
A Hydrogel as a Bespoke Delivery Platform for Stromal Cell-Derived Factor-1

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
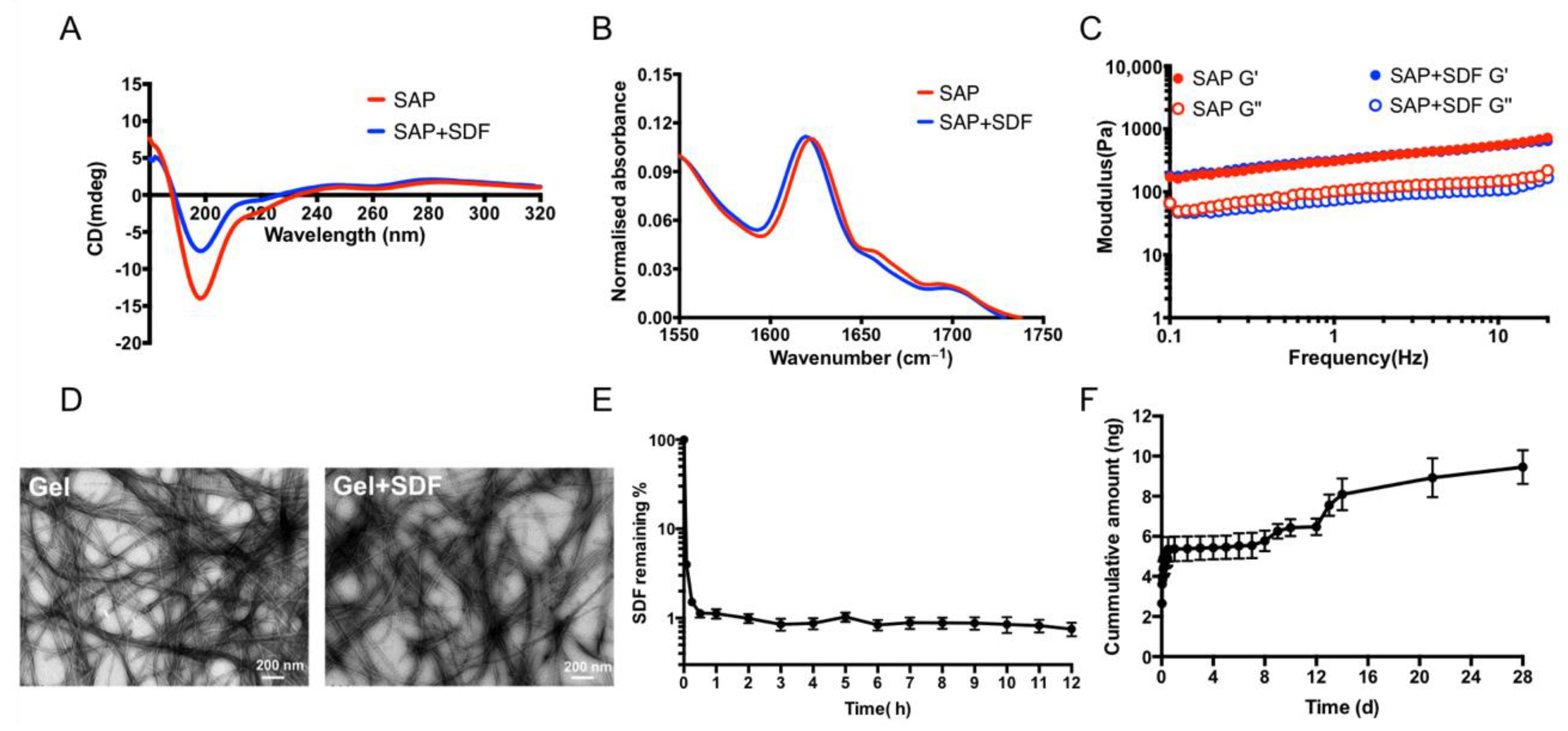
2.1.1. Homeostatic Hydrogels Enable the Sustained Release of SDF
2.1.2. Homeostatic Hydrogels Are Biocompatible In Vivo
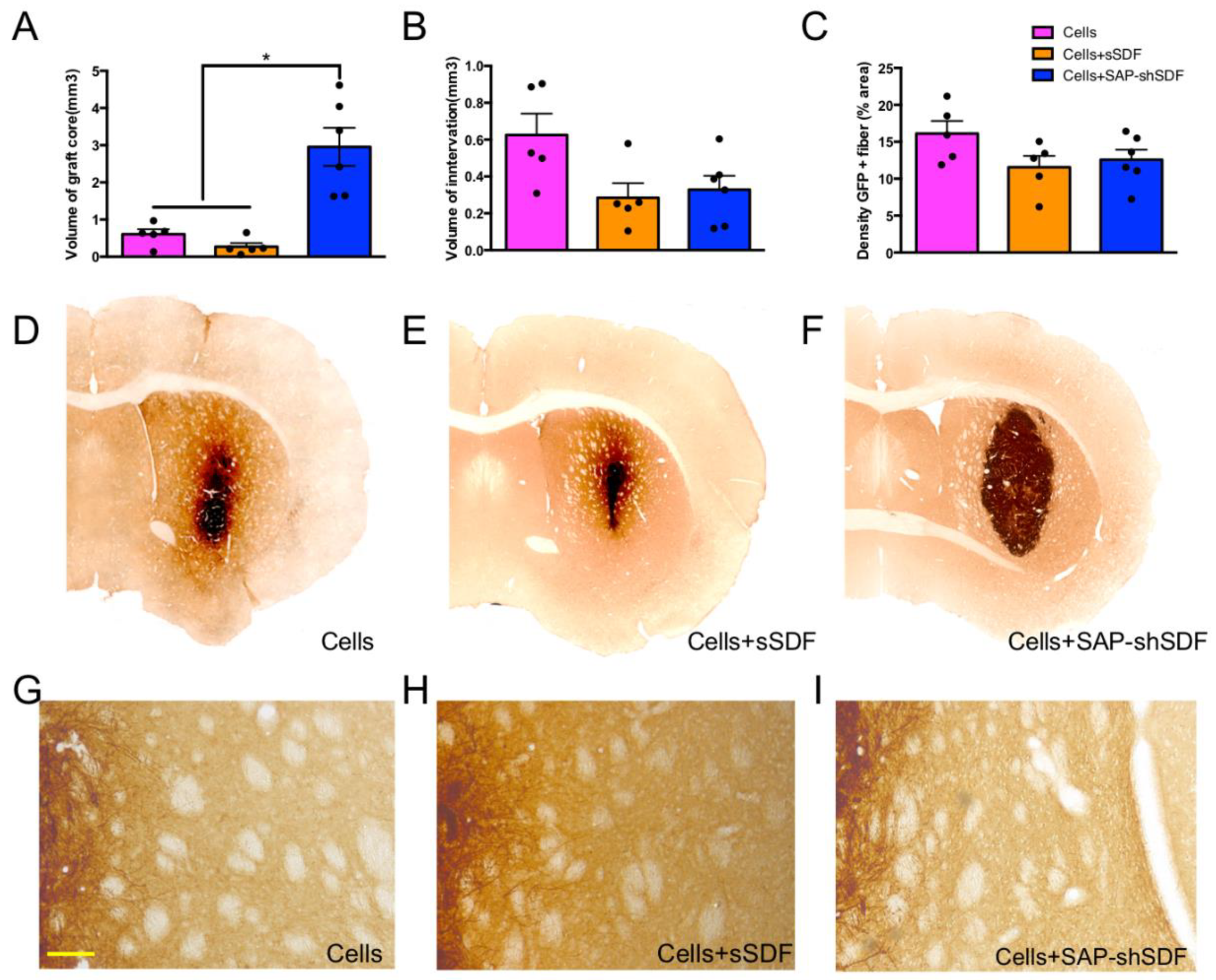
2.1.3. Homeostatic Hydrogels Support Graft Growth
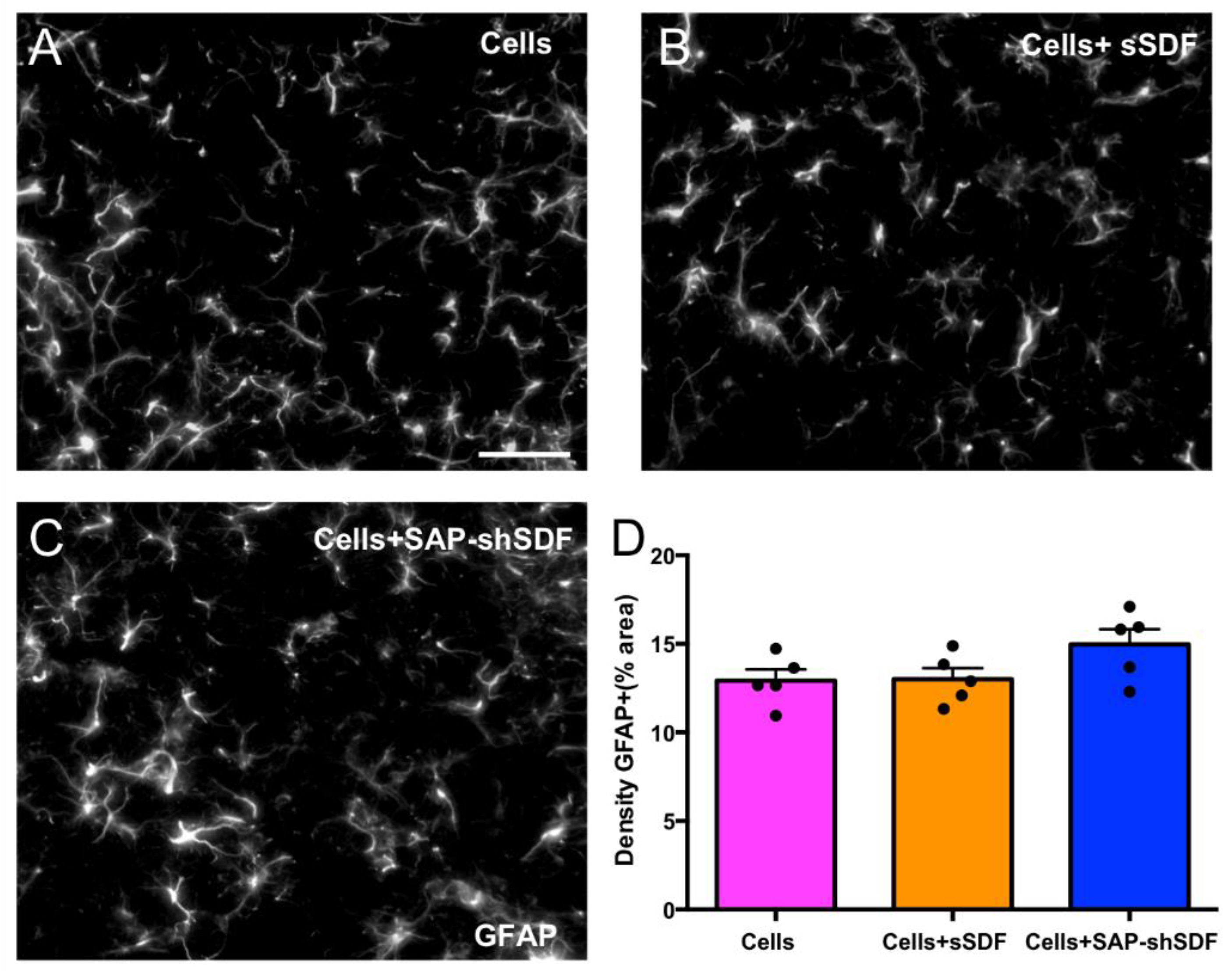
2.1.4. Homeostatic Hydrogels Promote Differentiation of Cell Graft
2.2. Discussion
3. Conclusions
4. Materials and Methods
4.1. Solid-Phase Peptide Synthesis
4.2. Preparation of SAP Hydrogel
4.3. Circular Dichroism (CD)
4.4. Fourier Transform Infrared Spectroscopy (FTIR)
4.5. Rheology
4.6. Transmission Electron Microscopy (TEM)
4.7. Release Profiles and Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. In Vivo Transplantation of Cells with SDF-1 and Tissue Processing
4.9. Immunohistochemistry
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, R.A.; Götz, M.; Parmar, M. New approaches for brain repair—From rescue to reprogramming. Nature 2018, 557, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P.; Kim, B.Y.; Trounson, A. How to design preclinical studies in nanomedicine and cell therapy to maximize the prospects of clinical translation. Nat. Biomed. Eng. 2018, 2, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.; Kirik, D.; Björklund, A. Cell transplantation in Parkinson’s disease: How can we make it work? Trends Neurosci. 2005, 28, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Nisbet, D.R.; Pattanawong, S.; Ritchie, N.E.; Shen, W.; Finkelstein, D.I.; Horne, M.K.; Forsythe, J.S. Interaction of embryonic cortical neurons on nanofibrous scaffolds for neural tissue engineering. J. Neural Eng. 2007, 4, 35–41. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, T.; Zhang, X.-C.; Nagasawa, T.; Wu, J.Y.; Rao, Y. Role of the chemokine SDF-1 as the meningeal attractant for embryonic cerebellar neurons. Nat. Neurosci. 2002, 5, 719–720. [Google Scholar] [CrossRef] [PubMed]
- Lazarini, F.; Tham, T.N.; Casanova, P.; Arenzana-Seisdedos, F.; Dubois-Dalcq, M. Role of the α-chemokine stromal cell-derived factor (SDF-1) in the developing and mature central nervous system. Glia 2003, 42, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chang, S.; Li, W.; Tang, G.; Ma, Y.; Liu, Y.; Yuan, F.; Zhang, Z.; Yang, G.-Y.; Wang, Y. cxcl12-engineered endothelial progenitor cells enhance neurogenesis and angiogenesis after ischemic brain injury in mice. Stem Cell Res. Ther. 2018, 9, 139. [Google Scholar] [CrossRef]
- Zhu, C.; Mahesula, S.; Temple, S.; Kokovay, E. Heterogeneous expression of SDF1 retains actively proliferating neural progenitors in the capillary compartment of the niche. Stem Cell Rep. 2019, 12, 6–13. [Google Scholar] [CrossRef] [Green Version]
- O’Dwyer, J.; Cullen, M.; Fattah, S.; Murphy, R.; Stefanovic, S.; Kovarova, L.; Pravda, M.; Velebny, V.; Heise, A.; Duffy, G.P. Development of a sustained release nano-in-gel delivery system for the chemotactic and angiogenic growth factor stromal-derived factor 1α. Pharmaceutics 2020, 12, 513. [Google Scholar] [CrossRef]
- Lataillade, J.-J.; Clay, D.; Dupuy, C.; Rigal, S.; Jasmin, C.; Bourin, P.; Bousse-Kerdilès, M.-C.L. Chemokine SDF-1 enhances circulating CD34+ cell proliferation in synergy with cytokines: Possible role in progenitor survival. Blood 2000, 95, 756–768. [Google Scholar] [CrossRef]
- Burns, J.M.; Summers, B.C.; Wang, Y.; Melikian, A.; Berahovich, R.; Miao, Z.; Penfold, M.E.; Sunshine, M.J.; Littman, D.R.; Kuo, C.J. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J. Exp. Med. 2006, 203, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Somaa, F.A.; Bye, C.R.; Thompson, L.H.; Parish, C.L. Meningeal cells influence midbrain development and the engraftment of dopamine progenitors in Parkinsonian mice. Exp. Neurol. 2015, 267, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.; Tokunou, T.; Higgins, L.J.; MacGillivray, C.; Gannon, J.; Lee, R.T. Local delivery of protease-resistant stromal cell derived factor-1 for stem cell recruitment after myocardial infarction. Circulation 2007, 116, 1683–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromage, D.I.; Davidson, S.M.; Yellon, D.M. Stromal derived factor 1α: A chemokine that delivers a two-pronged defence of the myocardium. Pharmacol. Ther. 2014, 143, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, K.F.; Rodriguez, A.L.; Parish, C.L.; Williams, R.J.; Nisbet, D.R. Temporally controlled release of multiple growth factors from a self-assembling peptide hydrogel. Nanotechnology 2016, 27, 385102. [Google Scholar] [CrossRef]
- Hunt, C.P.; Penna, V.; Gantner, C.W.; Moriarty, N.; Wang, Y.; Franks, S.; Ermine, C.M.; de Luzy, I.R.; Pavan, C.; Long, B.M. Tissue programmed hydrogels functionalized with GDNF improve human neural grafts in Parkinson’s disease. Adv. Funct. Mater. 2021, 31, 2105301. [Google Scholar] [CrossRef]
- Nisbet, D.; Wang, T.; Bruggeman, K.; Niclis, J.; Somaa, F.; Penna, V.; Hunt, C.; Wang, Y.; Kauhausen, J.; Williams, R.J. Shear containment of BDNF within molecular hydrogels promotes human stem cell engraftment and postinfarction remodeling in stroke. Adv. Biosyst. 2018, 2, 1800113. [Google Scholar] [CrossRef]
- Campbell, E.C.; Grant, J.; Wang, Y.; Sandhu, M.; Williams, R.J.; Nisbet, D.R.; Perriman, A.W.; Lupton, D.W.; Jackson, C.J. Hydrogel-immobilized supercharged proteins. Adv. Biosyst. 2018, 2, 1700240. [Google Scholar] [CrossRef]
- Wang, Y.; Bruggeman, K.F.; Franks, S.; Gautam, V.; Hodgetts, S.I.; Harvey, A.R.; Williams, R.J.; Nisbet, D.R. Is viral vector gene delivery more effective using biomaterials? Adv. Healthc. Mater. 2021, 10, 2001238. [Google Scholar] [CrossRef]
- Rodriguez, A.L.; Wang, T.-Y.; Bruggeman, K.F.; Li, R.; Williams, R.J.; Parish, C.L.; Nisbet, D.R. Tailoring minimalist self-assembling peptides for localized viral vector gene delivery. Nano Res. 2016, 9, 674–684. [Google Scholar] [CrossRef]
- Franks, S.J.; Firipis, K.; Ferreira, R.; Hannan, K.M.; Williams, R.J.; Hannan, R.D.; Nisbet, D.R. Harnessing the self-assembly of peptides for the targeted delivery of anti-cancer agents. Mater. Horiz. 2020, 7, 1996–2010. [Google Scholar] [CrossRef]
- Bruggeman, K.; Moriarty, N.; Dowd, E.; Nisbet, D.; Parish, C. Harnessing stem cells and biomaterials to promote neural repair. Br. J. Pharmacol. 2019, 176, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Maclean, F.L.; Ims, G.M.; Horne, M.K.; Williams, R.J.; Nisbet, D.R. A programmed anti-Inflammatory nanoscaffold (PAIN) as a 3D tool to understand the brain injury response. Adv. Mater. 2018, 30, 1805209. [Google Scholar] [CrossRef]
- Rodriguez, A.; Wang, T.-Y.; Bruggeman, K.; Horgan, C.; Li, R.; Williams, R.J.; Parish, C.L.; Nisbet, D. In vivo assessment of grafted cortical neural progenitor cells and host response to functionalized self-assembling peptide hydrogels and the implications for tissue repair. J. Mater. Chem. B 2014, 2, 7771–7778. [Google Scholar] [CrossRef]
- Somaa, F.A.; Wang, T.-Y.; Niclis, J.C.; Bruggeman, K.F.; Kauhausen, J.A.; Guo, H.; McDougall, S.; Williams, R.J.; Nisbet, D.R.; Thompson, L.H. Peptide-based scaffolds support human cortical progenitor graft integration to reduce atrophy and promote functional repair in a model of stroke. Cell Rep. 2017, 20, 1964–1977. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; He, X.; Bruggeman, K.F.; Gayen, B.; Tricoli, A.; Lee, W.M.; Williams, R.J.; Nisbet, D.R. Peptide programmed hydrogels as safe sanctuary microenvironments for cell transplantation. Adv. Funct. Mater. 2020, 30, 1900390. [Google Scholar] [CrossRef]
- Cembran, A.; Bruggeman, K.F.; Williams, R.J.; Parish, C.L.; Nisbet, D.R. Biomimetic materials and their utility in modeling the 3-dimensional neural environment. Iscience 2020, 23, 100788. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Bruggeman, K.; Wang, Y.; Wang, T.-Y.; Williams, R.J.; Parish, C.L.; Nisbet, D. Using minimalist self-assembling peptides as hierarchical scaffolds to stabilise growth factors and promote stem cell integration in the injured brain. J. Tissue Eng. Regener. Med. 2018, 12, e1571–e1579. [Google Scholar] [CrossRef]
- Sur, S.; Pashuck, E.T.; Guler, M.O.; Ito, M.; Stupp, S.I.; Launey, T. A hybrid nanofiber matrix to control the survival and maturation of brain neurons. Biomaterials 2012, 33, 545–555. [Google Scholar] [CrossRef]
- Matson, J.B.; Zha, R.H.; Stupp, S.I. Peptide self-assembly for crafting functional biological materials. Curr. Opin. Solid State Mater. Sci. 2011, 15, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Aye, S.-S.S.; Li, R.; Boyd-Moss, M.; Long, B.; Pavuluri, S.; Bruggeman, K.; Wang, Y.; Barrow, C.R.; Nisbet, D.R.; Williams, R.J. Scaffolds formed via the non-equilibrium supramolecular assembly of the synergistic ECM peptides RGD and PHSRN demonstrate improved cell attachment in 3D. Polymers 2018, 10, 690. [Google Scholar] [CrossRef] [Green Version]
- Dityatev, A.; Schachner, M. Extracellular matrix molecules and synaptic plasticity. Nat. Rev. Neurosci. 2003, 4, 456–468. [Google Scholar] [CrossRef]
- Aguado, B.A.; Mulyasasmita, W.; Su, J.; Lampe, K.J.; Heilshorn, S.C. Improving viability of stem cells during syringe needle flow through the design of hydrogel cell carriers. Tissue Eng. Part A 2012, 18, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.; Ko, M.K.; Jin, Y.; Shin, W.J.; Park, W.; Chae, S.; Lee, H.J.; Jang, J.; Yi, H.-G.; Lee, D.S. Neural stem cell delivery using brain-derived tissue-specific bioink for recovering from traumatic brain injury. Biofabrication 2021, 13, 44110. [Google Scholar] [CrossRef]
- Gantner, C.W.; de Luzy, I.R.; Kauhausen, J.A.; Moriarty, N.; Niclis, J.C.; Bye, C.R.; Penna, V.; Hunt, C.P.J.; Ermine, C.M.; Pouton, C.W.; et al. Viral delivery of GDNF promotes functional integration of human stem cell grafts in Parkinson’s disease. Cell Stem Cell 2020, 26, 511–526. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, H.; Zhang, X.; Zhao, S.; Zhou, Z.; Mu, X.; Zhao, C.; Teng, W. The Role of SDF-1/CXCR4/CXCR7 in Neuronal Regeneration after Cerebral Ischemia. Front. Neurosci. 2017, 11, 590. [Google Scholar] [CrossRef]
- Tsui, C.; Koss, K.; Churchward, M.A.; Todd, K.G. Biomaterials and glia: Progress on designs to modulate neuroinflammation. Acta Biomater. 2019, 83, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chang, C.J.; Lathia, J.D.; Wang, L.; Pacenta, H.L.; Cotleur, A.; Ransohoff, R.M. Chemokine receptor CXCR4 signaling modulates the growth factor-induced cell cycle of self-renewing and multipotent neural progenitor cells. Glia 2011, 59, 108–118. [Google Scholar] [CrossRef]
- Khan, M.Z.; Brandimarti, R.; Shimizu, S.; Nicolai, J.; Crowe, E.; Meucci, O. The chemokine CXCL12 promotes survival of postmitotic neurons by regulating Rb protein. Cell Death Differ. 2008, 15, 1663–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wei, Y.; Liu, J.; Zhang, H. Chemotactic responses of neural stem cells to SDF-1α correlate closely with their differentiation status. J. Mol. Neurosci. 2014, 54, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-Y.; Bruggeman, K.F.; Kauhausen, J.A.; Rodriguez, A.L.; Nisbet, D.R.; Parish, C.L. Functionalized composite scaffolds improve the engraftment of transplanted dopaminergic progenitors in a mouse model of Parkinson’s disease. Biomaterials 2016, 74, 89–98. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Penna, V.; Williams, R.J.; Parish, C.L.; Nisbet, D.R. A Hydrogel as a Bespoke Delivery Platform for Stromal Cell-Derived Factor-1. Gels 2022, 8, 224. https://doi.org/10.3390/gels8040224
Wang Y, Penna V, Williams RJ, Parish CL, Nisbet DR. A Hydrogel as a Bespoke Delivery Platform for Stromal Cell-Derived Factor-1. Gels. 2022; 8(4):224. https://doi.org/10.3390/gels8040224
Chicago/Turabian StyleWang, Yi, Vanessa Penna, Richard J. Williams, Clare L. Parish, and David R. Nisbet. 2022. "A Hydrogel as a Bespoke Delivery Platform for Stromal Cell-Derived Factor-1" Gels 8, no. 4: 224. https://doi.org/10.3390/gels8040224