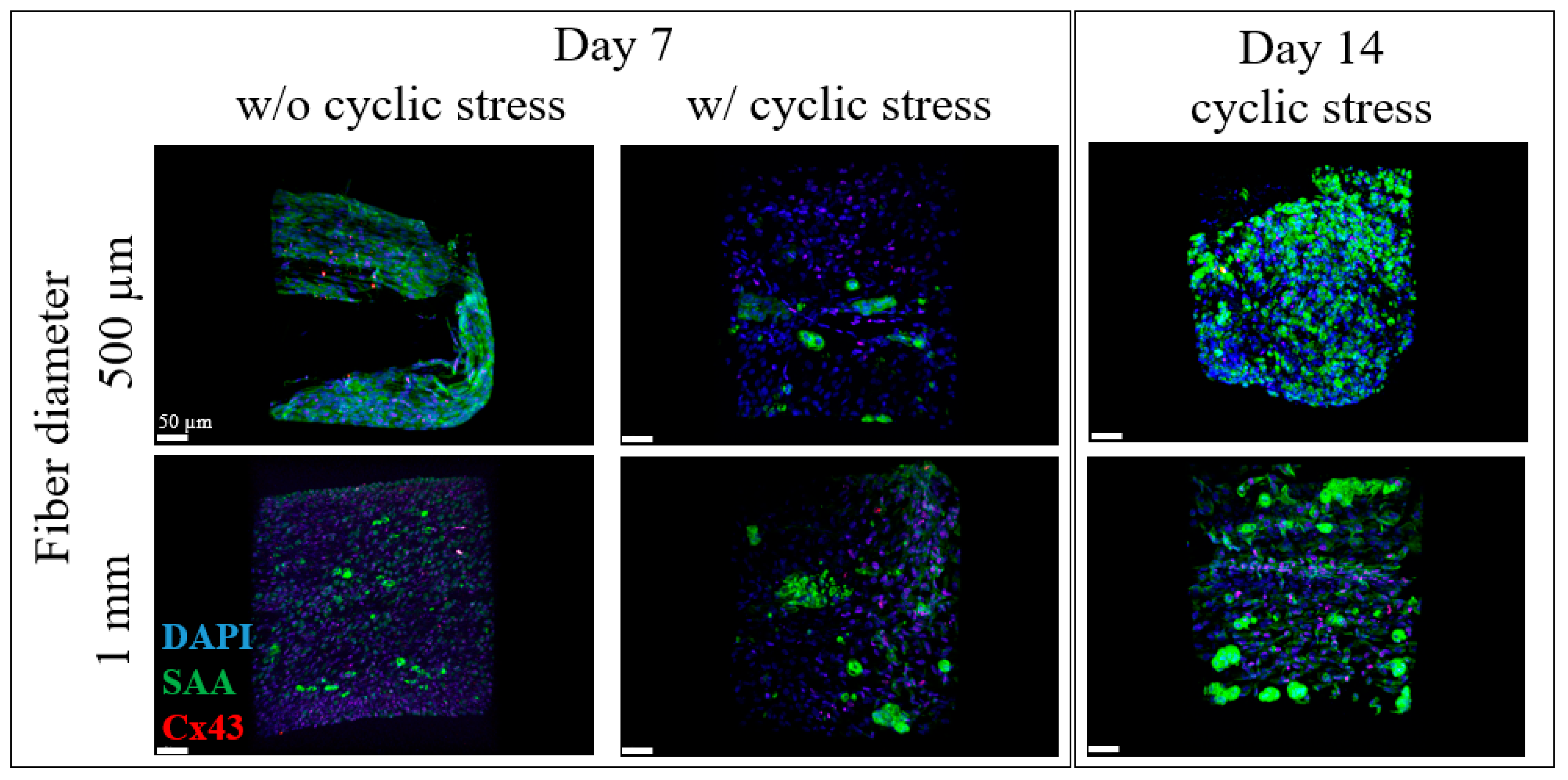
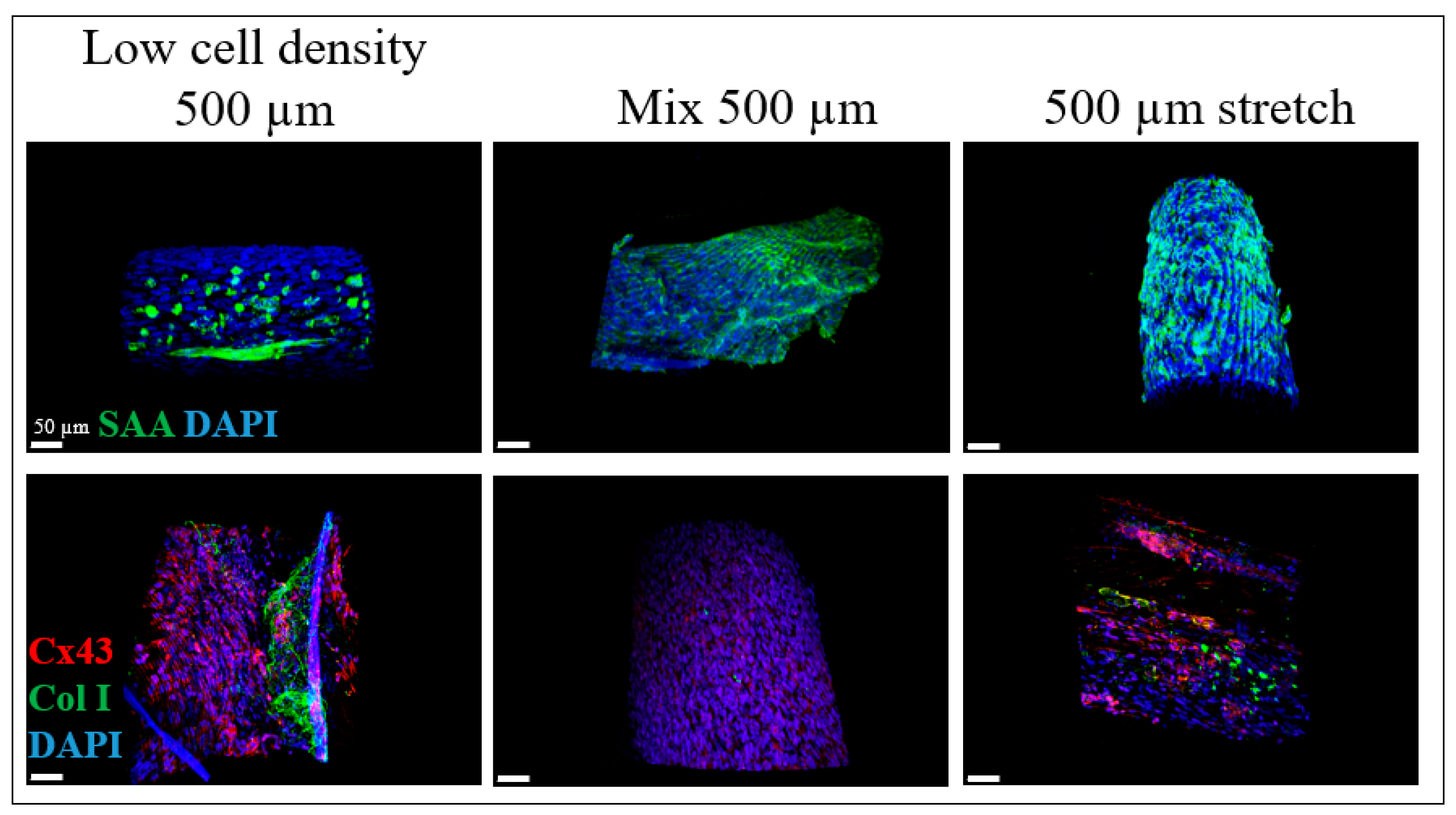
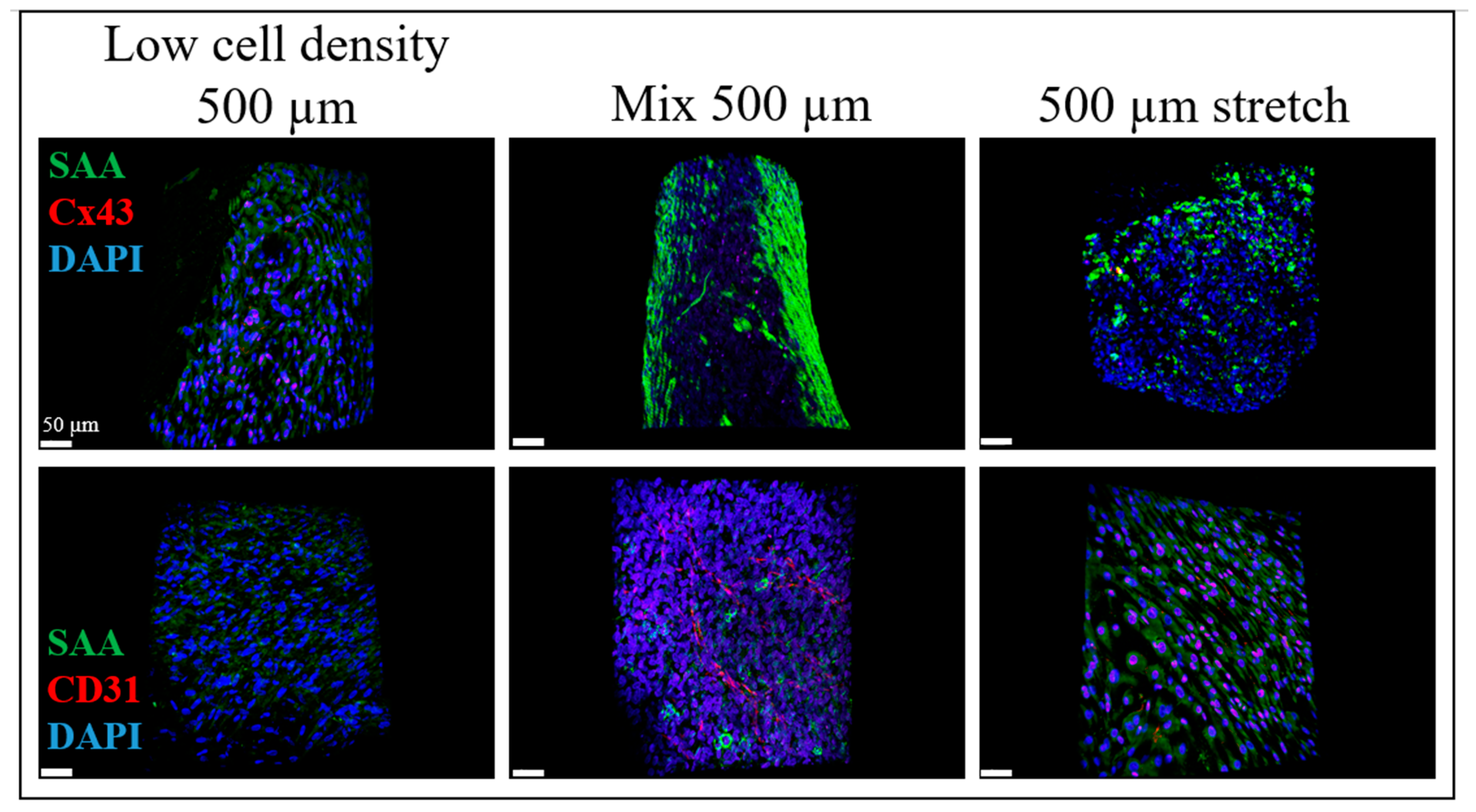
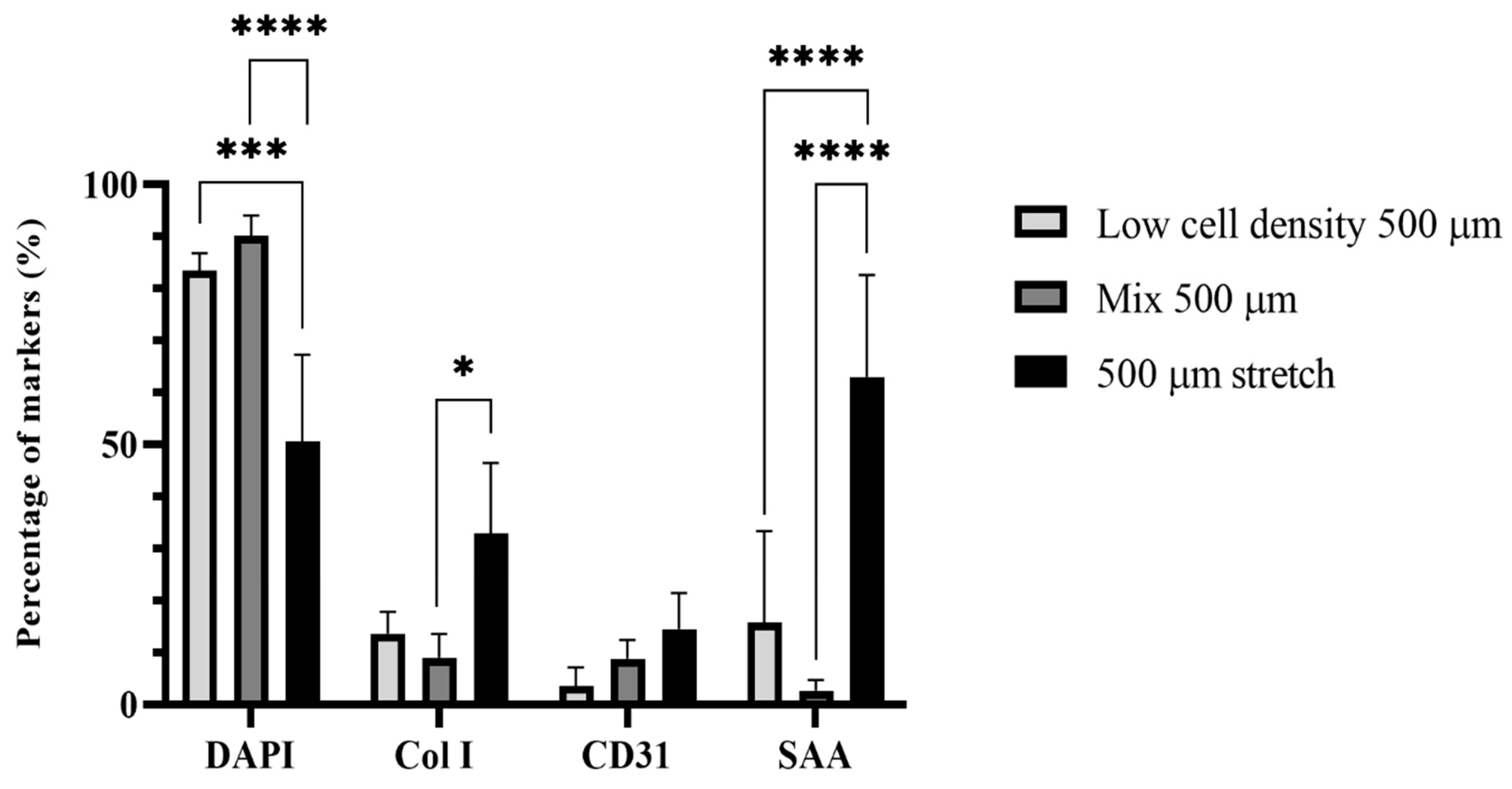
2.2. iPSC-CM Markers for the BioPacer
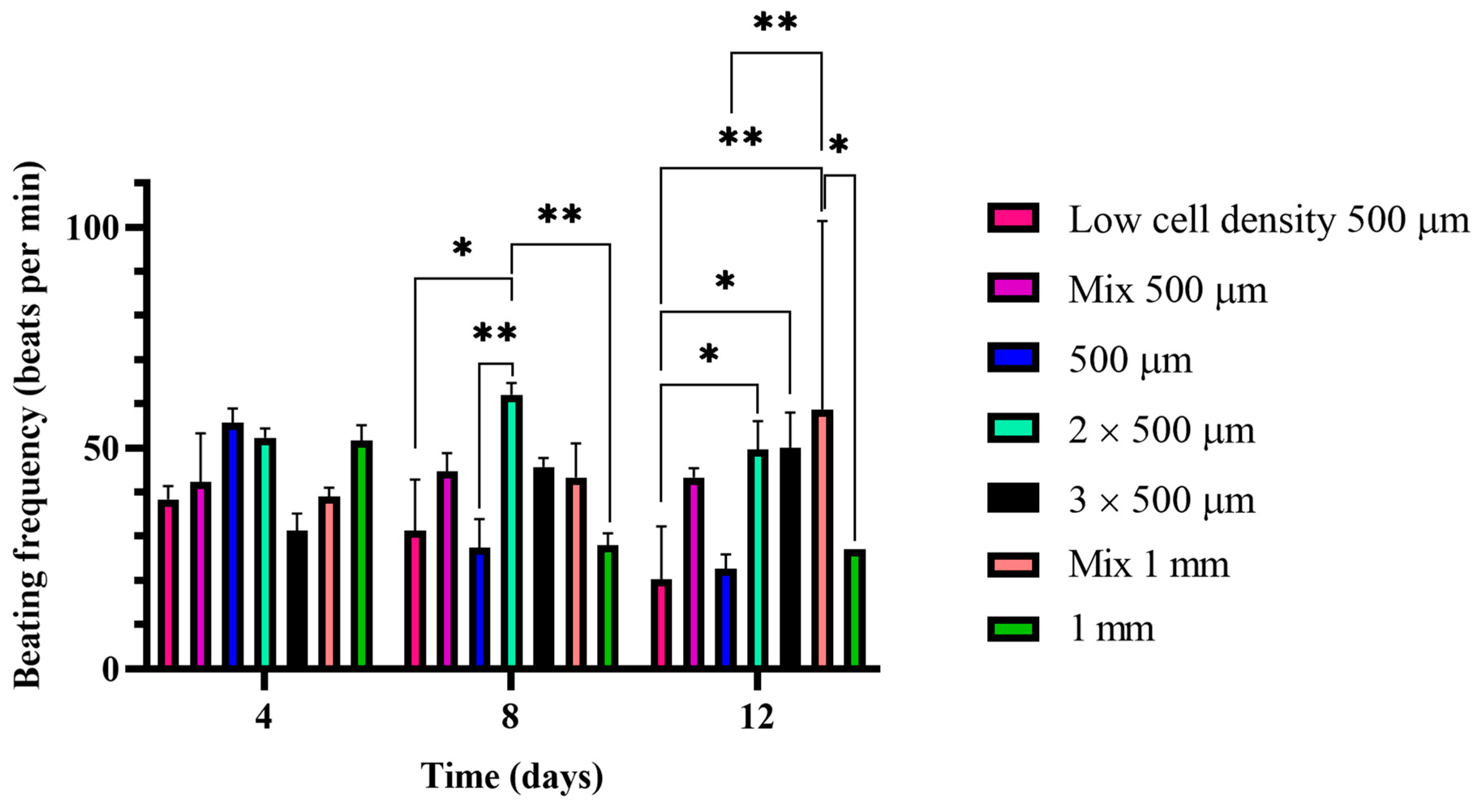
2.3. Beating Frequency
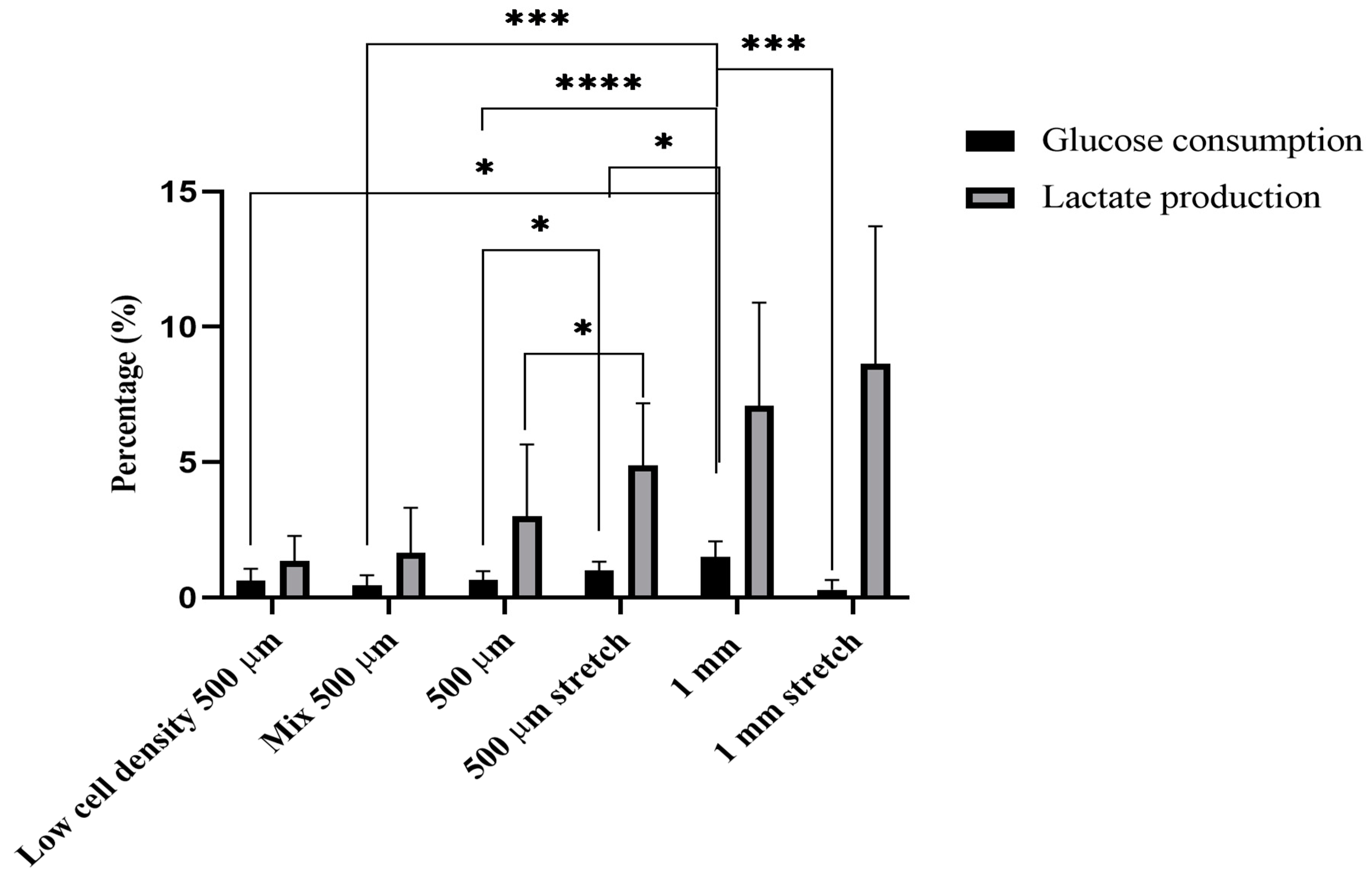
2.4. Glucose and Lactate Measurements
2.5. Discussion
3. Conclusions
4. Methods
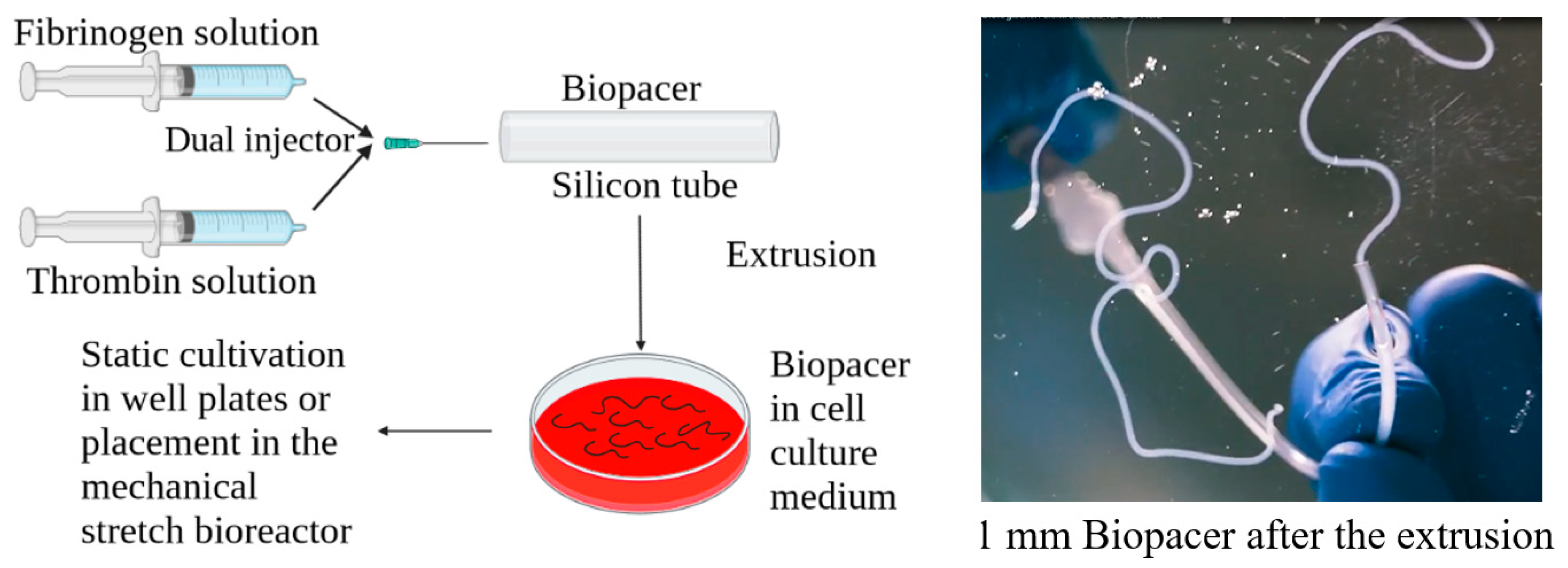
4.1. Cylindrical Fibrin Gel Constructs
4.2. Cell Culturing
4.3. Bioreactor Set-Up
4.4. Glucose and Lactate Measurements
4.5. Immunohistochemistry
4.6. Beating Frequency
4.7. Quantification of the Markers Content
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baruteau, A.E.; Pass, R.H.; Thambo, J.B.; Behaghel, A.; Le Pennec, S.; Perdreau, E.; Combes, N.; Liberman, L.; McLeod, C.J. Congenital and childhood atrioventricular blocks: Pathophysiology and contemporary management. Eur. J. Pediatr. 2016, 175, 1235–1248. [Google Scholar] [CrossRef]
- Bartakian, S.; Fagan, T.E.; Schaffer, M.S.; Darst, J.R. Device closure of secundum atrial septal defects in children <15 kg: Complication rates and indications for referral. JACC Cardiovasc. Interv. 2012, 5, 1178–1184. [Google Scholar]
- Vogler, J.; Breithardt, G.; Eckardt, L. Bradyarrhythmias and Conduction Blocks. Rev. Española Cardiol. (Engl. Ed.) 2012, 65, 656–667. [Google Scholar] [CrossRef]
- Aksu, T.; Gopinathannair, R.; Bozyel, S.; Yalin, K.; Gupta, D. Cardioneuroablation for Treatment of Atrioventricular Block. Circ. Arrhythmia Electrophysiol. 2021, 14, e010018. [Google Scholar] [CrossRef]
- Yeung, C.; Baranchuk, A. Systematic Approach to the Diagnosis and Treatment of Lyme Carditis and High-Degree Atrioventricular Block. Healthcare 2018, 6, 119. [Google Scholar] [CrossRef]
- Pyngottu, A.; Werner, H.; Lehmann, P.; Balmer, C. Health-Related Quality of Life and Psychological Adjustment of Children and Adolescents with Pacemakers and Implantable Cardioverter Defibrillators: A Systematic Review. Pediatr. Cardiol. 2019, 40, 1–16. [Google Scholar] [CrossRef]
- Gheissari, A.; Hordof, A.J.; Spotnitz, H.M. Transvenous pacemakers in children: Relation of lead length to anticipated growth. Ann. Thorac. Surg. 1991, 52, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Song, M.K.; Kim, N.Y.; Bae, E.J.; Kim, G.B.; Kwak, J.G.; Kim, W.H.; Lee, J.R. Long-term Follow-up of Epicardial Pacing and Left Ventricular Dysfunction in Children With Congenital Heart Block. Ann. Thorac. Surg. 2020, 109, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Silvetti, M.S.; Drago, F.; Ravà, L. Determinants of early dilated cardiomyopathy in neonates with congenital complete atrio-ventricular block. Europace 2010, 12, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Cate, F.E.U.T.; Breur, J.M.; Cohen, I.M.; Boramanand, N.; Kapusta, L.; Crosson, E.J.; Brenner, J.I.; Lubbers, L.J.; Friedman, A.H.; Vetter, V.L.; et al. Dilated cardiomyopathy in isolated congenital complete atrioventricular block: Early and long-term risk in children. J. Am. Coll. Cardiol. 2001, 37, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Liberman, L.; Silver, E.S.; Chai, P.J.; Anderson, B.R. Incidence and characteristics of heart block after heart surgery in pediatric patients: A multicenter study. J. Thorac. Cardiovasc. Surg. 2016, 152, 197–202. [Google Scholar] [CrossRef]
- McLeod, K.A. Cardiac pacing in infants and children. Heart 2010, 96, 1502–1508. [Google Scholar] [CrossRef]
- Gorabi, A.M.; Hajighasemi, S.; Khori, V.; Soleimani, M.; Rajaei, M.; Rabbani, S.; Atashi, A.; Ghiaseddin, A.; Saeid, A.K.; Tafti, H.A.; et al. Functional biological pacemaker generation by T-Box18 protein expression via stem cell and viral delivery approaches in a murine model of complete heart block. Pharmacol. Res. 2019, 141, 443–450. [Google Scholar] [CrossRef]
- Lu, W.; Yaoming, N.; Boli, R.; Jun, C.; Changhai, Z.; Yang, Z.; Zhiyuan, S. mHCN4 Genetically Modified Canine Mesenchymal Stem Cells Provide Biological Pacemaking Function in Complete Dogs with Atrioventricular Block. Pacing Clin. Electrophysiol. 2013, 36, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Yokokawa, M.; Ohnishi, S.; Ishibashi-Ueda, H.; Obata, H.; Otani, K.; Miyahara, Y.; Tanaka, K.; Shimizu, W.; Nakazawa, K.; Kangawa, K. Transplantation of mesenchymal stem cells improves atrioventricular conduction in a rat model of com-plete atrioventricular block. Cell Transplant. 2008, 17, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, S.; Anyukhovsky, E.P.; Ben-Ari, M.; Naor, S.; Jiang, Y.-P.; Danilo, P.; Rahim, T.; Burke, S.; Qiu, X.; Potapova, I.A.; et al. Induced Pluripotent Stem Cell–Derived Cardiomyocytes Provide In Vivo Biological Pacemaker Function. Circ. Arrhythmia Electrophysiol. 2017, 10, e004508. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, E.; Ionta, V.; Cheng, K.; Giacomello, A.; Cho, H.C.; Marbán, E. Engineered electrical conduction tract restores conduction in complete heart block: From in vitro to in vivo proof of concept. J. Am. Coll. Cardiol. 2014, 64, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Miklas, J.W.; Nunes, S.S.; Sofla, A.; Reis, L.A.; Pahnke, A.; Xiao, Y.; Laschinger, C.; Radisic, M. Bioreactor for modulation of cardiac microtissue phenotype by combined static stretch and electrical stim-ulation. Biofabrication 2014, 6, 024113. [Google Scholar] [CrossRef]
- Nunes, S.S.; Miklas, J.W.; Liu, J.; Aschar-Sobbi, R.; Xiao, Y.; Zhang, B.; Jiang, J.; Massé, S.; Gagliardi, M.; Hsieh, A.; et al. Biowire: A platform for maturation of human pluripotent stem cell–derived cardiomyocytes. Nat. Methods 2013, 10, 781–787. [Google Scholar] [CrossRef]
- Kensah, G.; Lara, A.R.; Dahlmann, J.; Zweigerdt, R.; Schwanke, K.; Hegermann, J.; Skvorc, D.; Gawol, A.; Azizian, A.; Wagner, S.; et al. Murine and human pluripotent stem cell-derived cardiac bodies form contractile myocardial tissue in vitro. Eur. Heart J. 2013, 34, 1134–1146. [Google Scholar] [CrossRef]
- Keijdener, H.; Konrad, J.; Hoffmann, B.; Gerardo-Nava, J.; Rütten, S.; Merkel, R.; Vázquez-Jiménez, J.; Brook, G.A.; Jockenhoevel, S.; Mela, P. A bench-top molding method for the production of cell-laden fibrin micro-fibers with longitudinal topog-raphy. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 1198–1212. [Google Scholar] [CrossRef] [PubMed]
- Jockenhoevel, S.; Zund, G.; Hoerstrup, S.P.; Chalabi, K.; Sachweh, J.S.; Demircan, L.; Messmer, B.J.; Turina, M. Fibrin gel—Advantages of a new scaffold in cardiovascular tissue engineering. Eur. J. Cardio-Thorac. Surg. 2001, 19, 424–430. [Google Scholar] [CrossRef]
- de Melo, B.A.; Jodat, Y.A.; Cruz, E.M.; Benincasa, J.C.; Shin, S.R.; Porcionatto, M.A. Strategies to use fibrinogen as bioink for 3D bioprinting fibrin-based soft and hard tissues. Acta Biomater. 2020, 117, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Hsiao, S.T.; Lokmic, Z.; Sivakumaran, P.; Dusting, G.J.; Dilley, R.J. Ischemic Preconditioning Promotes Intrinsic Vascularization and Enhances Survival of Implanted Cells in an In Vivo Tissue Engineering Model. Tissue Eng. Part A 2012, 18, 2210–2219. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wu, B.; Niedzielski, S.M.; Hill, M.T.; Coleman, R.M.; Ono, A.; Shikanov, A. Characterizing natural hydrogel for reconstruction of three-dimensional lymphoid stromal network to model T-cell interactions. J. Biomed. Mater. Res. Part A 2015, 103, 2701–2710. [Google Scholar] [CrossRef]
- Weisel, J.; Cederholm-Williams, S. Fibrinogen and fibrin: Characterization, processing and medical applications. In Handbook of Biodegradable Polymers; Harwood: Amsterdam, The Netherlands, 1997; pp. 347–365. [Google Scholar]
- Rubalskii, E.; Ruemke, S.; Salmoukas, C.; Aleshkin, A.; Bochkareva, S.; Modin, E.; Mashaqi, B.; Boyle, E.C.; Boethig, D.; Rubalsky, M.; et al. Fibrin glue as a local drug-delivery system for bacteriophage PA5. Sci. Rep. 2019, 9, 2091. [Google Scholar] [CrossRef]
- Wu, X.; Ren, J.; Li, J. Fibrin glue as the cell-delivery vehicle for mesenchymal stromal cells in regenerative medicine. Cytotherapy 2012, 14, 555–562. [Google Scholar] [CrossRef]
- Luo, L.; Foster, N.C.; Man, K.L.; Brunet, M.; Hoey, D.A.; Cox, S.C.; Kimber, S.J.; El Haj, A.J. Hydrostatic pressure promotes chondrogenic differentiation and microvesicle release from human embryonic and bone marrow stem cells. Biotechnol. J. 2022, 17, e2100401. [Google Scholar] [CrossRef]
- Graceffa, V. Development of a fibrin-mediated gene delivery system for the treatment of cystinosis via design of experiment. Sci. Rep. 2022, 12, 3752. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, Y.; Chen, Z.; Yue, Z.; Wallace, G. Catechol functionalized ink system and thrombin-free fibrin gel for fabricating cellular constructs with me-chanical support and inner micro channels. Biofabrication 2022, 14, 015004. [Google Scholar] [CrossRef]
- Uehlin, A.F.; Vines, J.B.; Feldman, D.S.; Nyairo, E.; Dean, D.R.; Thomas, V. Uni-Directionally Oriented Fibro-Porous PLLA/Fibrin Bio-Hybrid Scaffold: Mechano-Morphological and Cell Studies. Pharmaceutics 2022, 14, 277. [Google Scholar] [CrossRef]
- de Torre, I.G.; Weber, M.; Quintanilla, L.; Alonso, M.; Jockenhoevel, S.; Cabello, J.C.R.; Mela, P. Hybrid elastin-like recombinamer-fibrin gels: Physical characterization and in vitro evaluation for cardio-vascular tissue engineering applications. Biomater. Sci. 2016, 4, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Sawadkar, P.; Mandakhbayar, N.; Patel, K.D.; Buitrago, J.O.; Kim, T.H.; Rajasekar, P.; Lali, F.; Kyriakidis, C.; Rahmani, B.; Mohanakrishnan, J. Three dimensional porous scaffolds derived from collagen, elastin and fibrin proteins orchestrate adipose tissue regeneration. J. Tissue Eng. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.; Velz, T.; Alves, N.; Gesche, V.N.; Malischewski, A.; Schmitz-Rode, T.; Frese, J.; Jockenhoevel, S.; Mela, P. Tissue-Engineered Heart Valve with a Tubular Leaflet Design for Minimally Invasive Transcatheter Implantation. Tissue Eng. Part C Methods 2015, 21, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Lee, Y.-K.; Ran, X.; Lai, W.-H.; Li, R.A.; Keung, W.; Tse, K.; Tse, H.-F.; Yao, X. An abnormal TRPV4-related cytosolic Ca2+ rise in response to uniaxial stretch in induced pluripotent stem cells-derived cardiomyocytes from dilated cardiomyopathy patients. Biochim. Et Biophys. Acta (BBA)—Mol. Basis Dis. 2017, 1863, 2964–2972. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Jang, Y.; Kim, S.-J.; Park, Y. Cyclic Stretching Induces Maturation of Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes through Nuclear-Mechanotransduction. Tissue Eng. Regen. Med. 2022, 19, 781–792. [Google Scholar] [CrossRef]
- Yu, T.; Miyagawa, S.; Miki, K.; Saito, A.; Fukushima, S.; Higuchi, T.; Kawamura, M.; Kawamura, T.; Ito, E.; Kawaguchi, N.; et al. In Vivo Differentiation of Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Circ. J. 2013, 77, 1297–1306. [Google Scholar] [CrossRef]
- Lux, M.; Andrée, B.; Horvath, T.; Nosko, A.; Manikowski, D.; Hilfiker-Kleiner, D.; Haverich, A.; Hilfiker, A. In vitro maturation of large-scale cardiac patches based on a perfusable starter matrix by cyclic mechanical stimulation. Acta Biomater. 2016, 30, 177–187. [Google Scholar] [CrossRef]
- Kreutzer, J.; Viehrig, M.; Pölönen, R.-P.; Zhao, F.; Ojala, M.; Aalto-Setälä, K.; Kallio, P. Pneumatic unidirectional cell stretching device for mechanobiological studies of cardiomyocytes. Biomech. Model. Mechanobiol. 2020, 19, 291–303. [Google Scholar] [CrossRef]
- Dou, W.; Malhi, M.; Cui, T.; Wang, M.; Wang, T.; Shan, G.; Law, J.; Gong, Z.; Plakhotnik, J.; Filleter, T. A Carbon-Based Biosensing Platform for Simultaneously Measuring the Contraction and Electrophysiology of iPSC-Cardiomyocyte Monolayers. ACS Nano 2022, 16, 11278–11290. [Google Scholar] [CrossRef]
- Kroll, K.; Chabria, M.; Wang, K.; Häusermann, F.; Schuler, F.; Polonchuk, L. Electro-mechanical conditioning of human iPSC-derived cardiomyocytes for translational research. Prog. Biophys. Mol. Biol. 2017, 130, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Burnham, M.P.; Harvey, R.; Sargeant, R.; Fertig, N.; Haddrick, M. A Scalable Approach Reveals Functional Responses of iPSC Cardiomyocyte 3D Spheroids. SLAS Discov. Adv. Sci. Drug Discov. 2021, 26, 352–363. [Google Scholar] [CrossRef] [PubMed]
- LaBarge, W.; Mattappally, S.; Kannappan, R.; Fast, V.G.; Pretorius, D.; Berry, J.L.; Zhang, J. Maturation of three-dimensional, hiPSC-derived cardiomyocyte spheroids utilizing cyclic, uniaxial stretch and electrical stimulation. PLoS ONE 2019, 14, e0219442. [Google Scholar]
- Yoshida, S.; Sumomozawa, K.; Nagamine, K.; Nishizawa, M. Hydrogel microchambers integrated with organic electrodes for efficient electrical stimulation of human iPSC-derived cardiomyocytes. Macromol. Biosci. 2019, 19, 1900060. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Fujisawa, K.; Yukawa, Y.; Matsunaga, Y.T. Bottom-up fabrication of artery-mimicking tubular co-cultures in collagen-based microchannel scaffolds. Biomater. Sci. 2016, 4, 1503–1514. [Google Scholar] [CrossRef]
- Schimek, K.; Busek, M.; Brincker, S.; Groth, B.; Hoffmann, S.; Lauster, R.; Lindner, G.; Lorenz, A.; Menzel, U.; Sonntag, F.; et al. Integrating biological vasculature into a multi-organ-chip microsystem. Lab Chip 2013, 13, 3588–3598. [Google Scholar] [CrossRef]
- Heywood, H.K.; Bader, D.L.; Lee, D.A. Rate of oxygen consumption by isolated articular chondrocytes is sensitive to medium glucose concentration. J. Cell. Physiol. 2006, 206, 402–410. [Google Scholar] [CrossRef]
- Ząbczyk, M.; Natorska, J.; Janion-Sadowska, A.; Malinowski, K.P.; Janion, M.; Undas, A. Elevated Lactate Levels in Acute Pulmonary Embolism Are Associated with Prothrombotic Fibrin Clot Properties: Contribution of NETs Formation. J. Clin. Med. 2020, 9, 953. [Google Scholar] [CrossRef]
- Zheng, D.-W.; Hong, S.; Zhang, Q.-L.; Dong, X.; Pan, P.; Song, W.-F.; Cheng, S.-X.; Zhang, X.-Z. Controllable gelation of artificial extracellular matrix for altering mass transport and improving cancer therapies. Nat. Commun. 2020, 11, 4907. [Google Scholar] [CrossRef]
- Flanagan, T.C.; Cornelissen, C.; Koch, S.; Tschoeke, B.; Sachweh, J.S.; Schmitzrode, T.; Jockenhoevel, S. The in vitro development of autologous fibrin-based tissue-engineered heart valves through optimised dynamic conditioning. Biomaterials 2007, 28, 3388–3397. [Google Scholar] [CrossRef]
- Necela, B.M.; Velz, T.; Alves, N.; Gesche, V.N.; Malischewski, A.; Schmitz-Rode, T.; Frese, J.; Jockenhoevel, S.; Mela, P. The antineoplastic drug, trastuzumab, dysregulates metabolism in iPSC-derived cardiomyocytes. Clin. Transl. Med. 2017, 6, 5. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
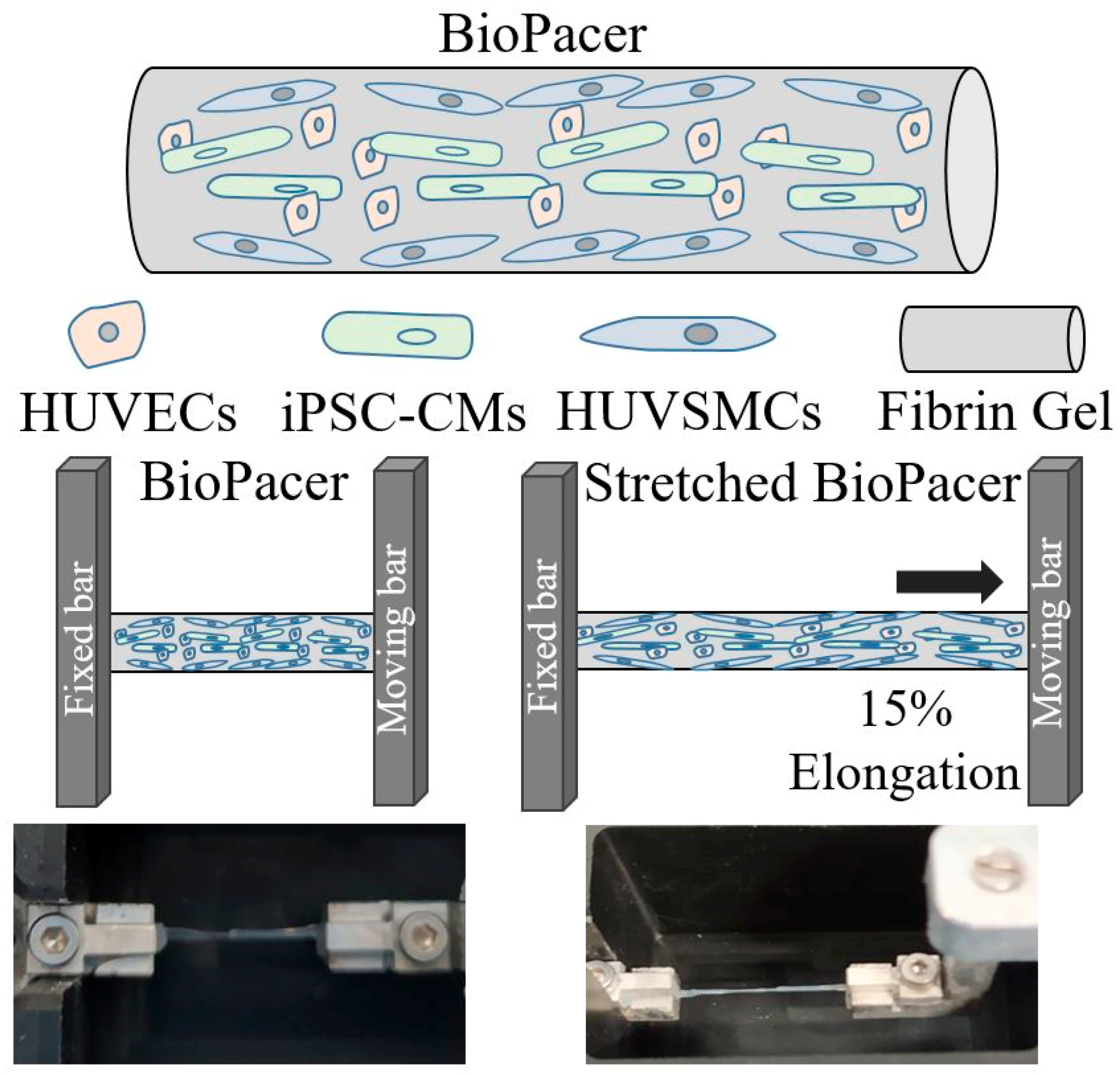{kind=link}
| Sample Type | Fiber Number | [iPSC-CMs] (Cells/Fiber) | [HUVSMCs] (Cells/Fiber) | [HUVECs] (Cells/Fiber) |
|---|---|---|---|---|
| Low cell density 500 µm (G1) | 2 | 150,000 | 60,000 | 20,000 |
| Mix 500 µm (G2) | 1 | 300,000 | 60,000 | 20,000 |
| 500 µm (G3) | 2 | 300,000 | 60,000 | 20,000 |
| 500 µm stretch (G4) | 2 | 300,000 | 60,000 | 20,000 |
| 2 × 500 µm (G5) | 2 | 300,000 | ||
| 3 × 500 µm (G6) | 3 | 300,000 | ||
| 1 mm (G7) | 2 | 500,000 | 120,000 | 60,000 |
| 1 mm stretch (G8) | 2 | 500,000 | 120,000 | 60,000 |
| Mix 1 mm (G9) | 1 | 500,000 | 120,000 | 60,000 |
| Antibody | Dilution | Supply |
|---|---|---|
| Anti-α-Actinin (sarcomeric), mouse monoclonal | 1:50 | Sigma-Aldrich |
| Connexin 43 (polyclonal) | 1:50 | Thermofischer |
| Connexin 43 Monoclonal Antibody (3D8A5) | 1:50 | Thermofischer |
| Anti-CD31 (PECAM-1), mouse monoclonal | 1:100 | Sigma-Aldrich |
| CD31 rabbit polyclonal | 1:100 | Abbiotec |
| α-smooth muscle actin (monoclonal, clone 1A4) | 1:1000 | Sigma-Aldrich |
| Anti-Collagen I antibody (ab34710) | 1:200 | Abcam |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriakou, S.; Lubig, A.; Sandhoff, C.A.; Kuhn, Y.; Jockenhoevel, S. Influence of Diameter and Cyclic Mechanical Stimulation on the Beating Frequency of Myocardial Cell-Laden Fibers. Gels 2023, 9, 677. https://doi.org/10.3390/gels9090677
Kyriakou S, Lubig A, Sandhoff CA, Kuhn Y, Jockenhoevel S. Influence of Diameter and Cyclic Mechanical Stimulation on the Beating Frequency of Myocardial Cell-Laden Fibers. Gels. 2023; 9(9):677. https://doi.org/10.3390/gels9090677
Chicago/Turabian StyleKyriakou, Stavroula, Andreas Lubig, Cilia A. Sandhoff, Yasmin Kuhn, and Stefan Jockenhoevel. 2023. "Influence of Diameter and Cyclic Mechanical Stimulation on the Beating Frequency of Myocardial Cell-Laden Fibers" Gels 9, no. 9: 677. https://doi.org/10.3390/gels9090677
{kind=link}