
Secretion of Bacillus amyloliquefaciens Transglutaminase from Lactococcus lactis and Its Enhancement of Food Gel Properties
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Results Analysis
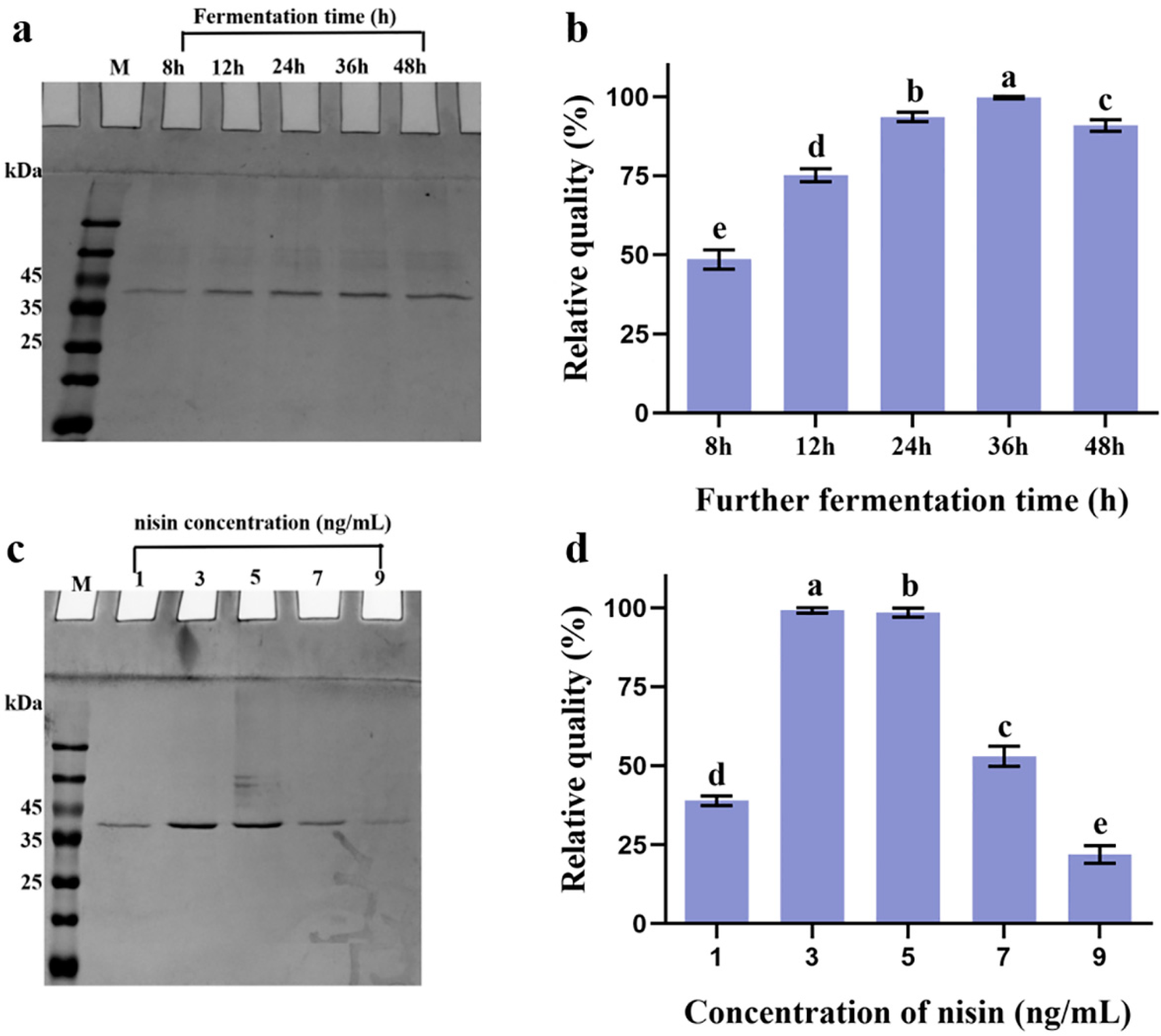
2.1.1. Secretion of Heterologous BaMTGase in L. lactis NZ9000
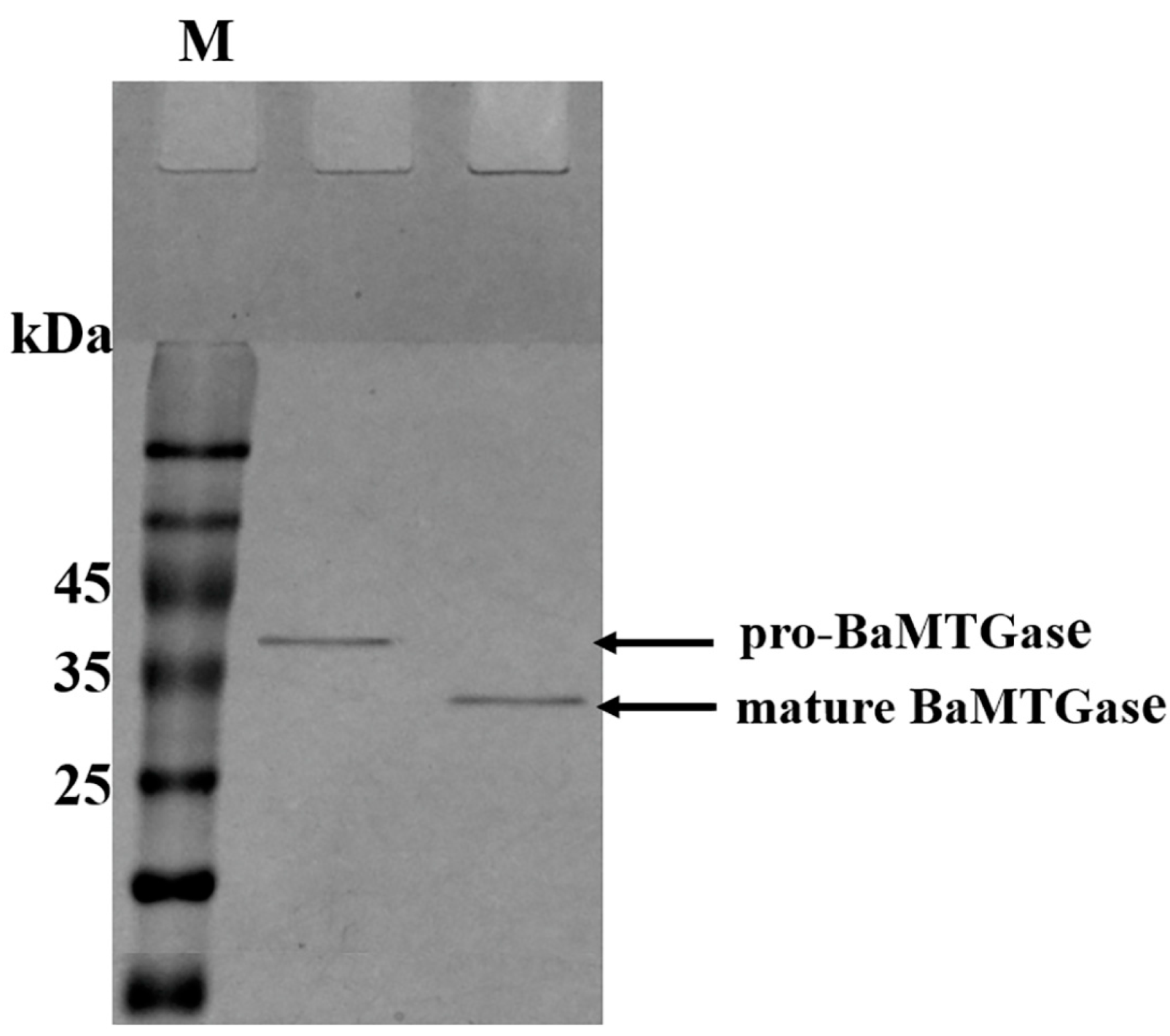
2.1.2. Activation of Pro-BaMTGase and Determination of Enzyme Activity
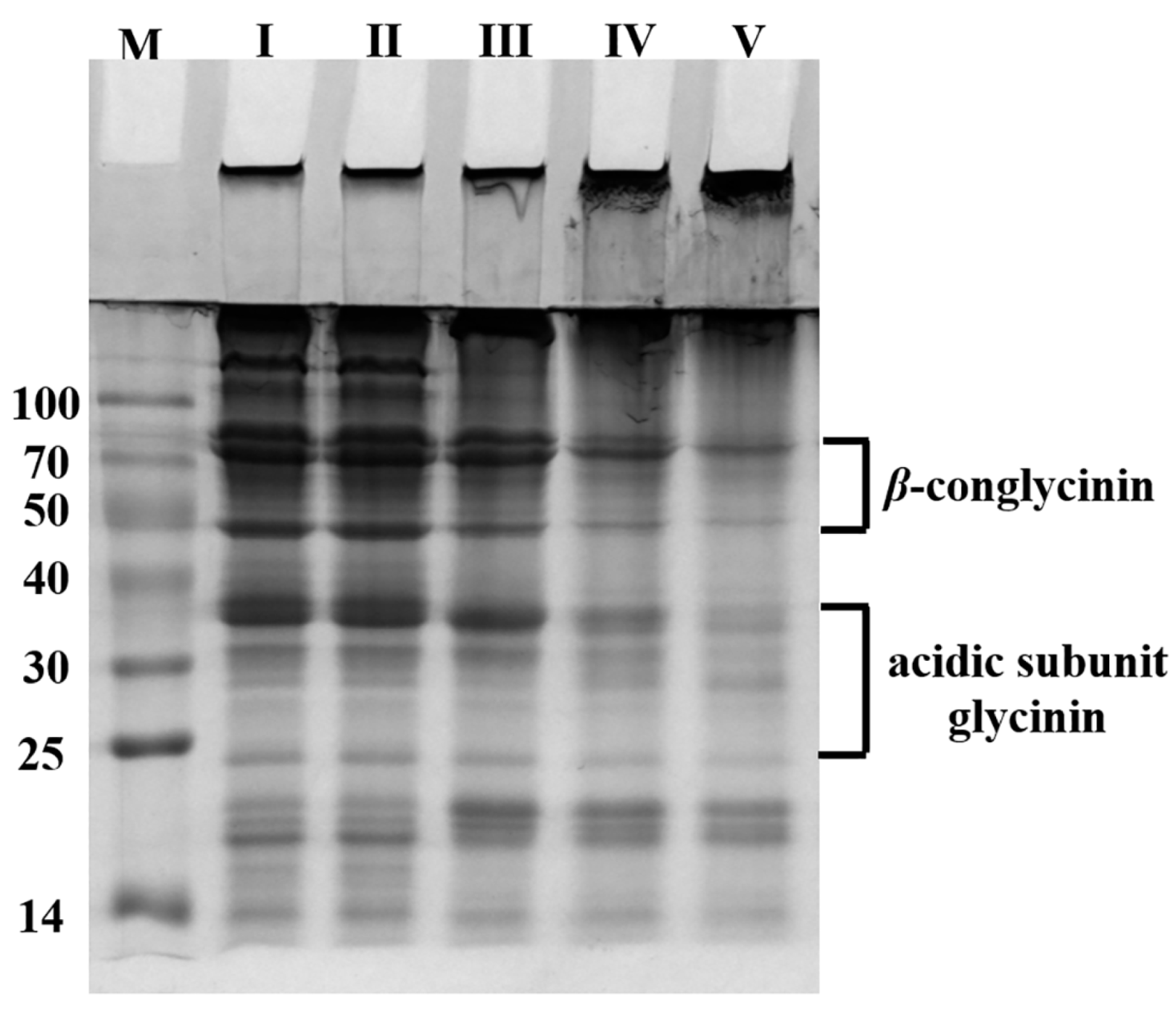
2.1.3. Crosslinking of 1% (w/v) SPI
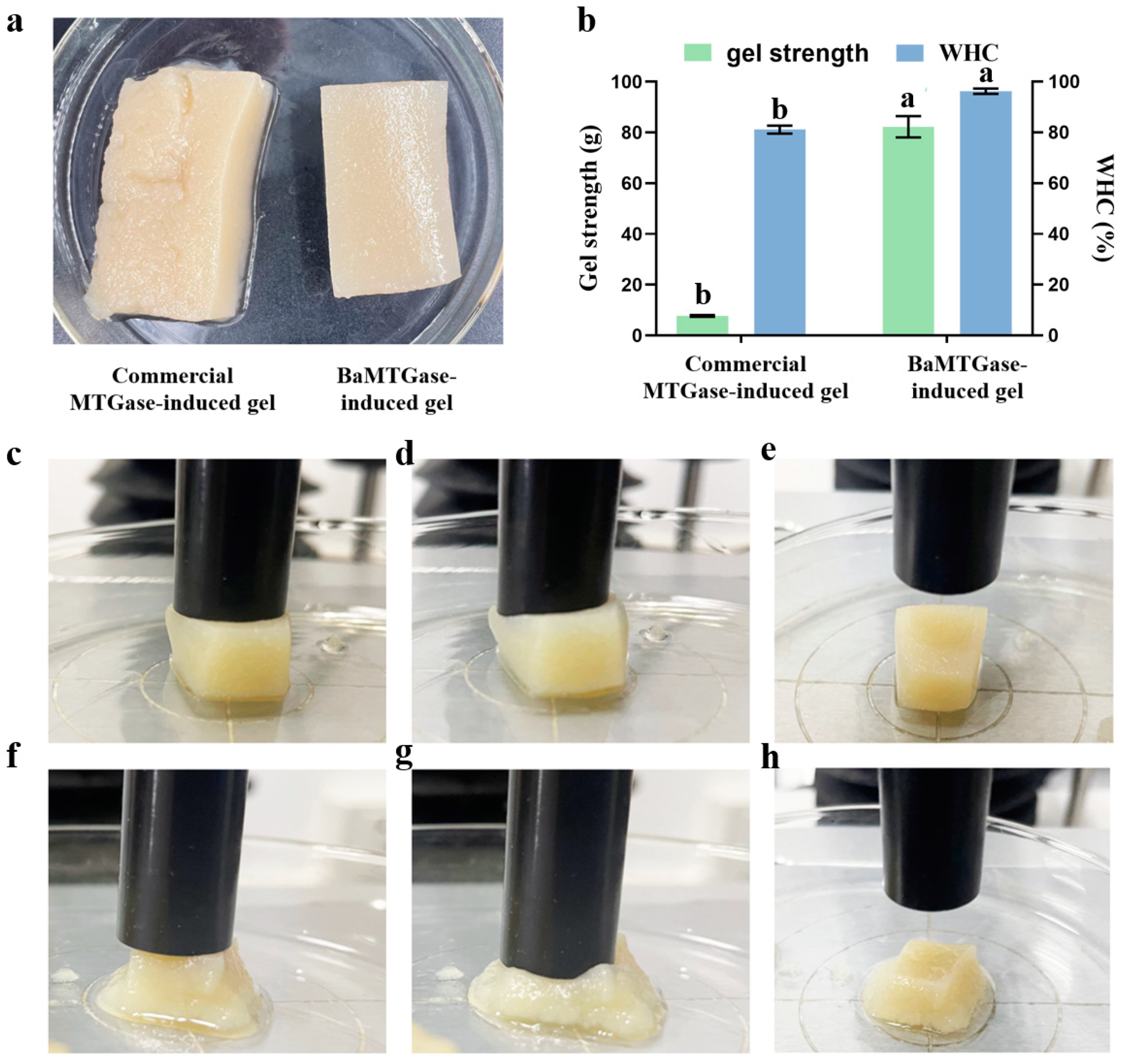
2.1.4. Gel Strength and Water-Holding Capacity
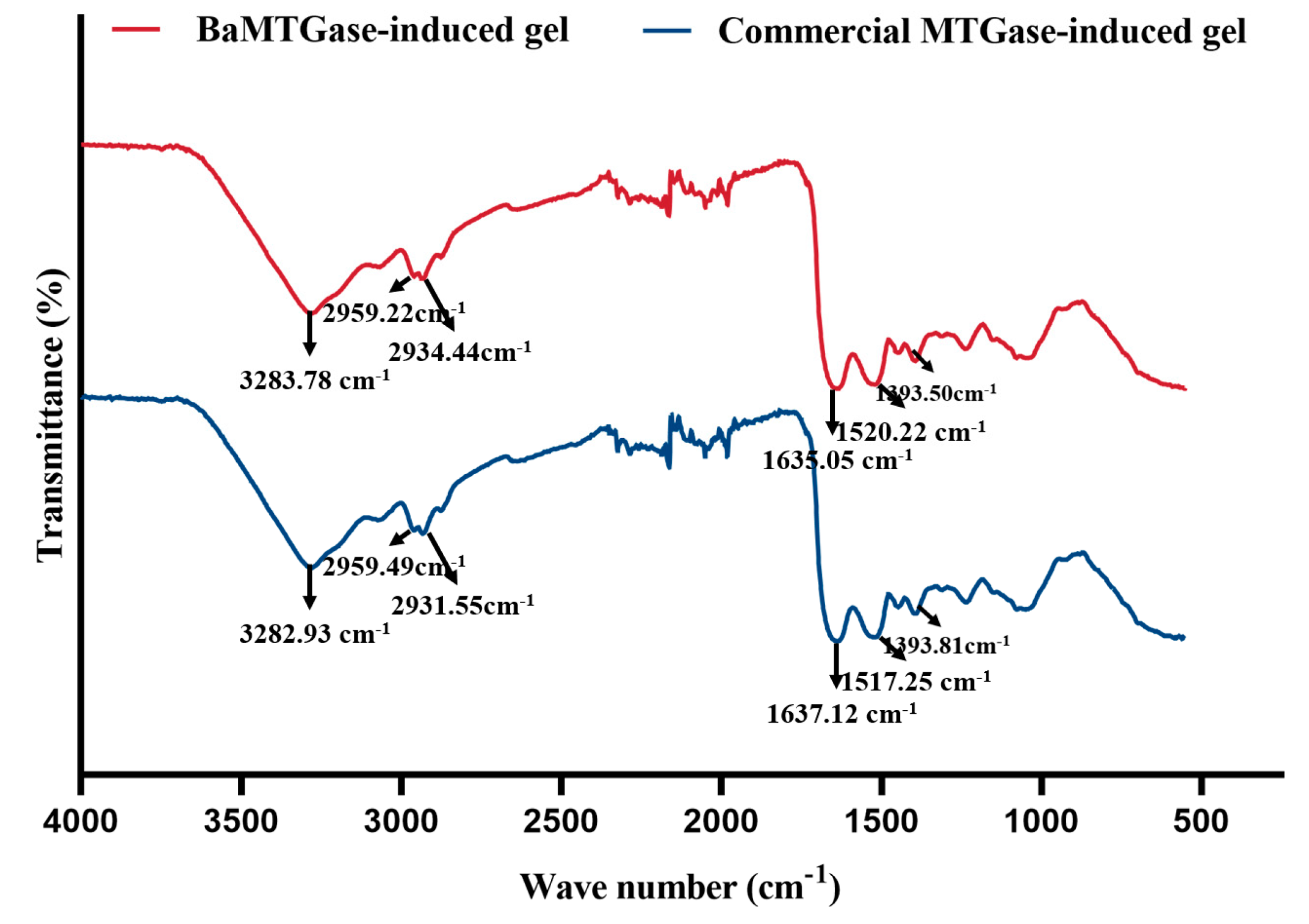
2.1.5. FT-IR Analysis
2.1.6. Microstructure of the Gel
2.2. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of MTGase | Expression Host Strain | Activity | Ref |
|---|---|---|---|
| Streptomyces mobaraensis | Escherichia coli | 23 U/mg | [30] |
| Streptomyces fradiae | Pichia pastoris strain GS115 | 0.7 U/mL | [31] |
| Streptomyces mobaraensis | Bacillus subtilis | 16.1 U/mg | [32] |
| Streptomyces netropsis | Escherichia coli | 18.2 U/mg | [33] |
| B. amyloliquefaciens DSM7 | Escherichia coli | 37 mU/mg | [34] |
| Streptomyces hygroscopicus WSH03-13 | Yarrowia lipolytica | 5.3 U/mL | [35] |
| Streptomyces mobaraensis | Bacillus subtilis | 29.6 U/mg | [14] |
| Streptomyces mobaraensis | Lactococcus lactis NZ9000 | 27.6 U/mg | [15] |
3. Conclusions
4. Materials and Methods
4.1. Materials, Bacterial Strains and Growth Conditions
4.2. Construction of Transformants
4.3. Recombinant BaMTGase Overexpression and the Optimization of Secretion Conditions in L. lactis NZ9000
4.4. Protein Purification and Determination of Enzyme Activity
4.5. SPI Gelation Crosslinked by MTGase (SDS-PAGE)
4.6. Preparation of SPI Gels with MTGase
4.7. Determination of SPI Gel Texture
4.8. Water-Holding Capacity Determination
4.9. Fourier Transform Infrared Spectroscope (FT-IR) Analysis
4.10. Characterization of SPI Gel Microstructure
4.11. Protein Band Analysis and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mattice, K.D.; Marangoni, A.G. Physical properties of zein networks treated with microbial transglutaminase. Food Chem. 2021, 338, 128010. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Wang, G.; Gao, D.; Wang, L.; Zhang, A.; Wang, X.; Xu, N.; Jiang, L. Improving the gel properties of transgenic microbial transglutaminase cross-linked soybean-whey mixed protein by ultrasonic pretreatment. Process. Biochem. 2020, 91, 104–112. [Google Scholar] [CrossRef]
- Damodaran, S.; Agyare, K.K. Effect of microbial transglutaminase treatment on thermal stability and pH-solubility of heat-shocked whey protein isolate. Food Hydrocoll. 2013, 30, 12–18. [Google Scholar] [CrossRef]
- Wang, L.; Yu, B.; Wang, R.; Xie, J. Biotechnological routes for transglutaminase production: Recent achievements, perspectives and limits. Trends Food Sci. Technol. 2018, 81, 116–120. [Google Scholar] [CrossRef]
- Gaspar, A.L.C.; de Góes-Favoni, S.P. Action of microbial transglutaminase (MTGase) in the modification of food proteins: A review. Food Chem. 2015, 171, 315–322. [Google Scholar] [CrossRef]
- Tokay, F.G.; Alp, A.C.; Yerlikaya, P. Production and shelf life of restructured fish meat binded by microbial transglutaminase. LWT-Food Sci. Technol. 2021, 152, 112369. [Google Scholar] [CrossRef]
- Aalami, M.; Leelavathi, K. Effect of microbial transglutaminase on spaghetti quality. J. Food Sci. 2008, 73, C306–C312. [Google Scholar] [CrossRef]
- Kuraishi, C.; Yamazaki, K.; Susa, Y. Transglutaminase: Its utilization in the food industry. Food Rev. Int. 2007, 17, 221–246. [Google Scholar] [CrossRef]
- Akbari, M.; Razavi, S.H.; Kieliszek, M. Recent advances in microbial transglutaminase biosynthesis and its application in the food industry. Trends Food Sci. Technol. 2021, 110, 458–469. [Google Scholar] [CrossRef]
- Ogilvie, O.; Roberts, S.; Sutton, K.; Larsen, N.; Gerrard, J.; Domigan, L. The use of microbial transglutaminase in a bread system: A study of gluten protein structure, deamidation state and protein digestion. Food Chem. 2021, 340, 127903. [Google Scholar] [CrossRef]
- Ando, H.; Adachi, M.; Umeda, K.; Matsuura, A.; Nonaka, M. Purification and Characteristics of a Novel Transglutaminase Derived from Microorganisms. Agric. Biol. Chem. 1989, 53, 2613–2617. [Google Scholar]
- Kieliszek, M.; Misiewicz, A. Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. 2014, 59, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Sorde, K.L.; Ananthanarayan, L. Isolation, screening, and optimization of bacterial strains for novel transglutaminase production. Prep. Biochem. Biotechnol. 2019, 49, 64–73. [Google Scholar] [CrossRef]
- Mu, D.; Lu, J.; Qiao, M.; Kuipers, O.P.; Zhu, J.; Li, X.; Yang, P.; Zhao, Y.; Luo, S.; Wu, X.; et al. Heterologous signal peptides-directing secretion of Streptomyces mobaraensis transglutaminase by Bacillus subtilis. Appl. Microbiol. Biotechnol. 2018, 102, 5533–5543. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Lu, J.; Zhu, J.; Li, X.; Gu, H.; Montalbán-López, M.; Wu, X.; Luo, S.; Zhao, Y.; Jiang, S.; et al. The Secretion of Streptomyces monbaraensis Transglutaminase from Lactococcus lactis and Immobilization on Porous Magnetic Nanoparticles. Front. Microbiol. 2019, 10, 1675. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yang, Z.; Li, Z.; Tian, Y. Heterologous Expression of Recombinant Transglutaminase in Bacillus subtilis SCK6 with Optimized Signal Peptide and Codon, and Its Impact on Gelatin Properties. J. Microbiol. Biotechnol. 2020, 30, 1082–1091. [Google Scholar] [CrossRef]
- Neef, J.; van Dijl, J.M.; Buist, G. Recombinant protein secretion by Bacillus subtilis and Lactococcus lactis: Pathways, applications, and innovation potential. Essays Biochem. 2021, 65, 187–195. [Google Scholar]
- Ma, T.; Li, X.; Wu, X.; Jiang, S.; Zheng, Z.; Mu, D. Expression of Bacillus amyloliquefaciens γ-Glutamyltransferase in Lactococcus lactis and Immobilization on Magnetic Nanoparticles. ACS Food Sci. Technol. 2021, 1, 778–787. [Google Scholar] [CrossRef]
- Kuipers, O.P.; de Ruyter, P.G.; Kleerebezem, M.; de Vos, W.M. Controlled overproduction of proteins by lactic acid bacteria. Trends Biotechnol. 1997, 15, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Valdes, J.A.; Huang, C.; Kok, J.; Kuipers, O.P. Another Breaker of the Wall: The Biological Function of the Usp45 Protein of Lactococcus lactis. Appl. Environ. Microbiol. 2020, 86, e00903-20. [Google Scholar] [CrossRef]
- Qin, X.S.; Sun, Q.Q.; Zhao, Y.Y.; Zhong, X.Y.; Mu, D.D.; Jiang, S.T.; Luo, S.Z.; Zheng, Z. Transglutaminase-set colloidal properties of wheat gluten with ultrasound pretreatments. Ultrason. Sonochem. 2017, 39, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Luo, S.; Cai, J.; Sun, Q.; Zhao, Y.; Zhong, X.; Jiang, S.; Zheng, Z. Effects of partial hydrolysis and subsequent cross-linking on wheat gluten physicochemical properties and structure. Food Chem. 2016, 197, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Seyed-Moslemi, S.A.; Hesari, J.; Peighambardoust, S.H.; Peighambardoust, S.J. Effect of microbial lipase and transglutaminase on the textural, physicochemical, and microbial parameters of fresh quark cheese. J. Dairy Sci. 2021, 104, 7489–7499. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N. Innovation in the food industry using microbial transglutaminase: Keys to success and future prospects. Anal. Biochem. 2020, 597, 113638. [Google Scholar] [CrossRef]
- Dinh, T.N.; Hou, S.; Park, S.; Shalek, B.A.; Jeong, K.J. Gelatin Hydrogel Combined with Polydopamine Coating to Enhance Tissue Integration of Medical Implants. ACS Biomater. Sci. Eng. 2018, 4, 3471–3477. [Google Scholar] [CrossRef]
- Türkanoğlu Özçelik, A.; Ersöz, F.; İnan, M. Extracellular production of the recombinant bacterial transglutaminase in Pichia pastoris. Protein Expres. Purif. 2019, 159, 83–90. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kumazawa, Y.; Miwa, K.; Yamanaka, S. ε-(γ-Glutamyl)lysine cross-links of spore coat proteins and transglutaminase activity in Bacillus subtilis. FEMS Microbiol. Lett. 1996, 144, 157–160. [Google Scholar] [CrossRef]
- Souza, C.F.V.D.; Venzke, J.G.E.; Flores, S.H.; Ayub, M.A.Z. Enzymatic properties of transglutaminase produced by a new strain of Bacillus circulans BL32 and its action over food proteins. LWT-Food Sci. Technol. 2011, 44, 443–450. [Google Scholar] [CrossRef]
- Matthias, T.; Jeremias, P.; Neidhöfer, S.; Lerner, A. The industrial food additive, microbial transglutaminase, mimics tissue transglutaminase and is immunogenic in celiac disease patients. Autoimmun. Rev. 2016, 15, 1111–1119. [Google Scholar] [CrossRef]
- Marx, C.K.; Hertel, T.C.; Pietzsch, M. Purification and activation of a recombinant histidine-tagged pro-transglutaminase after soluble expression in Escherichia coli and partial characterization of the active enzyme. Enzym. Microb. Technol. 2008, 42, 568–575. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y. Expression of recombinant transglutaminase gene in Pichia pastoris and its uses in restructured meat products. Food Chem. 2019, 291, 245–252. [Google Scholar] [CrossRef]
- Fu, L.; Wang, Y.; Ju, J.; Cheng, L.; Xu, Y.; Yu, B.; Wang, L. Extracellular production of active-form Streptomyces mobaraensis transglutaminase in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2020, 104, 623–631. [Google Scholar] [CrossRef]
- Yu, Y.J.; Wu, S.C.; Chan, H.H.; Chen, Y.C.; Yang, C. Overproduction of soluble recombinant transglutaminase from Streptomyces netropsis in Escherichia coli. Appl. Microbiol. Biotechnol. 2008, 81, 523. [Google Scholar] [CrossRef]
- Duarte, L.S.; Barsé, L.Q.; Dalberto, P.F.; da Silva, W.T.S.; Rodrigues, R.C.; Machado, P.; Basso, L.A.; Bizarro, C.V.; Ayub, M.A.Z. Cloning and expression of the Bacillus amyloliquefaciens transglutaminase gene in E. coli using a bicistronic vector construction. Enzym. Microb. Technol. 2020, 134, 109468. [Google Scholar] [CrossRef]
- Liu, S.; Wan, D.; Wang, M.; Madzak, C.; Du, G. Overproduction of pro-transglutaminase from Streptomyces hygroscopicus in Yarrowia lipolytica and its biochemical characterization. BMC Biotechnol. 2015, 15, 75. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, P.A.; Ismail, A.A.; Ramaswamy, H.S. High pressure gelation of soy proteins: Effect of concentration, pH and additives. J. Food Eng. 2008, 88, 331–340. [Google Scholar] [CrossRef]
- Qin, X.S.; Chen, S.S.; Li, X.J.; Luo, S.Z.; Zhong, X.Y.; Jiang, S.T.; Zhao, Y.Y.; Zheng, Z. Gelation Properties of Transglutaminase-Induced Soy Protein Isolate and Wheat Gluten Mixture with Ultrahigh Pressure Pretreatment. Food Bioprocess. Technol. 2017, 10, 866–874. [Google Scholar] [CrossRef]
- Li, C.; Wu, X.; Mu, D.; Zhao, Y.; Luo, S.; Zhong, X.; Jiang, S.; Li, X.; Zheng, Z. Effect of Partial Hydrolysis with Papain on the Characteristics of Transglutaminase-Crosslinked Tofu Gel. J. Food Sci. 2018, 83, 3092–3098. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W.; Maccallum, P.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zhao, X.; de Jong, A.; Zhou, Z.; Kuipers, O.P. Complete Genome Sequence of Bacillus amyloliquefaciens Strain BH072, Isolated from Honey. Genome Announc. 2015, 3, e00098-15. [Google Scholar] [CrossRef] [Green Version]
- de Ruyter, P.G.; Kuipers, O.P.; de Vos, W.M. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol. 1996, 62, 3662–3667. [Google Scholar] [CrossRef] [Green Version]
- Brunelle, J.L.; Green, R. One-dimensional SDS-polyacrylamide gel electrophoresis (1D SDS-PAGE). Method Enzym. 2014, 541, 151–159. [Google Scholar]
- Ji, F.; Xu, J.; Ouyang, Y.; Mu, D.; Li, X.; Luo, S.; Shen, Y.; Zheng, Z. Effects of NaCl concentration and temperature on fibrillation, structure, and functional properties of soy protein isolate fibril dispersions. LWT-Food Sci. Technol. 2021, 149, 111862. [Google Scholar] [CrossRef]
- Fu, H.; Li, J.; Yang, X.; Swallah, M.S.; Gong, H.; Ji, L.; Meng, X.; Lyu, B.; Yu, H. The heated-induced gelation of soy protein isolate at subunit level: Exploring the impacts of α and α′ subunits on SPI gelation based on natural hybrid breeding varieties. Food Hydrocolloid. 2023, 134, 108008. [Google Scholar] [CrossRef]
- Gao, H.L.; Xu, J.J.; Tan, M.N.; Mu, D.D.; Li, X.J.; Zhao, Y.Y.; Zheng, Z. Effect of high-intensity ultrasound soymilk pretreatment on the physicochemical properties of microbial transglutaminase-catalyzed tofu gel. J. Food Sci. 2021, 86, 2410–2420. [Google Scholar] [CrossRef]






| Sample | Enzyme Dosage (mL) | Hardness (g) | Springines-s (mm) | Cohesivene-ss (-) | Gumminess (g) |
|---|---|---|---|---|---|
| commercialMTGase | 1 | 14.78 ± 0.7 b | 0.74 ± 0.08 b | 0.39 ± 0.03 b | 14.17 ± 1.03 b |
| BaMTGase | 1 | 108.33 ± 0.6 a | 1.80 ± 1.20 a | 0.81 ± 0.02 ab | 192.71 ± 3.88 a |
| Content of Secondary Structure (%) | ||||
|---|---|---|---|---|
| Sample | α-Helix | β-Sheets | β-Turns | Random Coils |
| commercialMTGase | 16.91 ± 0.69 a | 51.79 ± 0.75 b | 13.65 ± 0.49 b | 17.65 ± 0.58 a |
| BaMTGase | 15.02 ± 1.01 b | 53.1 ± 0.24 a | 16.40 ± 0.52 a | 15.48 ± 0.21 b |
| Strain | Characteristic | Information |
|---|---|---|
| Bacillus amyloliquefaciens BH072 | Amplification of mtg | Lab collection [40], BaMTGase wild-type producer |
| Lactococcus lactis NZ9000 | heterologous host | pepN::nisRK [41] |
| Plasmid | Characteristic | Information |
| pNZ8048 | PnisA promoter, CmR | Inducible plasmid for NICE expression |
| pNZ8048-SPusp45-promtg | Amplification of spusp45-proregion gene, CmR | Lab collection [15] |
| pNZ8048-SPusp45-proBamtg | Recombinant expression plasmid, CmR | This work |
| Primer | Sequence (5′–3′) | Property/Function |
| P1 | CATGCCATGGCAAAAAAGATTATCTCAGCTATTTTAAT | Amplification of the SPusp45-proregionwhich preceded by NcoI restriction site |
| P2 | TGGCCGGATATGATAATCATGGGGGCCCGGA | Amplification of the SPusp45-proregion, P2 and P3 complement in reverse |
| P3 | TCCGGGCCCCCATGATTATCATATCCGGCCA | Amplification of mtg gene, P3 and P2 are reverse complementary sequences |
| P4 | GGGGTACCTTAGTGATGGTGATGGTGATGATGCATGATCTGATAAAGCG | Amplification of mtg gene. Hexahistidine codons were followed by a KpnI restriction site |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Li, X.; Montalbán-López, M.; Wu, X.; Zheng, Z.; Mu, D. Secretion of Bacillus amyloliquefaciens Transglutaminase from Lactococcus lactis and Its Enhancement of Food Gel Properties. Gels 2022, 8, 674. https://doi.org/10.3390/gels8100674
Ma T, Li X, Montalbán-López M, Wu X, Zheng Z, Mu D. Secretion of Bacillus amyloliquefaciens Transglutaminase from Lactococcus lactis and Its Enhancement of Food Gel Properties. Gels. 2022; 8(10):674. https://doi.org/10.3390/gels8100674
Chicago/Turabian StyleMa, Tiange, Xingjiang Li, Manuel Montalbán-López, Xuefeng Wu, Zhi Zheng, and Dongdong Mu. 2022. "Secretion of Bacillus amyloliquefaciens Transglutaminase from Lactococcus lactis and Its Enhancement of Food Gel Properties" Gels 8, no. 10: 674. https://doi.org/10.3390/gels8100674