

Clorgyline Analogs Synergize with Azoles against Drug Efflux in Candida auris

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Chemicals
2.3. Agarose Diffusion Assay
2.4. Antifungal Susceptibility Testing
2.5. Checkerboard Assay
2.6. Nile Red Assay
2.7. ATPase Activity Assay
3. Results
3.1. Antifungal Susceptibilities of C. auris Clade I and II Clinical Isolates
3.2. Screening CLO Analogs for Drug Efflux Modulation Using Disk Diffusion
3.3. Antifungal Activity of Modulators on Recombinant Strains Expressing Efflux Pumps
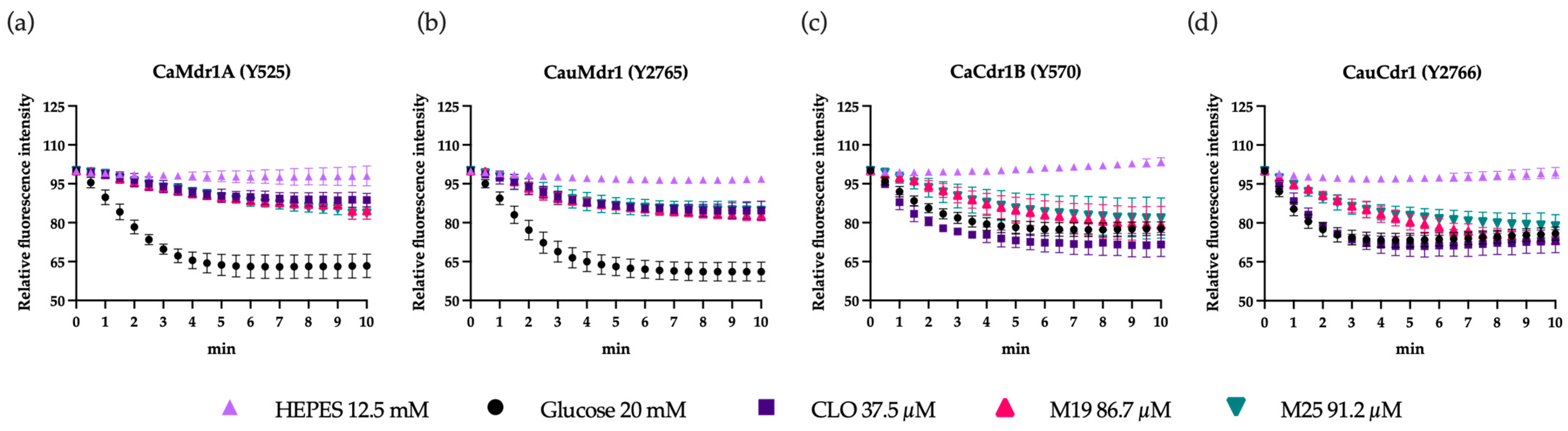
3.4. Effect of Efflux Pump Inhibitors on Recombinant Strains
3.5. Effect of the Modulators CLO, M19, and M25 on Cdr1 ATPase Activity In Vitro
3.6. Combinatorial Effect of Azoles and Modulators on Recombinant Strains Overexpressing Mdr1 and Cdr1
3.7. Combinatorial Effect of Azoles and CLO, M19, and M25 on C. auris Clinical Isolates
3.8. Effects on C. albicans Clinical Isolates of Azoles in Combination with M19 or M25
4. Discussion
4.1. Expression of Azole Resistance Determinants in C. auris Clade I and Clade II Clinical Isolates
4.2. Identification and Characterization of CLO Analogs as Candidate Modulators of Drug Efflux in Combination Therapy
4.3. Characterization of Other Potential Modulators of Drug Efflux
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 15 February 2015).
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Jacobs, J.L.; Dennis, E.K.; Taimur, S.; Rana, M.; Patel, D.; Gitman, M.; Patel, G.; Schaefer, S.; Iyer, K.; et al. Candida auris Pan-Drug-Resistant to Four Classes of Antifungal Agents. Antimicrob. Agents Chemother. 2022, 66, e00053-22. [Google Scholar] [CrossRef]
- Ostrowsky, B.; Greenko, J.; Adams, E.; Quinn, M.; O’Brien, B.; Chaturvedi, V.; Berkow, E.; Vallabhaneni, S.; Forsberg, K.; Chaturvedi, S.; et al. Candida auris Isolates Resistant to Three Classes of Antifungal Medications. MMWR Morb. Mortal. Wkly. Rep. 2019, 69, 6–9. [Google Scholar] [CrossRef]
- Chowdhary, A.; Anil Kumar, V.; Sharma, C.; Prakash, A.; Agarwal, K.; Babu, R.; Dinesh, K.R.; Karim, S.; Singh, S.K.; Hagen, F.; et al. Multidrug-resistant endemic clonal strain of Candida auris in India. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2014, 33, 919–926. [Google Scholar] [CrossRef]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control 2016, 5, 35. [Google Scholar] [CrossRef]
- Zhu, Y.; O’Brien, B.; Leach, L.; Clarke, A.; Bates, M.; Adams, E.; Ostrowsky, B.; Quinn, M.; Dufort, E.; Southwick, K.; et al. Laboratory Analysis of an Outbreak of Candida auris in New York from 2016 to 2018: Impact and Lessons Learned. J. Clin. Microbiol. 2020, 58, e01503-19. [Google Scholar] [CrossRef]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple introductions and subsequent transmission of multidrug-resistant Candida auris in the USA: A molecular epidemiological survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef]
- Huang, X.; Welsh, R.M.; Deming, C.; Proctor, D.M.; Thomas, P.J.; Gussin, G.M.; Huang, S.S.; Kong, H.H.; Bentz, M.L.; Vallabhaneni, S.; et al. Skin Metagenomic Sequence Analysis of Early Candida auris Outbreaks in U.S. Nursing Homes. mSphere 2021, 6, e0028721. [Google Scholar] [CrossRef]
- Farooqi, J.Q.; Soomro, A.S.; Baig, M.A.; Sajjad, S.F.; Hamid, K.; Jabeen, K.; Naqvi, M.F.; Nasir, N.; Roshan, R.; Mahmood, S.F.; et al. Outbreak investigation of Candida auris at a tertiary care hospital in Karachi, Pakistan. J. Infect. Prev. 2020, 21, 189–195. [Google Scholar] [CrossRef]
- Armstrong, P.A.; Rivera, S.M.; Escandon, P.; Caceres, D.H.; Chow, N.; Stuckey, M.J.; Díaz, J.; Gomez, A.; Vélez, N.; Espinosa-Bode, A.; et al. Hospital-Associated Multicenter Outbreak of Emerging Fungus Candida auris, Colombia, 2016. Emerg. Infect. Dis. 2019, 25, 1339–1346. [Google Scholar] [CrossRef]
- Nobrega de Almeida, J., Jr.; Brandão, I.B.; Francisco, E.C.; de Almeida, S.L.R.; de Oliveira Dias, P.; Pereira, F.M.; Santos Ferreira, F.; de Andrade, T.S.; de Miranda Costa, M.M.; de Souza Jordão, R.T.; et al. Axillary Digital Thermometers uplifted a multidrug-susceptible Candida auris outbreak among COVID-19 patients in Brazil. Mycoses 2021, 64, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Sabino, R.; Veríssimo, C.; Pereira, Á.A.; Antunes, F. Candida auris, an Agent of Hospital-Associated Outbreaks: Which Challenging Issues Do We Need to Have in Mind? Microorganisms 2020, 8, 181. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Duggal, S.; Agarwal, K.; Prakash, A.; Singh, P.K.; Jain, S.; Kathuria, S.; Randhawa, H.S.; Hagen, F.; et al. New clonal strain of Candida auris, Delhi, India. Emerg. Infect. Dis. 2013, 19, 1670–1673. [Google Scholar] [CrossRef]
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action. Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 15 February 2023).
- Chow, N.A.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential Fifth Clade of Candida auris, Iran, 2018. Emerg. Infect. Dis. 2019, 25, 1780–1781. [Google Scholar] [CrossRef]
- Spruijtenburg, B.; Badali, H.; Abastabar, M.; Mirhendi, H.; Khodavaisy, S.; Sharifisooraki, J.; Taghizadeh Armaki, M.; de Groot, T.; Meis, J.F. Confirmation of fifth Candida auris clade by whole genome sequencing. Emerg. Microbes Infect. 2022, 11, 2405–2411. [Google Scholar] [CrossRef]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of clinically derived mutations in ERG11 to azole resistance in Candida albicans. Antimicrob. Agents Chemother. 2015, 59, 450–460. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Koymans, L.; Bille, J. Amino acid substitutions in the cytochrome P-450 lanosterol 14alpha-demethylase (CYP51A1) from azole-resistant Candida albicans clinical isolates contribute to resistance to azole antifungal agents. Antimicrob. Agents Chemother. 1998, 42, 241–253. [Google Scholar] [CrossRef]
- Marichal, P.; Koymans, L.; Willemsens, S.; Bellens, D.; Verhasselt, P.; Luyten, W.; Borgers, M.; Ramaekers, F.C.S.; Odds, F.C.; Vanden Bossche, H. Contribution of mutations in the cytochrome P450 14α-demethylase (Erg11p, Cyp51p) to azole resistance in Candida albicans. Microbiology 1999, 145, 2701–2713. [Google Scholar] [CrossRef]
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the Evolutionary History and Global Expansion of Candida auris Using Population Genomic Analyses. mBio 2020, 11, e03364-19. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Shin, J.H.; Byun, S.A.; Choi, M.J.; Won, E.J.; Lee, D.; Lee, S.Y.; Chun, S.; Lee, J.H.; Choi, H.J.; et al. Candida auris Clinical Isolates from South Korea: Identification, Antifungal Susceptibility, and Genotyping. J. Clin. Microbiol. 2019, 57, e01624-18. [Google Scholar] [CrossRef]
- Toepfer, S.; Lackner, M.; Keniya, M.V.; Monk, B.C. Functional Expression of Recombinant Candida auris Proteins in Saccharomyces cerevisiae Enables Azole Susceptibility Evaluation and Drug Discovery. J. Fungi 2023, 9, 168. [Google Scholar] [CrossRef]
- Flowers, S.A.; Barker, K.S.; Berkow, E.L.; Toner, G.; Chadwick, S.G.; Gygax, S.E.; Morschhäuser, J.; Rogers, P.D. Gain-of-function mutations in UPC2 are a frequent cause of ERG11 upregulation in azole-resistant clinical isolates of Candida albicans. Eukaryot. Cell 2012, 11, 1289–1299. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene Duplication Associated with Increased Fluconazole Tolerance in Candida auris cells of Advanced Generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef]
- Albertson, G.D.; Niimi, M.; Cannon, R.D.; Jenkinson, H.F. Multiple efflux mechanisms are involved in Candida albicans fluconazole resistance. Antimicrob. Agents Chemother. 1996, 40, 2835–2841. [Google Scholar] [CrossRef]
- Nakamura, K.; Niimi, M.; Niimi, K.; Holmes, A.R.; Yates, J.E.; Decottignies, A.; Monk, B.C.; Goffeau, A.; Cannon, R.D. Functional expression of Candida albicans drug efflux pump Cdr1p in a Saccharomyces cerevisiae strain deficient in membrane transporters. Antimicrob. Agents Chemother. 2001, 45, 3366–3374. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Ambudkar, S.V.; Prasad, R. Substitution of threonine-1351 in the multidrug transporter Cdr1p of Candida albicans results in hypersusceptibility to antifungal agents and threonine-1351 is essential for synergic effects of calcineurin inhibitor FK520. J. Antimicrob. Chemother. 2004, 54, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; Lin, Y.H.; Niimi, K.; Lamping, E.; Keniya, M.; Niimi, M.; Tanabe, K.; Monk, B.C.; Cannon, R.D. ABC transporter Cdr1p contributes more than Cdr2p does to fluconazole efflux in fluconazole-resistant Candida albicans clinical isolates. Antimicrob. Agents Chemother. 2008, 52, 3851–3862. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, S.; Michel, S.; Morschhäuser, J. Targeted gene disruption in Candida albicans wild-type strains: The role of the MDR1 gene in fluconazole resistance of clinical Candida albicans isolates. Mol. Microbiol. 2000, 36, 856–865. [Google Scholar] [CrossRef]
- Dunkel, N.; Blass, J.; Rogers, P.D.; Morschhäuser, J. Mutations in the multi-drug resistance regulator MRR1, followed by loss of heterozygosity, are the main cause of MDR1 overexpression in fluconazole-resistant Candida albicans strains. Mol. Microbiol. 2008, 69, 827–840. [Google Scholar] [CrossRef]
- Schubert, S.; Rogers, P.D.; Morschhäuser, J. Gain-of-function mutations in the transcription factor MRR1 are responsible for overexpression of the MDR1 efflux pump in fluconazole-resistant Candida dubliniensis strains. Antimicrob. Agents Chemother. 2008, 52, 4274–4280. [Google Scholar] [CrossRef]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef]
- Nishimoto, A.T.; Sharma, C.; Rogers, P.D. Molecular and genetic basis of azole antifungal resistance in the opportunistic pathogenic fungus Candida albicans. J. Antimicrob. Chemother. 2020, 75, 257–270. [Google Scholar] [CrossRef]
- Thakur, J.K.; Arthanari, H.; Yang, F.; Pan, S.J.; Fan, X.; Breger, J.; Frueh, D.P.; Gulshan, K.; Li, D.K.; Mylonakis, E.; et al. A nuclear receptor-like pathway regulating multidrug resistance in fungi. Nature 2008, 452, 604–609. [Google Scholar] [CrossRef]
- Li, J.; Coste, A.T.; Bachmann, D.; Sanglard, D.; Lamoth, F. Deciphering the Mrr1/Mdr1 Pathway in Azole Resistance of Candida auris. Antimicrob. Agents Chemother. 2022, 66, e0006722. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Rybak, J.M.; Doorley, L.A.; Nishimoto, A.T.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Abrogation of Triazole Resistance upon Deletion of CDR1 in a Clinical Isolate of Candida auris. Antimicrob. Agents Chemother. 2019, 63, e00057-19. [Google Scholar] [CrossRef]
- Iyer, K.R.; Camara, K.; Daniel-Ivad, M.; Trilles, R.; Pimentel-Elardo, S.M.; Fossen, J.L.; Marchillo, K.; Liu, Z.; Singh, S.; Muñoz, J.F.; et al. An oxindole efflux inhibitor potentiates azoles and impairs virulence in the fungal pathogen Candida auris. Nat. Commun. 2020, 11, 6429. [Google Scholar] [CrossRef]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding Echinocandin Resistance in the Emerging Pathogen Candida auris. Antimicrob. Agents Chemother. 2018, 62, e00238-00218. [Google Scholar] [CrossRef]
- Biagi, M.J.; Wiederhold, N.P.; Gibas, C.; Wickes, B.L.; Lozano, V.; Bleasdale, S.C.; Danziger, L. Development of High-Level Echinocandin Resistance in a Patient With Recurrent Candida auris Candidemia Secondary to Chronic Candiduria. Open Forum Infect. Dis. 2019, 6, ofz262. [Google Scholar] [CrossRef]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–17) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Kumar, N.; Sharma, S.; Govil, D.; Ali, T.; Mehta, Y.; Rattan, A. Candidemia caused by amphotericin B and fluconazole resistant Candida auris. Indian J. Med. Microbiol. 2013, 31, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Manuel, R.; Brown, C.S. Candida auris: A Review of the Literature. Clin. Microbiol. Rev. 2017, 31, e00029-17. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Candida auris. Available online: https://www.cdc.gov/fungal/candida-auris/index.html (accessed on 16 February 2023).
- Liu, L.; Jiang, T.; Zhou, J.; Mei, Y.; Li, J.; Tan, J.; Wei, L.; Li, J.; Peng, Y.; Chen, C.; et al. Repurposing the FDA-approved anticancer agent ponatinib as a fluconazole potentiator by suppression of multidrug efflux and Pma1 expression in a broad spectrum of yeast species. Microb. Biotechnol. 2022, 15, 482–498. [Google Scholar] [CrossRef]
- Lu, M.; Yan, H.; Yu, C.; Yuan, L.; Sun, S. Proton pump inhibitors act synergistically with fluconazole against resistant Candida albicans. Sci. Rep. 2020, 10, 498. [Google Scholar] [CrossRef]
- Holmes, A.R.; Keniya, M.V.; Ivnitski-Steele, I.; Monk, B.C.; Lamping, E.; Sklar, L.A.; Cannon, R.D. The Monoamine Oxidase A Inhibitor Clorgyline Is a Broad-Spectrum Inhibitor of Fungal ABC and MFS Transporter Efflux Pump Activities Which Reverses the Azole Resistance of Candida albicans and Candida glabrata Clinical Isolates. Antimicrob. Agents Chemother. 2012, 56, 1508–1515. [Google Scholar] [CrossRef]
- Eldesouky, H.E.; Salama, E.A.; Hazbun, T.R.; Mayhoub, A.S.; Seleem, M.N. Ospemifene displays broad-spectrum synergistic interactions with itraconazole through potent interference with fungal efflux activities. Sci. Rep. 2020, 10, 6089. [Google Scholar] [CrossRef]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef]
- Baddley, J.W.; Pappas, P.G. Antifungal combination therapy: Clinical potential. Drugs 2005, 65, 1461–1480. [Google Scholar] [CrossRef]
- Ostrosky-Zeichner, L. Combination antifungal therapy: A critical review of the evidence. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2008, 14 (Suppl. S4), 65–70. [Google Scholar] [CrossRef]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef]
- Monk, B.C.; Keniya, M.V. Using Yeast to Discover Inhibitors of Multidrug Efflux in Candida albicans. In Candida albicans: Cellular and Molecular Biology; Prasad, R., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 491–543. [Google Scholar]
- Fakhim, H.; Chowdhary, A.; Prakash, A.; Vaezi, A.; Dannaoui, E.; Meis, J.F.; Badali, H. In Vitro Interactions of Echinocandins with Triazoles against Multidrug-Resistant Candida auris. Antimicrob. Agents Chemother. 2017, 61, e01056-17. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, B.; Chaturvedi, S.; Chaturvedi, V. In Vitro Evaluation of Antifungal Drug Combinations against Multidrug-Resistant Candida auris Isolates from New York Outbreak. Antimicrob. Agents Chemother. 2020, 64, e02195-19. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Deshpande, L.M.; Rhomberg, P.R.; Utt, E.A.; Castanheira, M. Evaluation of Synergistic Activity of Isavuconazole or Voriconazole plus Anidulafungin and the Occurrence and Genetic Characterization of Candida auris Detected in a Surveillance Program. Antimicrob. Agents Chemother. 2021, 65, e02031-20. [Google Scholar] [CrossRef]
- Caballero, U.; Kim, S.; Eraso, E.; Quindós, G.; Vozmediano, V.; Schmidt, S.; Jauregizar, N. In Vitro Synergistic Interactions of Isavuconazole and Echinocandins against Candida auris. Antibiotics 2021, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Bidaud, A.L.; Botterel, F.; Chowdhary, A.; Dannaoui, E. In vitro antifungal combination of flucytosine with amphotericin B, voriconazole, or micafungin against Candida auris shows no antagonism. Antimicrob. Agents Chemother. 2019, 63, e01393-19. [Google Scholar] [CrossRef]
- Eldesouky, H.E.; Li, X.; Abutaleb, N.S.; Mohammad, H.; Seleem, M.N. Synergistic interactions of sulfamethoxazole and azole antifungal drugs against emerging multidrug-resistant Candida auris. Int. J. Antimicrob. Agents 2018, 52, 754–761. [Google Scholar] [CrossRef]
- Wu, Y.; Totten, M.; Memon, W.; Ying, C.; Zhang, S.X. In Vitro Antifungal Susceptibility of the Emerging Multidrug-Resistant Pathogen Candida auris to Miltefosine Alone and in Combination with Amphotericin B. Antimicrob. Agents Chemother. 2020, 64, e02063-02019. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Salama, E.A.; Lanman, N.A.; Hazbun, T.R.; Seleem, M.N. Potent Synergistic Interactions between Lopinavir and Azole Antifungal Drugs against Emerging Multidrug-Resistant Candida auris. Antimicrob. Agents Chemother. 2020, 65, e00684-20. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, P.; Bidaud, A.L.; Dannaoui, E. In vitro synergy of isavuconazole in combination with colistin against Candida auris. Sci. Rep. 2020, 10, 21448. [Google Scholar] [CrossRef]
- Bidaud, A.L.; Djenontin, E.; Botterel, F.; Chowdhary, A.; Dannaoui, E. Colistin interacts synergistically with echinocandins against Candida auris. Int. J. Antimicrob. Agents 2020, 55, 105901. [Google Scholar] [CrossRef]
- Eldesouky, H.E.; Lanman, N.A.; Hazbun, T.R.; Seleem, M.N. Aprepitant, an antiemetic agent, interferes with metal ion homeostasis of Candida auris and displays potent synergistic interactions with azole drugs. Virulence 2020, 11, 1466–1481. [Google Scholar] [CrossRef] [PubMed]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Monk, B.C.; Tyndall, J.D.A. Structural Insights into Binding of the Antifungal Drug Fluconazole to Saccharomyces cerevisiae Lanosterol 14α-Demethylase. Antimicrob. Agents Chemother. 2015, 59, 4982–4989. [Google Scholar] [CrossRef]
- Lamping, E.; Monk, B.C.; Niimi, K.; Holmes, A.R.; Tsao, S.; Tanabe, K.; Niimi, M.; Uehara, Y.; Cannon, R.D. Characterization of three classes of membrane proteins involved in fungal azole resistance by functional hyperexpression in Saccharomyces cerevisiae. Eukaryot. Cell 2007, 6, 1150–1165. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Lyons, C.N.; Ha, K.; Rustad, T.R.; White, T.C. Inducible azole resistance associated with a heterogeneous phenotype in Candida albicans. Antimicrob. Agents Chemother. 2001, 45, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.; Miller, S.M.; Kurtz, M.B.; Kirsch, D.R. Directed mutagenesis in Candida albicans: One-step gene disruption to isolate ura3 mutants. Mol. Cell. Biol. 1987, 7, 199–208. [Google Scholar] [CrossRef]
- Ohmomo, Y.; Hirata, M.; Murakami, K.; Magata, Y.; Tanaka, C.; Yokoyama, A. Synthesis and characterization of 11C-labeled fluoroclorgyline: A monoamine oxidase A specific inhibitor for positron emission tomography. Chem. Pharm. Bull. 1993, 41, 1994–1997. [Google Scholar] [CrossRef]
- Leonardi, A.; Barlocco, D.; Montesano, F.; Cignarella, G.; Motta, G.; Testa, R.; Poggesi, E.; Seeber, M.; De Benedetti, P.G.; Fanelli, F. Synthesis, screening, and molecular modeling of new potent and selective antagonists at the alpha 1d adrenergic receptor. J. Med. Chem. 2004, 47, 1900–1918. [Google Scholar] [CrossRef]
- Monk, B.C.; Mason, A.B.; Abramochkin, G.; Haber, J.E.; Seto-Young, D.; Perlin, D.S. The yeast plasma membrane proton pumping ATPase is a viable antifungal target. I. Effects of the cysteine-modifying reagent omeprazole. Biochim. Biophys. Acta 1995, 1239, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.N.; Köhler, J.R. Antagonism of Fluconazole and a Proton Pump Inhibitor against Candida albicans. Antimicrob. Agents Chemother. 2016, 60, 1145–1147. [Google Scholar] [CrossRef]
- Keniya, M.V.; Fleischer, E.; Klinger, A.; Cannon, R.D.; Monk, B.C. Inhibitors of the Candida albicans Major Facilitator Superfamily Transporter Mdr1p Responsible for Fluconazole Resistance. PLoS ONE 2015, 10, e0126350. [Google Scholar] [CrossRef]
- EUCAS. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7.3.2_Yeast_testing_definitive_revised_2020.pdf (accessed on 18 August 2020).
- Loewe, S. The problem of synergism and antagonism of combined drugs. Arzneimittelforschung 1953, 3, 285–290. [Google Scholar]
- Johnson, M.D.; MacDougall, C.; Ostrosky-Zeichner, L.; Perfect, J.R.; Rex, J.H. Combination antifungal therapy. Antimicrob. Agents Chemother. 2004, 48, 693–715. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Ivnitski-Steele, I.; Holmes, A.R.; Lamping, E.; Monk, B.C.; Cannon, R.D.; Sklar, L.A. Identification of Nile red as a fluorescent substrate of the Candida albicans ATP-binding cassette transporters Cdr1p and Cdr2p and the major facilitator superfamily transporter Mdr1p. Anal. Biochem. 2009, 394, 87–91. [Google Scholar] [CrossRef]
- Madani, G.; Lamping, E.; Cannon, R.D. Engineering a Cysteine-Deficient Functional Candida albicans Cdr1 Molecule Reveals a Conserved Region at the Cytosolic Apex of ABCG Transporters Important for Correct Folding and Trafficking of Cdr1. mSphere 2021, 6, e01318-20. [Google Scholar] [CrossRef]
- Madani, G.; Lamping, E.; Lee, H.J.; Niimi, M.; Mitra, A.K.; Cannon, R.D. Small-Scale Plasma Membrane Preparation for the Analysis of Candida albicans Cdr1-mGFPHis. J. Vis. Exp. JoVE 2021, 172, e62592. [Google Scholar] [CrossRef]
- Monk, B.C.; Kurtz, M.B.; Marrinan, J.A.; Perlin, D.S. Cloning and characterization of the plasma membrane H(+)-ATPase from Candida albicans. J. Bacteriol. 1991, 173, 6826–6836. [Google Scholar] [CrossRef] [PubMed]
- CDC. Antifungal Susceptibility Testing and Interpretation (Candida auris). Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 19 January 2023).
- Healey, K.R.; Kordalewska, M.; Jiménez Ortigosa, C.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mutations Identified in Isolates of Candida auris Directly Contribute to Reduced Azole Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e01427-18. [Google Scholar] [CrossRef] [PubMed]
- Keniya, M.V.; Ruma, Y.N.; Tyndall, J.D.A.; Monk, B.C. Heterologous Expression of Full-Length Lanosterol 14α-Demethylases of Prominent Fungal Pathogens Candida albicans and Candida glabrata Provides Tools for Antifungal Discovery. Antimicrob. Agents Chemother. 2018, 62, e01131-18. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric studies of the lipid probe, nile red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-mediated antifungal drug resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef]
- Bock, C.; Lengauer, T. Managing drug resistance in cancer: Lessons from HIV therapy. Nat. Rev. Cancer 2012, 12, 494–501. [Google Scholar] [CrossRef]
- Eastman, R.T.; Fidock, D.A. Artemisinin-based combination therapies: A vital tool in efforts to eliminate malaria. Nat. Rev. Microbiol. 2009, 7, 864–874. [Google Scholar] [CrossRef]
- Zumla, A.; Hafner, R.; Lienhardt, C.; Hoelscher, M.; Nunn, A. Advancing the development of tuberculosis therapy. Nat. Rev. Drug Discov. 2012, 11, 171–172. [Google Scholar] [CrossRef]
- Hill, J.A.; Cowen, L.E. Using combination therapy to thwart drug resistance. Future Microbiol. 2015, 10, 1719–1726. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- WHO. Guidelines for Diagnosing, Preventing and Managing cryptococcal Disease among Adults, Adolescents and Children Living with HIV. Available online: https://www.who.int/publications/i/item/9789240052178 (accessed on 22 April 2023).
- Ma, J.; Yoshimura, M.; Yamashita, E.; Nakagawa, A.; Ito, A.; Tsukihara, T. Structure of rat monoamine oxidase A and its specific recognitions for substrates and inhibitors. J. Mol. Biol. 2004, 338, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Milczek, E.M.; Bonivento, D.; Binda, C.; Mattevi, A.; McDonald, I.A.; Edmondson, D.E. Structural and mechanistic studies of mofegiline inhibition of recombinant human monoamine oxidase B. J. Med. Chem. 2008, 51, 8019–8026. [Google Scholar] [CrossRef]
- Wichitnithad, W.; O’Callaghan, J.P.; Miller, D.B.; Train, B.C.; Callery, P.S. Time-dependent slowly-reversible inhibition of monoamine oxidase A by N-substituted 1,2,3,6-tetrahydropyridines. Bioorg. Med. Chem. 2011, 19, 7482–7492. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.B.; Zieske, P.A. 1-Benzylcyclopropylamine and 1-(phenylcyclopropyl)methylamine: An inactivator and a substrate of monoamine oxidase. J. Med. Chem. 1985, 28, 1953–1957. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.H.; Rawal, M.K.; Dhamgaye, S.; Komath, S.S.; Saxena, A.K.; Prasad, R. Mutational Analysis of Intracellular Loops Identify Cross Talk with Nucleotide Binding Domains of Yeast ABC Transporter Cdr1p. Sci. Rep. 2015, 5, 11211. [Google Scholar] [CrossRef] [PubMed]
- Ernst, R.; Kueppers, P.; Klein, C.M.; Schwarzmueller, T.; Kuchler, K.; Schmitt, L. A mutation of the H-loop selectively affects rhodamine transport by the yeast multidrug ABC transporter Pdr5. Proc. Natl. Acad. Sci. USA 2008, 105, 5069–5074. [Google Scholar] [CrossRef]
- Banerjee, A.; Pata, J.; Sharma, S.; Monk, B.C.; Falson, P.; Prasad, R. Directed Mutational Strategies Reveal Drug Binding and Transport by the MDR Transporters of Candida albicans. J. Fungi 2021, 7, 68. [Google Scholar] [CrossRef]
- Shukla, S.; Rai, V.; Banerjee, D.; Prasad, R. Characterization of Cdr1p, a major multidrug efflux protein of Candida albicans: Purified protein is amenable to intrinsic fluorescence analysis. Biochemistry 2006, 45, 2425–2435. [Google Scholar] [CrossRef]
- Spitzer, M.; Robbins, N.; Wright, G.D. Combinatorial strategies for combating invasive fungal infections. Virulence 2017, 8, 169–185. [Google Scholar] [CrossRef]
- El-Ganiny, A.M.; Kamel, H.A.; Yossef, N.E.; Mansour, B.; El-Baz, A.M. Repurposing pantoprazole and haloperidol as efflux pump inhibitors in azole resistant clinical Candida albicans and non-albicans isolates. Saudi Pharm. J. 2022, 30, 245–255. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Description | Reference/Source |
|---|---|---|---|
| Y1857 (ADΔΔ) | MATα pdr1-3 his1 Δyor1::hisG Δsnq2::hisG Δpdr10::hisG Δpdr11::hisG Δycf1::hisG Δpdr3::hisG Δpdr5::hisG Δpdr15::hisG, Δura3 Δhis1::dpl2000 | S. cerevisiae host | [69] |
| Y525 (CaMdr1A) | AD Δpdr5::CaMDR1A | C. albicans Mdr1A overexpressed | [70] |
| Y570 (CaCdr1B) | AD Δpdr5::CaCDR1B | C. albicans Cdr1B overexpressed | [70] |
| Y2765 (CauMdr1) | ADΔΔ Δpdr5::CauMDR1 | C. auris Mdr1 overexpressed | [25] |
| Y2766 (CauCdr1) | ADΔΔ Δpdr5::CauCDR1 | C. auris Cdr1 overexpressed | [25] |
| Y2767 (CauErg11) | ADΔΔ Δpdr5::CauERG11 | C. auris Erg11 overexpressed | [25] |
| CBS10913T | Wild type, clade II | C. auris clinical isolate | Westerdijk Fungal Biodiversity Institute |
| CBS12875 | Erg11 Y132F, clade I | C. auris clinical isolate | Westerdijk Fungal Biodiversity Institute |
| AR0389 | Erg11 Y132F, CDR1 overexpression, clade I | C. auris clinical isolate | Antimicrobial Resistance Isolate Bank |
| TL1 | FLC susceptible | C. albicans clinical isolate | [71] |
| TL3 | FLC resistant | C. albicans clinical isolate | [71] |
| SGY-243 | ade2/ade2 Δura3::ADE21/Δura3::ADE2 | Mutagenized C. albicans isolate | [72] |
| FR2 | SGY-243 passaged in FLC | C. albicans Mdr1 overexpressed | [28] |
| Strain | FLC | VRC | ITC | POS | ANA | MFG | AMB |
|---|---|---|---|---|---|---|---|
| CBS10913T | 2.0 (S) | 0.008 (n.d.) | 0.016 (n.d.) | 0.03 (n.d.) | 0.03 (S) | 0.03 (S) | 0.5 (S) |
| CBS12875 | >64.0 (R) | 2.0 (n.d.) | 0.25 (n.d.) | 0.06 (n.d.) | 0.125 (S) | 0.125 (S) | 4.0 (R) |
| AR0389 | >64.0 (R) | 4.0 (n.d.) | 0.5 (n.d.) | 0.25 (n.d.) | 0.25 (S) | 0.125 (S) | 1.0 (S) |
| Drug Class | Compound | CauErg11 | CauMdr1 | CauCdr1 | CaMdr1A | CaCdr1B | ADΔΔ |
|---|---|---|---|---|---|---|---|
| Cdr1 inhibitor | Enniatin B (ENB) | 15.8 | 15.5 | 10.7 | 16.8 | 5.4 | 14.4 |
| Beauvericin (BEA) | 11.0 | 9.0 | 6.7 | 11.8 | 11.8 | 11.2 | |
| Tyrosine kinase inhibitor | Ponatinib (PON) | 8.3 | 7.3 | 50 | 8.3 | >50 | 5.2 |
| PPI | Omeprazole (OME) | 58.3 | 315 | 1900 | 500 | >6000 | 75 |
| Rabeprazole (RAB) | 278 | 278 | >1875 | 156 | >1875 | 29 | |
| Estrogen receptor modulator | Ospemifene (OSP) | >250 | >250 | >250 | >250 | >250 | >250 |
| MAO-A inhibitor | Clorgyline (CLO) | 150 | 99 | 130 | 99 | 140 | 137 |
| Clorgyline analogs | M19 | 86.7 | 86.7 | 98.3 | 86.7 | 86.7 | 86.7 |
| M25 | 137 | 182 | 228 | 273 | 273 | 137 |
| Recombinant S. cerevisiae | ||
|---|---|---|
| Reagents | CauMdr1 Y2765 | CauCdr1 Y2766 |
| POS/CLO | NI | SYN |
| POS/M19 | NI | SYN |
| POS/M25 | SYN | SYN |
| POS/PON | NI | NI |
| POS/OSP | NI | NI |
| POS/OME | SYN | SYN |
| POS/RAB | SYN | NI |
| POS/ENB | NI | SYN |
| POS/BEA | NI | SYN |
| VRC/CLO | SYN | NI |
| VRC/M19 | NI | SYN |
| VRC/M25 | SYN | NI |
| VRC/PON | SYN | NI |
| VRC/OSP | NI | NI |
| VRC/OME | SYN | SYN |
| VRC/RAB | NI | NI |
| VRC/ENB | NI | SYN |
| VRC/BEA | NI | SYN |
| ISA/CLO | SYN | NI |
| ISA/M19 | NI | NI |
| ISA/M25 | SYN | NI |
| ISA/OSP | NI | NI |
| ISA/OME | SYN | NI |
| C. auris | |||
|---|---|---|---|
| Reagents | CBS12875 | AR0389 | CBS10913T |
| POS/CLO | NI | SYN | NI |
| POS/M19 | SYN | SYN | SYN |
| POS/M25 | SYN | SYN | SYN |
| VRC/CLO | NI | NI | NI |
| VRC/M19 | SYN | SYN | NI |
| VRC/M25 | SYN | SYN | SYN |
| ITC/CLO | NI | NI | NI |
| ITC/M19 | SYN | SYN | SYN |
| VT-1161/CLO | NI | NI | NI |
| VT-1161/M19 | SYN | SYN | SYN |
| MFG/CLO | NI | NI | NI |
| MFG/M19 | NI | NI | NI |
| C. albicans | ||||
|---|---|---|---|---|
| Reagents | TL1 | TL3 | SGY243 | FR2 |
| POS/M19 | NI | NI | NI | NI |
| POS/M25 | NI | NI | NI | NI |
| VRC/M19 | NI | NI | NI | NI |
| VRC/M25 | NI | NI | NI | NI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toepfer, S.; Lackner, M.; Keniya, M.V.; Zenz, L.-M.; Friemert, M.; Bracher, F.; Monk, B.C. Clorgyline Analogs Synergize with Azoles against Drug Efflux in Candida auris. J. Fungi 2023, 9, 663. https://doi.org/10.3390/jof9060663
Toepfer S, Lackner M, Keniya MV, Zenz L-M, Friemert M, Bracher F, Monk BC. Clorgyline Analogs Synergize with Azoles against Drug Efflux in Candida auris. Journal of Fungi. 2023; 9(6):663. https://doi.org/10.3390/jof9060663
Chicago/Turabian StyleToepfer, Stephanie, Michaela Lackner, Mikhail V. Keniya, Lisa-Maria Zenz, Marianne Friemert, Franz Bracher, and Brian C. Monk. 2023. "Clorgyline Analogs Synergize with Azoles against Drug Efflux in Candida auris" Journal of Fungi 9, no. 6: 663. https://doi.org/10.3390/jof9060663