
Effects of Arbuscular Mycorrhizal Fungi on Robinia pseudoacacia L. Growing on Soils Contaminated with Heavy Metals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Substrate Preparation
2.2. Plant and AMF Preparation
2.3. Pot Experiment
2.4. Sampling and Analysis
2.5. Statistical Analyzes
3. Results
3.1. Soil Characteristics
3.2. Mycorrhizal Colonization and Plant Dry Weight
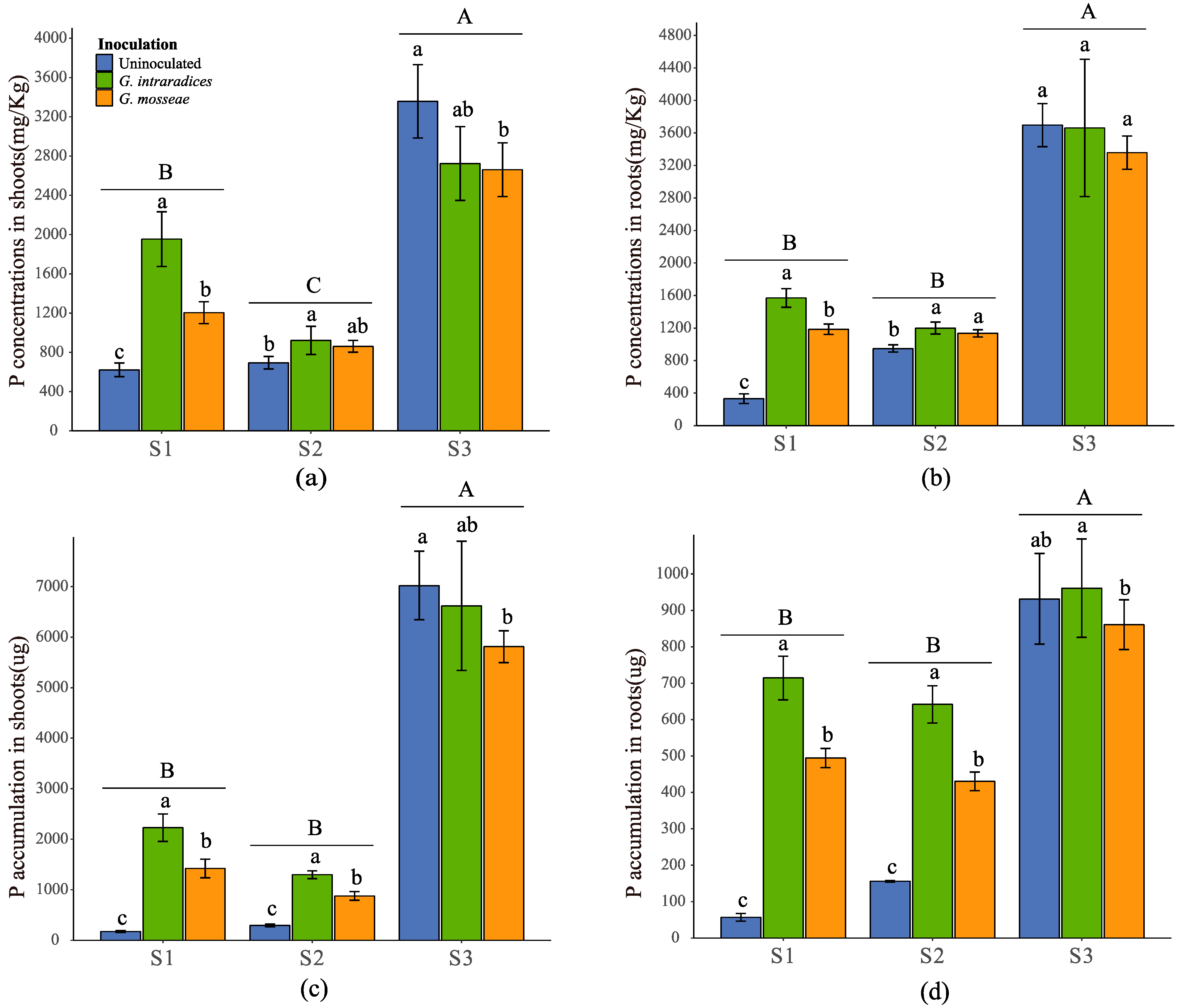
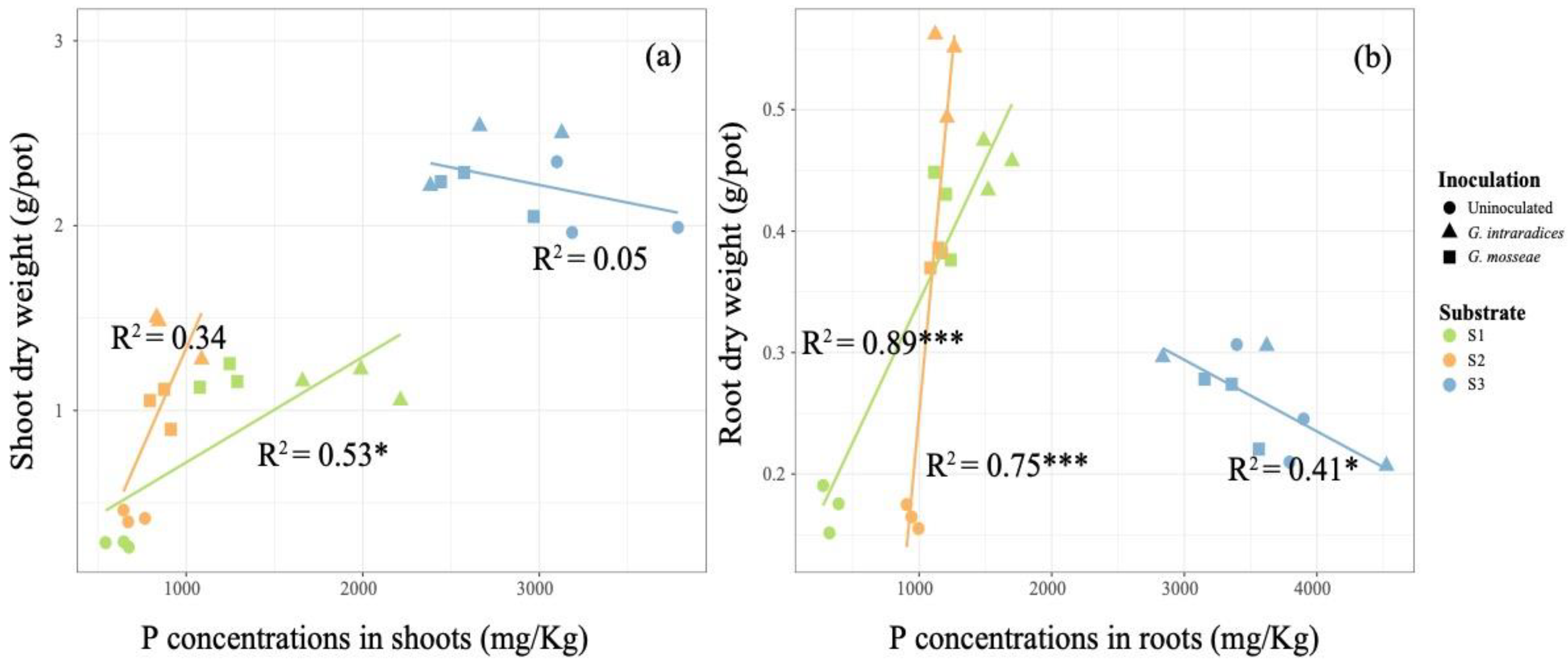
3.3. Plant Uptake of Available P
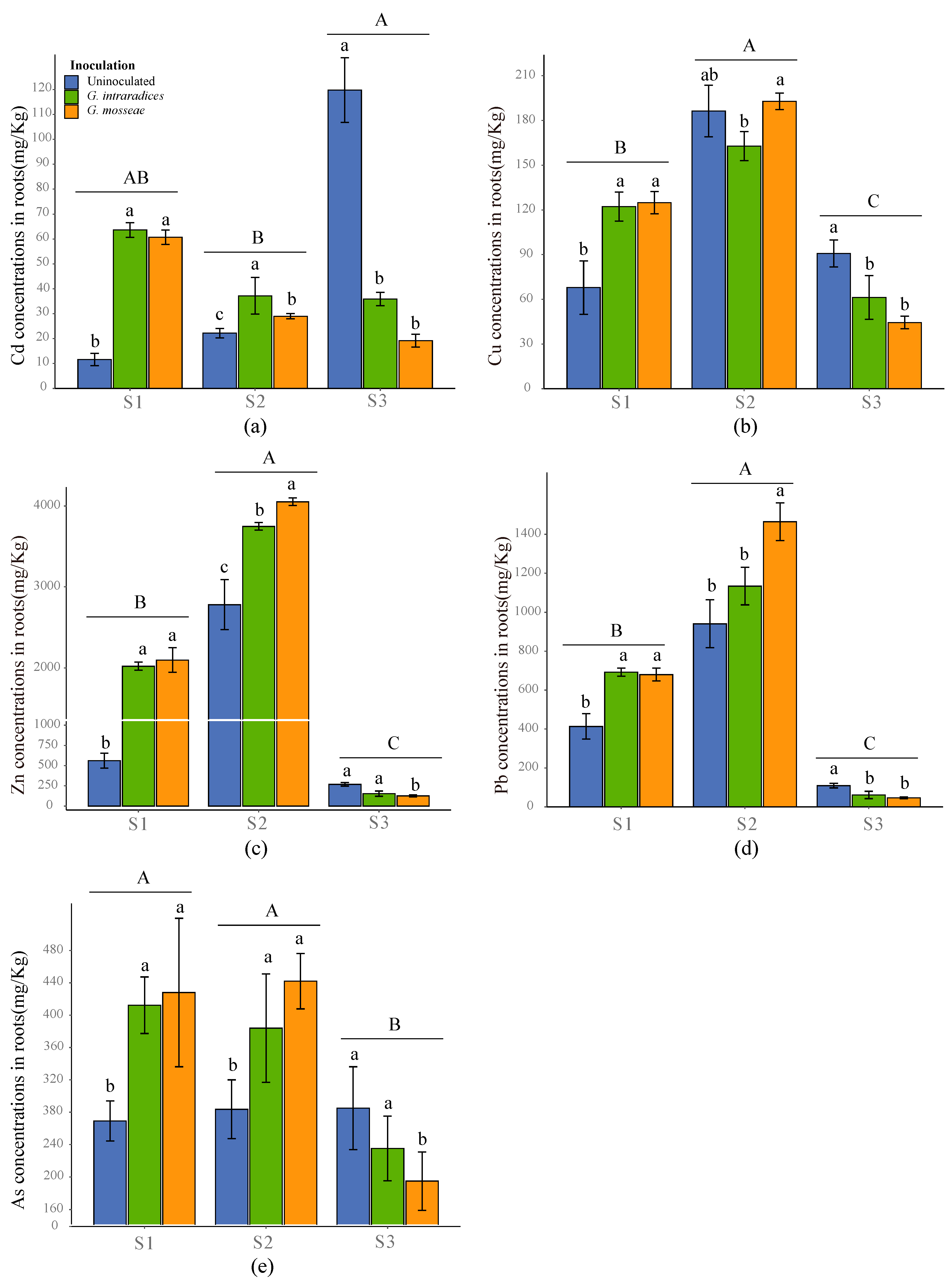
3.4. Uptake and Accumulation of Heavy Metals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, J.; Zhang, J.; Ren, L.; Zhou, Y.; Gao, J.; Luo, L.; Yang, Y.; Peng, Q.; Huang, H.; Chen, A. Diagnosis of Soil Contamination Using Microbiological Indices: A Review on Heavy Metal Pollution. J. Environ. Manag. 2019, 242, 121–130. [Google Scholar] [CrossRef]
- Khan, S.; Cao, Q.; Zheng, Y.M.; Huang, Y.Z.; Zhu, Y.G. Health Risks of Heavy Metals in Contaminated Soils and Food Crops Irrigated with Wastewater in Beijing, China. Environ. Pollut. 2008, 152, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Ren, B.; Hursthouse, A.; Deng, R.; Wang, Z. Distribution, Source Identification, and Ecological-Health Risks of Potentially Toxic Elements (PTEs) in Soil of Thallium Mine Area (Southwestern Guizhou, China). Environ. Sci. Pollut. Res. 2019, 26, 16556–16567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Wu, P.; Yang, F.; Sun, D.; Zhang, D.-X.; Zhou, Y.-K. Assessment of Heavy Metal Pollution and Human Health Risks in Urban Soils around an Electronics Manufacturing Facility. Sci. Total Environ. 2018, 630, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Douay, F.; Roussel, H.; Pruvot, C.; Loriette, A.; Fourrier, H. Assessment of a Remediation Technique Using the Replacement of Contaminated Soils in Kitchen Gardens Nearby a Former Lead Smelter in Northern France. Sci. Total Environ. 2008, 401, 29–38. [Google Scholar] [CrossRef]
- Saxena, G.; Purchase, D.; Mulla, S.I.; Saratale, G.D.; Bharagava, R.N. Phytoremediation of Heavy Metal-Contaminated Sites: Eco-Environmental Concerns, Field Studies, Sustainability Issues, and Future Prospects. In Reviews of Environmental Contamination and Toxicology; De Voogt, P., Ed.; Reviews of Environmental Contamination and Toxicology; Springer International Publishing: Cham, Switzerland, 2019; Volume 249, pp. 71–131. ISBN 978-3-030-20193-7. [Google Scholar]
- Vamerali, T.; Bandiera, M.; Mosca, G. Field Crops for Phytoremediation of Metal-Contaminated Land. A Review. Environ. Chem. Lett. 2010, 8, 1–17. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, J.; Huang, Z.; Li, H.; Liu, X.; Huang, J.; Zhuo, R.; Wu, Z.; Qin, X.; Gao, Y.; et al. Enhanced Cd Phytostabilization and Rhizosphere Bacterial Diversity of Robinia pseudoacacia L. by Endophyte Enterobacter sp. YG-14 Combined with Sludge Biochar. Sci. Total Environ. 2021, 787, 147660. [Google Scholar] [CrossRef]
- Das, S.; Chou, M.-L.; Jean, J.-S.; Yang, H.-J.; Kim, P.J. Arsenic-Enrichment Enhanced Root Exudates and Altered Rhizosphere Microbial Communities and Activities in Hyperaccumulator Pteris vittata. J. Hazard. Mater. 2017, 325, 279–287. [Google Scholar] [CrossRef]
- Hu, J.; Wu, S.; Wu, F.; Leung, H.M.; Lin, X.; Wong, M.H. Arbuscular Mycorrhizal Fungi Enhance Both Absorption and Stabilization of Cd by Alfred Stonecrop (Sedum alfredii Hance) and Perennial Ryegrass (Lolium perenne L.) in a Cd-Contaminated Acidic Soil. Chemosphere 2013, 93, 1359–1365. [Google Scholar] [CrossRef]
- Xiao, Z.; Zou, D.; Zeng, X.; Zhang, L.; Liu, F.; Wang, A.; Zeng, Q.; Zhang, G.; Li, L. Cadmium Accumulation in Oilseed Rape Is Promoted by Intercropping with Faba Bean and Ryegrass. Ecotoxicol. Environ. Saf. 2020, 205, 111162. [Google Scholar] [CrossRef]
- Filote, C.; Roșca, M.; Hlihor, R.; Cozma, P.; Simion, I.; Apostol, M.; Gavrilescu, M. Sustainable Application of Biosorption and Bioaccumulation of Persistent Pollutants in Wastewater Treatment: Current Practice. Processes 2021, 9, 1696. [Google Scholar] [CrossRef]
- Vamerali, T.; Barion, G.; Hewidy, M.; Mosca, G. Soybean Isoflavone Patterns in Main Stem and Branches as Affected by Water and Nitrogen Supply. Eur. J. Agron. 2012, 41, 1–10. [Google Scholar] [CrossRef]
- Wu, B.; Luo, H.; Wang, X.; Liu, H.; Peng, H.; Sheng, M.; Xu, F.; Xu, H. Effects of Environmental Factors on Soil Bacterial Community Structure and Diversity in Different Contaminated Districts of Southwest China Mine Tailings. Sci. Total Environ. 2022, 802, 149899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bian, F.; Zhong, Z.; Gai, X.; Yang, C. Deciphering the Rhizosphere Microbiome of a Bamboo Plant in Response to Different Chromium Contamination Levels. J. Hazard. Mater. 2020, 399, 123107. [Google Scholar] [CrossRef]
- Khalid, M.; Ur-Rahman, S.; Hassani, D.; Hayat, K.; Zhou, P.; Hui, N. Advances in Fungal-Assisted Phytoremediation of Heavy Metals: A Review. Pedosphere 2021, 31, 475–495. [Google Scholar] [CrossRef]
- Gunathilakae, N.; Yapa, N.; Hettiarachchi, R. Effect of Arbuscular Mycorrhizal Fungi on the Cadmium Phytoremediation Potential of Eichhornia crassipes (Mart.) Solms. Groundw. Sustain. Dev. 2018, 7, 477–482. [Google Scholar] [CrossRef]
- Guo, W.; Zhao, R.; Zhao, W.; Fu, R.; Guo, J.; Bi, N.; Zhang, J. Effects of Arbuscular Mycorrhizal Fungi on Maize (Zea mays L.) and Sorghum (Sorghum bicolor L. Moench) Grown in Rare Earth Elements of Mine Tailings. Appl. Soil Ecol. 2013, 72, 85–92. [Google Scholar] [CrossRef]
- Hu, Z.-H.; Zhuo, F.; Jing, S.-H.; Li, X.; Yan, T.-X.; Lei, L.-L.; Lu, R.-R.; Zhang, X.-F.; Jing, Y.-X. Combined Application of Arbuscular Mycorrhizal Fungi and Steel Slag Improves Plant Growth and Reduces Cd, Pb Accumulation in Zea mays. Int. J. Phytoremediat. 2019, 21, 857–865. [Google Scholar] [CrossRef]
- Jamal, A.; Ayub, N.; Usman, M.; Khan, A.G. Arbuscular Mycorrhizal Fungi Enhance Zinc and Nickel Uptake from Contaminated Soil by Soybean and Lentil. Int. J. Phytoremediat. 2002, 4, 205–221. [Google Scholar] [CrossRef]
- Jiang, Q.-Y.; Tan, S.-Y.; Zhuo, F.; Yang, D.-J.; Ye, Z.-H.; Jing, Y.-X. Effect of Funneliformis Mosseae on the Growth, Cadmium Accumulation and Antioxidant Activities of Solanum Nigrum. Appl. Soil Ecol. 2016, 98, 112–120. [Google Scholar] [CrossRef]
- Vlachodimos, K.; Papatheodorou, E.M.; Diamantopoulos, J.; Monokrousos, N. Assessment of Robinia pseudoacacia Cultivations as a Restoration Strategy for Reclaimed Mine Spoil Heaps. Environ. Monit. Assess 2013, 185, 6921–6932. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Zhang, H.; Song, Y.; Chen, H.; Tang, M. Funneliformis mosseae Enhances Root Development and Pb Phytostabilization in Robinia pseudoacacia in Pb-Contaminated Soil. Front. Microbiol. 2019, 10, 2591. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Jia, X.; Zhao, Y.; Chang, Y.; Bai, B. Response of Robinia pseudoacacia L. Rhizosphere Microenvironment to Cd and Pb Contamination and Elevated Temperature. Appl. Soil Ecol. 2016, 108, 269–277. [Google Scholar] [CrossRef]
- Yang, Y.; Song, Y.; Scheller, H.V.; Ghosh, A.; Ban, Y.; Chen, H.; Tang, M. Community Structure of Arbuscular Mycorrhizal Fungi Associated with Robinia pseudoacacia in Uncontaminated and Heavy Metal Contaminated Soils. Soil Biol. Biochem. 2015, 86, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhao, R.; Fu, R.; Bi, N.; Wang, L.; Zhao, W.; Guo, J.; Zhang, J. Contribution of Arbuscular Mycorrhizal Fungi to the Development of Maize (Zea mays L.) Grown in Three Types of Coal Mine Spoils. Environ. Sci. Pollut. Res. 2014, 21, 3592–3603. [Google Scholar] [CrossRef]
- Zarei, M.; König, S.; Hempel, S.; Nekouei, M.K.; Savaghebi, G.; Buscot, F. Community Structure of Arbuscular Mycorrhizal Fungi Associated to Veronica rechingeri at the Anguran Zinc and Lead Mining Region. Environ. Pollut. 2008, 156, 1277–1283. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular Mycorrhizal Fungi-Induced Mitigation of Heavy Metal Phytotoxicity in Metal Contaminated Soils: A Critical Review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Jin, H.-R.; Jiang, D.-H.; Zhang, P.-H. Effect of Carbon and Nitrogen Availability on Metabolism of Amino Acids in Germinating Spores of Arbuscular Mycorrhizal Fungi. Pedosphere 2011, 21, 432–442. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; Ent, A. A Global Database for Plants That Hyperaccumulate Metal and Metalloid Trace Elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Xi, L.; Shen, Y.; Zhao, X.; Zhou, M.; Mi, Y.; Li, X.; Chen, H.; Wei, Y.; Su, H.; Hou, H. Effects of Arbuscular Mycorrhizal Fungi on Frond Antimony Enrichment, Morphology, and Proteomics in Pteris cretica Var. Nervosa during Antimony Phytoremediation. Sci. Total Environ. 2022, 804, 149904. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, J.; Xia, X.; Chu, J.; Wei, Y.; Shi, S.; Chang, E.; Yin, W.; Jiang, Z. The Evaluation of Heavy Metal Accumulation and Application of a Comprehensive Bio-Concentration Index for Woody Species on Contaminated Sites in Hunan, China. Environ. Sci. Pollut. Res. 2014, 21, 5076–5085. [Google Scholar] [CrossRef]
- Kalembasa, S.J.; Jenkinson, D.S. A Comparative Study of Titrimetric and Gravimetric Methods for the Determination of Organic Carbon in Soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of Total, Organic, and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Phillips, C.W.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular–Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Geogr. J. 1970, 136, 158. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Wei, J.-L.; Lai, H.-Y.; Chen, Z.-S. Chelator Effects on Bioconcentration and Translocation of Cadmium by Hyperaccumulators, Tagetes patula and Impatiens walleriana. Ecotoxicol. Environ. Saf. 2012, 84, 173–178. [Google Scholar] [CrossRef]
- Ding, W.; Cong, W.-F.; Lambers, H. Plant Phosphorus-Acquisition and -Use Strategies Affect Soil Carbon Cycling. Trends Ecol. Evol. 2021, 36, 899–906. [Google Scholar] [CrossRef]
- Boilard, G.; Bradley, R.L.; Paterson, E.; Sim, A.; Brown, L.K.; George, T.S.; Bainard, L.; Carubba, A. Interaction between Root Hairs and Soil Phosphorus on Rhizosphere Priming of Soil Organic Matter. Soil Biol. Biochem. 2019, 135, 264–266. [Google Scholar] [CrossRef]
- Veresoglou, S.D.; Sen, R.; Mamolos, A.P.; Veresoglou, D.S. Plant Species Identity and Arbuscular Mycorrhizal Status Modulate Potential Nitrification Rates in Nitrogen-Limited Grassland Soils: Mycotrophic Plants Repress Nitrifiers. J. Ecol. 2011, 99, 1339–1349. [Google Scholar] [CrossRef]
- Visen, A.; Bohra, M.; Singh, P.N.; Srivastava, P.C.; Kumar, S.; Sharma, A.K.; Chakraborty, B. Two Pseudomonad Strains Facilitate AMF Mycorrhization of Litchi (Litchi chinensis Sonn.) and Improving Phosphorus Uptake. Rhizosphere 2017, 3, 196–202. [Google Scholar] [CrossRef]
- Zhao, R.; Guo, W.; Bi, N.; Guo, J.; Wang, L.; Zhao, J.; Zhang, J. Arbuscular Mycorrhizal Fungi Affect the Growth, Nutrient Uptake and Water Status of Maize (Zea mays L.) Grown in Two Types of Coal Mine Spoils under Drought Stress. Appl. Soil Ecol. 2015, 88, 41–49. [Google Scholar] [CrossRef]
- Ahmed, F.R.S.; Alexander, I.J.; Mwinyihija, M.; Killham, K. Effect of Superphosphate and Arbuscular Mycorrhizal Fungus Glomus Mosseae on Phosphorus and Arsenic Uptake in Lentil (Lens culinaris L.). Water Air Soil Pollut. 2011, 221, 169–182. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, B.; Christie, P.; Smith, F.A.; Wang, Y.; Li, X. Arsenic Uptake by Arbuscular Mycorrhizal Maize (Zea mays L.) Grown in an Arsenic-Contaminated Soil with Added Phosphorus. J. Environ. Sci. 2007, 19, 1245–1251. [Google Scholar] [CrossRef]
- Weyens, N.; Van Der Lelie, D.; Artois, T.; Smeets, K.; Taghavi, S.; Newman, L.; Carleer, R.; Vangronsveld, J. Bioaugmentation with Engineered Endophytic Bacteria Improves Contaminant Fate in Phytoremediation. Environ. Sci. Technol. 2009, 43, 9413–9418. [Google Scholar] [CrossRef]
- Hassan, S.E.; Hijri, M.; St-Arnaud, M. Effect of Arbuscular Mycorrhizal Fungi on Trace Metal Uptake by Sunflower Plants Grown on Cadmium Contaminated Soil. New Biotechnol. 2013, 30, 780–787. [Google Scholar] [CrossRef]
- Islam, E.; Yang, X.; Li, T.; Liu, D.; Jin, X.; Meng, F. Effect of Pb Toxicity on Root Morphology, Physiology and Ultrastructure in the Two Ecotypes of Elsholtzia argyi. J. Hazard. Mater. 2007, 147, 806–816. [Google Scholar] [CrossRef]
- Wan, J.; Zeng, G.; Huang, D.; Hu, L.; Xu, P.; Huang, C.; Deng, R.; Xue, W.; Lai, C.; Zhou, C.; et al. Rhamnolipid Stabilized Nano-Chlorapatite: Synthesis and Enhancement Effect on Pb-and Cd-Immobilization in Polluted Sediment. J. Hazard. Mater. 2018, 343, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Sudová, R.; Vosátka, M. Differences in the Effects of Three Arbuscular Mycorrhizal Fungal Strains on P and Pb Accumulation by Maize Plants. Plant Soil 2007, 296, 77–83. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Lou, X.; Tang, M. Mycorrhizal and Non-Mycorrhizal Medicago truncatula Roots Exhibit Differentially Regulated NADPH Oxidase and Antioxidant Response under Pb Stress. Environ. Exp. Bot. 2019, 164, 10–19. [Google Scholar] [CrossRef]
- Sabra, M.; Aboulnasr, A.; Franken, P.; Perreca, E.; Wright, L.P.; Camehl, I. Beneficial Root Endophytic Fungi Increase Growth and Quality Parameters of Sweet Basil in Heavy Metal Contaminated Soil. Front. Plant Sci. 2018, 9, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.D.; Li, X.L.; Tao, H.Q.; Christie, P.; Wong, M.H. The Role of Arbuscular Mycorrhiza in Zinc Uptake by Red Clover Growing in a Calcareous Soil Spiked with Various Quantities of Zinc. Chemosphere 2003, 50, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.R.F.S.; Siqueira, J.O. Mycorrhiza and Phosphate Protection of Tropical Grass Species against Heavy Metal Toxicity in Multi-Contaminated Soil. Biol. Fertil. Soils 2008, 44, 833–841. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; He, H.; Xiao, L.; Zhong, T.; Liu, H.; Li, S.; Deng, P.; Ye, Z.; Jing, Y. Enhancement of Cd Phytoextraction by Two Amaranthus Species with Endophytic Rahnella sp. JN27. Chemosphere 2014, 103, 99–104. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.B.; MartÍnez, C.E. Copper Phytotoxicity in a Contaminated Soil: Remediation Tests with Adsorptive Materials. Environ. Sci. Technol. 2000, 34, 4386–4391. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | S1 | S2 | S3 |
|---|---|---|---|
| pH | 7.87 ± 0.09ns | 7.82 ± 0.15ns | 7.75 ± 0.23ns |
| Organic matter (mg/kg) | 12.27 ± 2.13c | 27.44 ± 3.12a | 21.85 ± 2.56b |
| Available P (mg/kg) | 6.0 ± 1.21b | 2.5 ± 0.56c | 15.0 ± 1.54a |
| Available N (mg/kg) | 10.37 ± 1.89c | 16.29 ± 1.34b | 24.61 ± 2.12a |
| Cd (mg/kg) | 5.3 ± 0.31c | 21.7 ± 2.19a | 8.4 ± 1.23b |
| Zn (mg/kg) | 1170.8 ± 78.2c | 6619.8 ± 101.2a | 2248.7 ± 88.9b |
| Pb (mg/kg) | 549.1 ± 45.1c | 831.8 ± 78.1b | 4082.2 ± 121.2a |
| Cu (mg/kg) | 58.9 ± 8.2b | 187.7 ± 15.2a | 173.2 ± 11.1ab |
| As (mg/kg) | 13.2 ± 1.2b | 24.7 ± 2.5a | 9.6 ± 0.9c |
| Substrate | Inoculation | Mycorrhizal Colonization (%) | Height (cm) | Shoot Dry Weight (g/pot) | Root Dry Weight (g/pot) |
|---|---|---|---|---|---|
| S1 | Uninoculated | 16.0 ± 3.1b | 4.82 ± 0.49b | 0.278 ± 0.015b | 0.173 ± 0.020b |
| G. intraradices | 66.0 ± 8.3a | 9.86 ± 0.41a | 1.145 ± 0.085a | 0.455 ± 0.021a | |
| G. mosseae | 58.3 ± 8.1aB | 9.21 ± 0.13aB | 1.178 ± 0.066aB | 0.418 ± 0.038aA | |
| S2 | Uninoculated | 12.1 ± 2.3c | 5.60 ± 0.12b | 0.425 ± 0.032c | 0.165 ± 0.010c |
| G. intraradices | 83.7 ± 6.4a | 10.71 ± 0.95a | 1.421 ± 0.126a | 0.536 ± 0.037a | |
| G. mosseae | 68.3 ± 6.0bA | 8.47 ± 0.41aB | 1.022 ± 0.112bB | 0.379 ± 0.009bA | |
| S3 | Uninoculated | 13.3 ± 0.5c | 12.33 ± 0.39 | 2.100 ± 0.213 | 0.254 ± 0.049 |
| G. intraradices | 25.5 ± 6.9b | 12.68 ± 0.51 | 2.418 ± 0.177 | 0.270 ± 0.054 | |
| G. mosseae | 34.5 ± 6.3aC | 12.70 ± 0.59A | 2.191 ± 0.125A | 0.258 ± 0.032B |
| Substrate | Inoculation | Cd (μg/pot) | Zn (μg/pot) | Pb (μg/pot) | Cu (μg/pot) | As (μg/pot) |
|---|---|---|---|---|---|---|
| S1 | Uninoculated | 1.99 ± 0.40b | 95.70 ± 6.66b | 70.92 ± 11.32b | 11.71 ± 3.37b | 46.56 ± 7.63b |
| G. intraradices | 28.93 ± 1.28a | 919.91 ± 43.68a | 314.88 ± 5.55a | 55.73 ± 6.61a | 187.56 ± 15.45a | |
| G. mosseae | 25.47 ± 3.40aA | 875.83 ± 75.53aB | 284.45 ± 28.60aB | 52.06 ± 1.85aB | 180.12 ± 49.47aA | |
| S2 | Uninoculated | 3.66 ± 0.40c | 458.78 ± 62.79c | 155.57 ± 26.21c | 30.65 ± 2.23c | 46.63 ± 5.09c |
| G. intraradices | 20.05 ± 4.92a | 2007.89 ± 147.74a | 604.90 ± 21.79a | 86.93 ± 1.18a | 204.38 ± 25.89a | |
| G. mosseae | 11.00 ± 0.60bB | 1537.06 ± 17.50bA | 554.95 ± 26.64bA | 73.13 ± 3.19bA | 167.69 ± 14.53bA | |
| S3 | Uninoculated | 30.23 ± 5.35a | 68.28 ± 14.12a | 27.72 ± 6.53a | 22.81 ± 2.75a | 71.16 ± 8.83a |
| G. intraradices | 9.59 ± 1.40b | 40.25 ± 2.36b | 15.91 ± 2.24b | 16.12 ± 3.25b | 62.23 ± 8.65ab | |
| G. mosseae | 4.94 ± 1.00bC | 32.27 ± 3.28bC | 11.97 ± 2.31bC | 11.52 ± 2.34bC | 49.89 ± 9.49bB |
| Substrate | Inoculation | Cd (μg/pot) | Zn (μg/pot) | Pb (μg/pot) | Cu (μg/pot) | As (μg/pot) |
|---|---|---|---|---|---|---|
| S1 | Uninoculated | 0.44 ± 0.05b | 42.71 ± 1.29b | 6.92 ± 0.68b | 2.00 ± 0.08b | 52.48 ± 5.59b |
| G. intraradices | 2.67 ± 0.21a | 185.93 ± 19.90a | 22.18 ± 4.41a | 11.48 ± 2.61a | 252.70 ± 16.06a | |
| G. mosseae | 2.72 ± 0.25aA | 182.14 ± 6.89aB | 18.99 ± 2.30aB | 8.68 1.19aB | 268.68 ± 42.93aB | |
| S2 | Uninoculated | 0.75 ± 0.14b | 106.32 ± 13.02c | 9.64 ± 1.20c | 2.67 ± 0.51c | 122.86 ± 8.78c |
| G. intraradices | 1.80 ± 0.42a | 351.25 ± 42.46a | 45.71 ± 8.17a | 12.20 ± 0.80a | 352.63 ± 109.44a | |
| G. mosseae | 0.81 ± 0.16bB | 240.37 ± 5.64bA | 24.78 ± 4.64bA | 6.50 ± 0.94bB | 211.27 ± 34.17bB | |
| S3 | Uninoculated | 2.15 ± 0.18b | 160.99 ± 14.81 | 17.88 ± 4.57b | 31.31 ± 1.16 | 312.49 ± 40.42b |
| G. intraradices | 3.38 ± 0.49a | 161.60 ± 4.78 | 24.46 ± 3.49a | 32.74 ± 1.93 | 540.19 ± 99.39a | |
| G. mosseae | 0.92 ± 0.25cAB | 169.24 ± 24.63C | 16.03 ± 2.58bB | 32.40 ± 2.96A | 653.53 ± 31.99aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Yang, T.; Zhou, J.; Peng, X. Effects of Arbuscular Mycorrhizal Fungi on Robinia pseudoacacia L. Growing on Soils Contaminated with Heavy Metals. J. Fungi 2023, 9, 684. https://doi.org/10.3390/jof9060684
Zhao L, Yang T, Zhou J, Peng X. Effects of Arbuscular Mycorrhizal Fungi on Robinia pseudoacacia L. Growing on Soils Contaminated with Heavy Metals. Journal of Fungi. 2023; 9(6):684. https://doi.org/10.3390/jof9060684
Chicago/Turabian StyleZhao, Liuhui, Tao Yang, Jinxing Zhou, and Xiawei Peng. 2023. "Effects of Arbuscular Mycorrhizal Fungi on Robinia pseudoacacia L. Growing on Soils Contaminated with Heavy Metals" Journal of Fungi 9, no. 6: 684. https://doi.org/10.3390/jof9060684