
Antimicrobial and Antioxidant Activities of Endophytic Fungi Associated with Arrabidaea chica (Bignoniaceae)


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Endophytic Fungi
2.3. Production of Fungal Metabolites
2.4. Antimicrobial Testing
2.5. Antioxidant Assays
2.6. Chemical Profile of Fungal Extracts
2.7. Dosage of Total Phenolic Content
2.8. Identification of the Most Promising Fungi
2.9. Statistical Analysis
3. Results
3.1. Antimicrobial Activity of Fungal Extracts
3.2. Antioxidant Activity of Fungal Extracts
3.3. Chemical Profile of Promising Fungal Extracts
3.4. Total Phenolic Content of the Most-Active Fungal Extracts
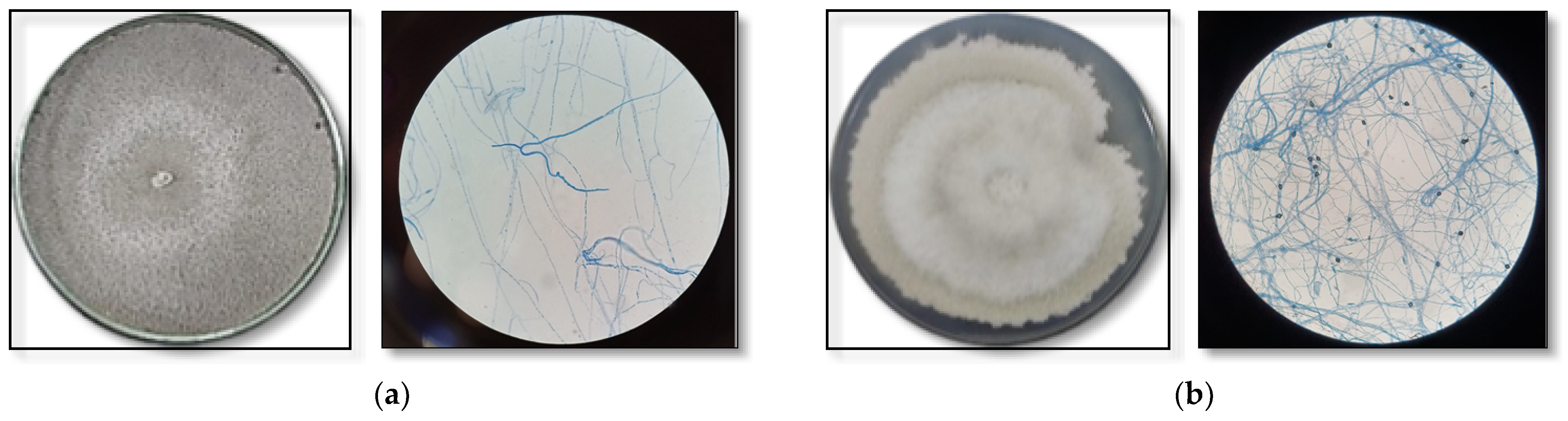
3.5. Identification of Endophytic Fungi That Produce Bioactive Substances
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferreira, M.G.R. Crajiru (Arrabidaea chica Verlot); Embrapa: Porto Velho, Brazil, 2005; Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/24786/1/folder-crajiru.pdf (accessed on 11 October 2019).
- Chapman, E.; Perkin, A.G.; Robinson, R. CCCCII—The colouring matters of Carajura. J. Chem. Soc. 1927, 1927, 3015–3041. [Google Scholar] [CrossRef]
- Zorn, B.; Garcia-Pineres, A.; Castro, V.; Murillo, R.; Mora, G.; Merfort, I. 3-Desoxyanthocyanidins from Arrabidaea chica. Phytochemistry 2001, 56, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.C.F.; Pohlit, A.M.; Chaves, F.C.M. Effect of substrate and cutting diameter on the propagation of Arrabidaea chica (Humb. & Bonpl.) B. Verl. (Bignoniaceae). Rev. Bras. Plantas Med. 2008, 10, 38–42. [Google Scholar]
- Mafioleti, L.; da Silva Junior, I.F.; Colodel, E.M.; Flach, A.; Martins, D.T. Evaluation of the toxicity and antimicrobial activity of hydroethanolic extract of Arrabidaea chica (Humb. & Bonpl.) B. Verl. J. Ethnopharmacol. 2013, 150, 576–582. [Google Scholar] [CrossRef]
- Michel, A.F.; Melo, M.M.; Campos, P.P.; Oliveira, M.S.; Oliveira, F.A.S.; Cassali, G.D.; Ferraz, V.P.; Cota, B.B.; Andrade, S.P.; Souza-Fagundes, E.M. Evaluation of anti-inflammatory, antiangiogenic and antiproliferative activities of Arrabidaea chica crude extracts. J. Ethnopharmacol. 2015, 165, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Aro, A.A.; Simões, G.F.; Esquisatto, M.A.M.; Foglio, M.A.; Carvalho, J.E.; Oliveira, A.L.R.; Gomes, L.; Pimentel, E.R. Arrabidaea chica extract improves gait recovery and changes collagen content during healing of the Achilles tendon. Injury 2013, 44, 884–892. [Google Scholar] [CrossRef]
- Miranda, N.; Gerola, A.P.; Novello, C.R.; Ueda-Nakamura, T.; de Oliveira Silva, S.; Dias-Filho, B.P.; Hioka, N.; de Mello, J.C.P.; Nakamura, C.V. Pheophorbide a, a compound isolated from the leaves of Arrabidaea chica, induces photodynamic inactivation of Trypanosoma cruzi. Photodiagn. Photodyn. Ther. 2017, 19, 256–265. [Google Scholar] [CrossRef]
- Siraichi, J.T.G.; Felipe, D.F.; Brambilla, L.Z.R.; Gatto, M.J.; Terra, V.A.; Cecchini, A.L.; Cortez, L.E.R.; Rodrigues-Filho, E.; Cortez, D.A.G. Antioxidant capacity of the leaf extract obtained from Arrabidaea chica cultivated in Southern Brazil. PLoS ONE 2013, 8, 72733. [Google Scholar] [CrossRef]
- Martins, F.J.; Caneschi, C.A.; Vieira, J.L.F.; Barbosa, W.; Raposo, N.R.B. Antioxidant activity and potential photoprotective from amazon native flora extracts. J. Photochem. Photobiol. B 2016, 161, 34–39. [Google Scholar] [CrossRef]
- Ribeiro, F.M.; Volpato, H.; Lazarin-Bidóia, D.; Desoti, V.C.; de Souza, R.O.; Fonseca, M.J.V.; Ueda-Nakamura, T.; Nakamura, C.V.; Silva, S.O. The extended production of UV-induced reactive oxygen species in L929 fibroblasts is attenuated by posttreatment with Arrabidaea chica through scavenging mechanisms. J. Photochem. Photobiol. B 2018, 178, 175–181. [Google Scholar] [CrossRef]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G. The emergence of endophytic microbes and their biological promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Gurgel, R.S.; Rodrigues, J.G.C.; Matias, R.R.; Barbosa, B.N.; Oliveira, R.L.; Albuquerque, P.M. Biological activity and production of metabolites from Amazon endophytic fungi. Afr. J. Microbiol. Res. 2020, 14, 85–93. [Google Scholar] [CrossRef]
- Omomowo, I.O.; Amao, J.A.; Abubakar, A.; Ogundola, A.F.; Ezediuno, L.O.; Bamigboye, C.O. A review on the trends of endophytic fungi bioactivities. Sci. Afr. 2023, 20, e01594. [Google Scholar] [CrossRef]
- Strobel, G.A. Endophytes as sources of bioactive products. Microbes Infect. 2003, 5, 535–544. [Google Scholar] [CrossRef]
- Banhos, E.F.; Souza, A.Q.; Andrade, J.C.; Souza, A.D.L.; Koolen, H.H.F.; Albuquerque, P.M. Endophytic fungi from Myrcia guianensis at the Brazilian Amazon: Distribution and bioactivity. Braz. J. Microbiol. 2014, 45, 153–161. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- Nagarajan, D. In vitro antioxidant potential of endophytic fungi isolated from Enicostemma axillare (Lam.) Raynal. and Ormocarpum cochinchinense (Lour.) Merr. J. Pharmacogn. Phytochem. 2019, 8, 1356–1363. [Google Scholar]
- Dhayanithy, G.; Subban, K.; Chelliah, J. Diversity and biological activities of endophytic fungi associated with Catharanthus roseus. BMC Microbiol. 2019, 19, 22. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Elucidating mechanisms of endophytes used in plant protection and other bioactives with multifunctional prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [Google Scholar] [CrossRef]
- Silva, A.A.; Polonio, J.C.; Bulla, A.M.; Polli, A.D.; Castro, J.C.; Soares, L.C.; Oliveira-Junior, V.A.; Vicentini, V.E.P.; Oliveira, A.J.B.; Gonçalves, J.E.; et al. Antimicrobial and antioxidant activities of secondary metabolites from endophytic fungus Botryosphaeria fabicerciana (MGN23-3) associated to Morus nigra L. Nat. Prod. Res. 2021, 36, 3158–3162. [Google Scholar] [CrossRef]
- Batista, B.N.; Matias, R.R.; Oliveira, R.L.E.; Albuquerque, P.M. Hydrolytic enzyme production from açai palm (Euterpe precatoria) endophytic fungi and characterization of the amylolytic and cellulolytic extracts. World J. Microbiol. Biotechnol. 2022, 38, 30. [Google Scholar] [CrossRef] [PubMed]
- Elawady, M.E.; Hamed, A.A.; Alsallami, W.M.; Gabr, E.Z.; Abdel-Monem, M.O.; Hassan, M.G. Bioactive metabolite from endophytic Aspergillus versicolor SB5 with anti-acetylcholinesterase, anti-inflammatory and antioxidant activities: In vitro and in silico studies. Microorganisms 2023, 11, 1062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Li, X.; Han, M.M.; Cai, Z.Y.; Gao, X.; Pang, M.X.; Qi, J.H.; Wang, F. Separating and purifying of endophytic fungi from Ginkgo biloba and screening of flavonoid-producing strains. IOP Conf. Ser. Earth Environ. Sci. 2019, 371, 042052. [Google Scholar] [CrossRef]
- Araújo, W.L.; Lima, A.O.S.; Azevedo, J.L.; Marcon, J.; Kuklinsky, S.J.; Lacava, P.T. Manual: Isolamento de Microrganismos Endofíticos; Universidade de São Paulo: Piracicaba, Brazil, 2002. [Google Scholar]
- Castellani, A. Viability of some pathogenic fungi in distilled water. J. Trop. Med. Hyg. 1939, 42, 225–226. [Google Scholar]
- Bose, P.; Gowrie, S.U.; Chathurdevi, G. Optimization of culture conditions of growth and production of bioactive metabolites by endophytic fungus—Aspergillus tamarii. Int. J. Pharm. Biol. Sci. 2019, 9, 469–478. [Google Scholar] [CrossRef]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D. Don. 3 Biotech 2016, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. CLSI Standard M27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Duarte-Almeida, J.M.; Dos Santos, R.J.; Genovese, M.I.; Lajolo, F.M. Evaluation of the antioxidant activity using the b-carotene/linoleic acid system and the DPPH scavenging method. Food Sci. Technol. 2006, 26, 446–452. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Lopes, J.L.C. Cromatografia em Camada Delgada. In Fundamentos de Cromatografia; Collins, C.H., Braga, G.L., Bonato, P.S., Eds.; Editora da Unicamp: Campinas, Brazil, 2006; pp. 67–86. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Watanabe, T. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key to Species; CRC Press: Boca Raton, FL, USA, 2002; 504p. [Google Scholar]
- Procop, G.W.; Church, D.L.; Hall, G.S.; Janda, W.S.; Koneman, E.W.; Schreckenberger, P.C.; Woods, G.L. Koneman’s Color Atlas and Textbook of Diagnostic Microbiology; Jones & Bartlett Learning: Burlington, NJ, USA, 2016; 1830p. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi; Burgess Publishing Company: Minneapolis, MN, USA, 1972. [Google Scholar]
- Hanlin, R.T.; Menezes, M. Gêneros Ilustrados de Ascomicetos; Imprensa da Universidade Federal Rural de Pernambuco: Recife, Brazil, 1996; 244p. [Google Scholar]
- Doyle, J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Oetari, A.; Rahmadewi, M.; Rachmania, M.K.; Sjamsuridzal, W. Molecular identification of fungal species from deteriorated old chinese manuscripts in Central Library Universitas Indonesia. AIP Conf. Proc. 2018, 2023, 020122. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis, program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S. Endophyte research: Going beyond isolation and metabolite documentation. Fungal Ecol. 2013, 6, 561–568. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, J.; Chen, C.; Mo, X.; Tan, Q.; He, Y.; Wang, Z.; Yin, J.; Zhou, G. The Multifunctions and future prospects of endophytes and their metabolites in plant disease management. Microorganisms 2022, 10, 1072. [Google Scholar] [CrossRef]
- Verpoorte, R. Exploration of nature’s chemodiversity: The role of secondary metabolites as leads in drug development. Drug Disc. Today 1998, 3, 232–238. [Google Scholar] [CrossRef]
- Santos, I.P.; Silva, L.C.N.; Silva, M.V.; Aragão, J.M.; Cavalcanti, M.S.; Lima, V.L. Antibacterial activity of endophytic fungi from leaves of Indigofera suffruticosa Miller (Fabaceae). Front. Microbiol. 2015, 6, 350. [Google Scholar] [CrossRef]
- Matias, J.N.; Souza, G.A.; Joshi, R.K.; Marqui, S.V.; Guiguer, E.L.; Araújo, A.C.; Otoboni, A.M.M.B.; Marineli, P.; Barbalho, M. Arrabidaea chica (Humb. And Bonpl.): A plant multipurpose medicinal applications. Int. J. Herb. Med. 2021, 9, 77–87. [Google Scholar]
- Gardner, D.E. Botryosphaeria mamane sp. nov. associated with witches’-brooms on the endemic forest tree Sophora chrysophylla in Hawaii. Mycologia 1997, 89, 298–303. [Google Scholar] [CrossRef]
- Pongcharoen, W.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J. A new dihydrobenzofuran derivative from the endophytic fungus Botryosphaeria mamane PSU-M76. Chem. Pharm. Bull. 2007, 55, 1404–1405. [Google Scholar] [CrossRef]
- Oliveira, F.C.; Barbosa, F.G.; Mafezoli, J.; Oliveira, M.C.F.; Camelo, A.L.M.; Longhinotti, E.; Lima, A.C.A.; Câmara, M.P.S.; Gonçalves, F.J.T.; Freired, F.C.O. Volatile organic compounds from filamentous fungi: A chemotaxonomic tool of the Botryosphaeriaceae family. J. Braz. Chem. Soc. 2015, 26, 2189–2194. [Google Scholar] [CrossRef]
- Triastuti, A.; Vansteelandt, M.; Barakat, F.; Trinel, M.; Jargeat, P.; Fabre, N.; Amasifuen, C.; Mejia, K.; Valentin, A.; Haddad, M. How histone deacetylase inhibitors alter the secondary metabolites of Botryosphaeria mamane, an endophytic fungus isolated from Bixa orellana, L. Chem. Biodivers. 2019, 16, e1800485. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Q.; Gao, Y.-Q.; Tang, J.-J.; Zhang, A.-L.; Gao, J.-M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef]
- Rukachaisirikul, V.; Arunpanichlert, J.; Sukpondma, Y.; Phongpaichit, S.; Sakayaroj, J. Metabolites from the endophytic fungi Botryosphaeria rhodina PSU-M35 and PSU-M114. Tetrahedron 2009, 65, 10590–10595. [Google Scholar] [CrossRef]
- Linares, A.M.P.; Hernandes, C.; França, S.C.; Lourenço, M.V. Phytoregulatory activity of jasmonates produced by Botryosphaeria rhodina. Hortic. Bras. 2010, 28, 430–434. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, C.-H.; Zheng, Z.-H.; Shen, Y.-M. New polyketides isolated from Botryosphaeria australis strain ZJ12-1A. Helv. Chim. Acta 2011, 94, 897–902. [Google Scholar] [CrossRef]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.A.; Zamora, C.M.P.; Nuñez, M.B.; Gonzalez, A.M. In vitro antioxidant, antilipoxygenase and antimicrobial activities of extracts from seven climbing plants belonging to the Bignoniaceae. J. Integr. Med. 2018, 16, 255–262. [Google Scholar] [CrossRef]
- Matos, F.J.A. Plantas Medicinais no Brasil: Nativas e Exóticas; Jardim Botânico Plantarum: Nova Odessa, Brazil, 2021. [Google Scholar]
- Prior, R.L.; Wu, X. Diet antioxidant capacity: Relationship to oxidative stress and health. Am. J. Biomed. Sci. 2013, 5, 126–139. [Google Scholar] [CrossRef]
- Putri, A.R.; Salni, S.; Widjajanti, H. Antioxidant activity of the secondary metabolites produced by endophytic fungi isolated from Jeruju (Acanthus ilicifolius L.) plant. Biovalentia Biol. Res. J. 2019, 5, 14–19. [Google Scholar] [CrossRef]
- Kim, J.W.; Shim, S.H. The fungus Colletotrichum as a source for bioactive secondary metabolites. Arch. Pharm. Res. 2019, 42, 735–753. [Google Scholar] [CrossRef]
- Colletotrichum gloeosporioides isolated from Piper nigrum. Phytomedicine 2014, 21, 534–540. [CrossRef]
- Tianpanich, K.; Prachya, S.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Radical scavenging and antioxidant activities of isocoumarins and a phthalide from the endophytic fungus Colletotrichum sp. J. Nat. Prod. 2011, 74, 79–81. [Google Scholar] [CrossRef]
- Mahmud, S.M.N.; Sohrab, M.H.; Begum, M.N.; Rony, S.R.; Sharmin, S.; Moni, F.; Akhter, S.; Mohiuddin, A.K.M.; Afroz, F. Cytotoxicity, antioxidant, antimicrobial studies and phytochemical screening of endophytic fungi isolated from Justicia gendarussa. Ann. Agric. Sci. 2020, 65, 225–232. [Google Scholar] [CrossRef]
- Budiono, B.; Elfita, E.; Muharni, M.; Yohandini, H.; Widjajanti, H. Antioxidant activity of Syzygium samarangense L. and their endophytic fungi. Molekul 2019, 14, 48–55. [Google Scholar] [CrossRef]
- Alves, D.R.; Silva, W.M.B.; Santos, D.L.; Freire, F.C.O.; Vasconcelos, F.R.; Morais, S.M. Antioxidant, anticolinesterasic and cytoxic activities of endophytic fungus metabolites. Braz. J. Dev. 2020, 6, 73684–73691. [Google Scholar] [CrossRef]
- Druzian, S.P.; Pinheiro, L.N.; Susin, N.M.B.; Dal Prá, V.; Mazutti, M.A.; Kuhn, R.C.; Terra, L.M. Production of metabolites with antioxidant activity by Botryosphaeria dothidea in submerged fermentation. Bioprocess Biosyst. Eng. 2019, 43, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Pulido, R.; Bravo, L.; Saura-Calixto, F. Antioxidant activity of dietary as determined by a modified ferric reducing/antioxidant power assay. J. Agr. Food Chem. 2000, 48, 3396–3402. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C.-M. Determination of antioxidants by DPPH radical scavenging activity and quantitative phytochemical analysis of Ficus religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef]
- Aguirre, J.J.; De La Garza, T.H.; Zugasti, C.A.; Belmares, C.R.; Aguilar, C.N. The optimization of phenolic compounds extraction from cactus pear (Opuntia ficus-indica) skin in a reflux system using response surface methodology. Asian Pac. J. Trop. Biomed. 2013, 3, 436–442. [Google Scholar] [CrossRef]
- Kaur, N.; Arora, D.S.; Kalia, N.; Kaur, M. Antibiofilm, antiproliferative, antioxidant and antimutagenic activities of on endophytic fungus Aspergillus fumigatus from Moringa oleifera. Mol. Biol. Rep. 2020, 47, 2901–2911. [Google Scholar] [CrossRef]
- Santos, R.I. Metabolismo Básico e Origem dos Metabólitos Secundários. In Farmacognosia—da Planta ao Medicamento, 5th ed.; Simões, C.M.O., Schenkel, E.P., Gosmann, G., Mello, J.C.P., Mentz, L.A., Petrovick, P.R., Eds.; UFRGS/UFSC: Porto Alegre/Florianópolis, Brazil, 2003; pp. 403–434. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosinthetic Approach; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Venkatasubbaiah, P.; Sutton, T.B.; Chilton, W.S. Effect of phytotoxins produced by Botryosphaeria obtusa, the cause of black rot of apple fruit and frogeye leaf spot. Phytopathology 1991, 81, 243–247. [Google Scholar] [CrossRef]
- Djoukeng, J.D.; Polli, S.; Larignon, P.; Abou-Mansour, E. Identification of phytotoxins from Botryosphaeria obtusa, a pathogen of black dead arm disease of grapevine. Eur. J. Plant Pathol. 2009, 124, 303–308. [Google Scholar] [CrossRef]
- Chen, X.W.; Yang, Z.D.; Sun, J.H.; Song, T.T.; Zhu, B.Y.; Zhao, J.W. Colletotrichine A, a new sesquiterpenoid from Colletotrichum gloeosporioides GT-7, a fungal endophyte of Uncaria rhynchophylla. Nat. Prod. Res. 2018, 32, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Sharma, R.K.; Arora, D.S. Antioxidant compounds from microbial sources: A review. Food Res. Int. 2020, 129, 108849. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endophytic Fungus Code | Specimen | Plant Part | Number of Isolates | |||
|---|---|---|---|---|---|---|
| CF1-1 | CF1-12 | CF1-22 | CF1-31 | 1 | Leaves | 33 |
| CF1-2 | CF1-13 | CF1-23 | CF1-32 | |||
| CF1-3 | CF1-15 | CF1-24 | CF1-34 | |||
| CF1-4 | CF1-16 | CF1-25 | CF1-35 | |||
| CF1-5 | CF1-17 | CF1-26 | CF1-36 | |||
| CF1-6 | CF1-18 | CF1-27 | CF1-37 | |||
| CF1-7 | CF1-19 | CF1-28 | ||||
| CF1-9 | CF1-20 | CF1-29 | ||||
| CF1-11 | CF1-21 | CF1-30 | ||||
| CG1-1 | CG1-5 | CG1-9 | CG1-12 | 1 | Branches | 11 |
| CG1-2 | CG1-7 | CG1-10 | CG1-14 | |||
| CG1-4 | CG1-8 | CG1-11 | ||||
| CF2-1 | CF2-7 | CF2-13 | CF2-17 | 2 | Leaves | 16 |
| CF2-2 | CF2-9 | CF2-14 | CF2-18 | |||
| CF2-3 | CF2-11 | CF2-15 | CF2-19 | |||
| CF2-6 | CF2-12 | CF2-16 | CF2-20 | |||
| CG2-2 | CG2-5 | CG2-10 | CG2-15 | 2 | Branches | 11 |
| CG2-3 | CG2-7 | CG2-11 | CG2-16 | |||
| CG2-4 | CG2-8 | CG2-12 | ||||
| CF3-1 | CF3-9 | CF3-15 | CF3-21 | 3 | Leaves | 22 |
| CF3-2 | CF3-10 | CF3-16 | CF3-23 | |||
| CF3-4 | CF3-11 | CF3-17 | CF3-24 | |||
| CF3-5 | CF3-13 | CF3-18 | CF3-26 | |||
| CF3-6 | CF3-12 | CF3-19 | ||||
| CF3-7 | CF3-14 | CF3-20 | ||||
| CG3-1 | CG3-8 | CG3-13 | CG3-18 | 3 | Branches | 14 |
| CG3-3 | CG3-10 | CG3-15 | CG3-19 | |||
| CG3-4 | CG3-11 | CG3-16 | ||||
| CG3-7 | CG3-12 | CG3-17 | ||||
| Total | 107 | |||||
| Endophytic Fungi Code | MIC (mg/mL) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC | SA | CA | PA | PM | BS | SEp | EF | SM | KP | Sen | CT | CP | AB | |
| CF1-2 | 5.00 | - | - | - | - | 5.00 | - | - | - | - | NT | NT | NT | NT |
| CF1-26 | 5.00 | - | - | - | - | - | - | - | - | - | 5.00 | NT | NT | NT |
| CF1-29 | 5.00 | - | - | - | - | - | - | - | - | - | 2.50 | NT | NT | NT |
| CF1-37 | - | 5.00 | 1.25 | - | - | - | - | - | - | - | - | 1.25 | 2.50 | - |
| CG1-10 | - | 5.00 | - | - | - | 5.00 | - | 5.00 | - | - | 2.50 | NT | NT | NT |
| CG1-1 | 5.00 | 1.25 | 1.25 | 5.00 | 5.00 | - | - | - | - | - | - | 5.00 | 5.00 | - |
| CG1-2 | - | 5.00 | - | 5.00 | 5.00 | - | - | - | 2.50 | 5.00 | 5.00 | NT | NT | NT |
| CG1-5 | 5.00 | 5.00 | - | - | - | - | - | 5.00 | - | - | 5.00 | NT | NT | NT |
| CG1-8 | 5.00 | - | - | - | - | 5.00 | 5.00 | 5.00 | - | 5.00 | 5.00 | NT | NT | NT |
| CG1-9 | 5.00 | - | - | - | - | - | 5.00 | 5.00 | - | - | - | NT | NT | NT |
| CF2-11 | - | 5.00 | 2.50 | - | - | 2.50 | 1.25 | - | - | - | - | 5.00 | 5.00 | - |
| CF2-13 | 2.50 | 0.312 | 1.25 | - | 5.00 | 2.50 | 1.25 | - | 5.00 | 5.00 | 2.50 | 1.25 | 0.312 | 2.50 |
| CF2-16 | - | - | 1.25 | NT | NT | NT | NT | NT | NT | NT | NT | 2.50 | 1.25 | - |
| CG2-5 | 5.00 | 5.00 | - | - | - | - | 5.00 | 2.50 | - | - | 5.00 | NT | NT | NT |
| CF3-5 | - | - | 1.25 | NT | NT | NT | NT | NT | NT | NT | NT | 2.50 | 1.25 | - |
| CF3-9 | - | 5.00 | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT |
| CF3-14 | 5.00 | 5.00 | - | - | - | - | 2.50 | - | - | - | - | NT | NT | NT |
| CF3-26 | - | - | 5.00 | NT | NT | NT | NT | NT | NT | NT | NT | 5.00 | 5.00 | - |
| Endophytic Fungi Code | AA *(%) | EC50 (μg/mL) | FRAP * (µmol TE/g) | Endophytic Fungi Code | AA * (%) | EC50 (μg/mL) | FRAP * (µmol TE/g) |
|---|---|---|---|---|---|---|---|
| CF1-3 | 98.61 a | 5490 | 110.6 B | CF2-11 | 100.0 a | 1080 | 128.5 B |
| CF1-4 | 100.0 a | 6870 | 77.4 C | CF2-12 | 86.32 b | 6450 | 93.6 C |
| CF1-7 | 92.38 b | 1250 | 174.4 A | CF2-13 | 92.47 b | 360 | 71.9 C |
| CF1-9 | 90.74 b | 5530 | 143.2 B | CF2-14 | 90.22 b | 1620 | 167.9 A |
| CF1-12 | 90.74 b | 5270 | 95.2 C | CF2-16 | 94.81 b | 1170 | 218.6 A |
| CF1-13 | 91.43 b | 6480 | 214.0 A | CF2-17 | 83.03 c | 6820 | 48.0 C |
| CF1-15 | 94.20 b | 2710 | 171.0 A | CF2-18 | 70.74 d | 7540 | 46.5 C |
| CF1-16 | 92.81 b | 6190 | 72.4 C | CF2-20 | 82.68 c | 1660 | 192.5 A |
| CF1-18 | 88.14 b | 2840 | 178.6 A | CG2-2 | 95.06 b | 3200 | 103.3 B |
| CF1-19 | 82.77 c | 6590 | 47.5 C | CG2-4 | 91.95 b | 1520 | 151.3 A |
| CF1-20 | 100.0 a | 990 | 121.9 B | CG2-5 | 89.87 b | 2720 | 51.0 C |
| CF1-23 | 94.20 b | 5730 | 154.0 B | CG2-7 | 95.76 b | 4710 | 59.7 C |
| CF1-24 | 89.70 b | 3420 | 39.1 C | CG2-10 | 98.01 a | 740 | 191.2 A |
| CF1-25 | 93.59 b | 1060 | 188.4 A | CG2-12 | 95.75 b | 4750 | 169.6 A |
| CF1-26 | 81.13 c | 6720 | 177.3 A | CG2-16 | 89.78 b | 3250 | 86.1 C |
| CF1-27 | 95.93 b | 1460 | 125.2 B | CF3-1 | 95.06 b | 2390 | 133.4 B |
| CF1-28 | 93.77 b | 5020 | 181.7 A | CF3-4 | 90.04 b | 6280 | 197.3 A |
| CF1-29 | 93.85 b | 2260 | 109.1 B | CF3-5 | 86.23 c | 940 | 161.9 B |
| CF1-30 | 88.83 b | 3060 | 197.5 A | CF3-9 | 100.0 a | 2480 | 142.3 B |
| CF1-31 | 100.0 a | 1410 | 55.9 C | CF3-11 | 80.26 b | 8310 | 53.2 C |
| CF1-36 | 88.23 b | 2550 | 82.6 C | CF3-13 | 98.35 a | 65,050 | 53.5 C |
| CF1-37 | 93.68 b | 680 | 171.1 A | CF3-14 | 95.76 b | 5400 | 152.8 B |
| CG1-1 | 90.56 b | 2920 | 131.7 B | CF3-16 | 91.52 b | 6100 | 158.2 B |
| CG1-2 | 90.39 b | 3560 | 148.6 B | CF3-17 | 76.97 c | 7610 | 56.9 C |
| CG1-4 | 89.78 b | 1060 | 199.0 A | CF3-18 | 100.0 a | 5650 | 175.8 A |
| CG1-5 | 92.38 b | 5770 | 166.0 A | CF3-20 | 90.30 b | 6250 | 148.5 B |
| CG1-7 | 100.0 a | 11 | 109.5 B | CF3-21 | 85.89 c | 6820 | 139.7 B |
| CG1-10 | 89.44 b | 6420 | 44.7 C | CF3-26 | 88.48 b | 3080 | 172.7 A |
| CG1-11 | 87.97 b | 6660 | 128.2 B | CG3-3 | 80.00 c | 5200 | 83.1 C |
| CG1-12 | 88.92 b | 3700 | 89.5 C | CG3-4 | 90.04 b | 4830 | 114.4 B |
| CG1-14 | 81.99 c | 2880 | 154.8 B | CG3-7 | 79.91 c | 500 | 167.0 A |
| CF2-2 | 75.06 c | 6170 | 106.7 B | CG3-8 | 89.27 b | 7300 | 79.9 C |
| CF2-6 | 80.43 c | 5850 | 82.0 C | CG3-13 | 77.40 c | 7610 | 169.3 A |
| CF2-7 | 78.61 c | 4100 | 59.2 C | CG3-18 | 97.75 a | 5670 | 158.5 B |
| CF2-9 | 94.98 b | 5630 | 78.9 C | CG3-19 | 91.17 b | 6210 | 96.2 C |
| Quercetin | 98.00 a | 8 | NT | Ascorbic acid | NT | NT | 163.1 A |
| GenBank Accession Number | |||||
|---|---|---|---|---|---|
| Isolate | Species | Source | ITS | Βtub | CaM |
| CG1-7 | Colletotrichum sp. | A. chica | OQ390099 | OQ412637 | OQ412636 |
| CF2-13 | Botryosphaeria mamane | A. chica | OQ696843 | OQ703591 | *- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gurgel, R.S.; de Melo Pereira, D.Í.; Garcia, A.V.F.; Fernandes de Souza, A.T.; Mendes da Silva, T.; de Andrade, C.P.; Lima da Silva, W.; Nunez, C.V.; Fantin, C.; de Lima Procópio, R.E.; et al. Antimicrobial and Antioxidant Activities of Endophytic Fungi Associated with Arrabidaea chica (Bignoniaceae). J. Fungi 2023, 9, 864. https://doi.org/10.3390/jof9080864
Gurgel RS, de Melo Pereira DÍ, Garcia AVF, Fernandes de Souza AT, Mendes da Silva T, de Andrade CP, Lima da Silva W, Nunez CV, Fantin C, de Lima Procópio RE, et al. Antimicrobial and Antioxidant Activities of Endophytic Fungi Associated with Arrabidaea chica (Bignoniaceae). Journal of Fungi. 2023; 9(8):864. https://doi.org/10.3390/jof9080864
Chicago/Turabian StyleGurgel, Raiana Silveira, Dorothy Ívila de Melo Pereira, Ana Vyktória França Garcia, Anne Terezinha Fernandes de Souza, Thaysa Mendes da Silva, Cleudiane Pereira de Andrade, Weison Lima da Silva, Cecilia Veronica Nunez, Cleiton Fantin, Rudi Emerson de Lima Procópio, and et al. 2023. "Antimicrobial and Antioxidant Activities of Endophytic Fungi Associated with Arrabidaea chica (Bignoniaceae)" Journal of Fungi 9, no. 8: 864. https://doi.org/10.3390/jof9080864