
Insights into Diversity, Distribution, and Systematics of Rust Genus Puccinia
 , ,
, ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. The Genus Puccinia: General Taxonomy and Ecology
- Kingdom: Fungi
- Division: Basidiomycota
- Class: Pucciniomycetes
- Order: Pucciniales
- Family: Pucciniaceae
- Genus: Puccinia Pers. (1801)
- Type species: Puccinia graminis Pers. (1794)
3. Data Collection and Compilation
4. Phylogenetic Analyses
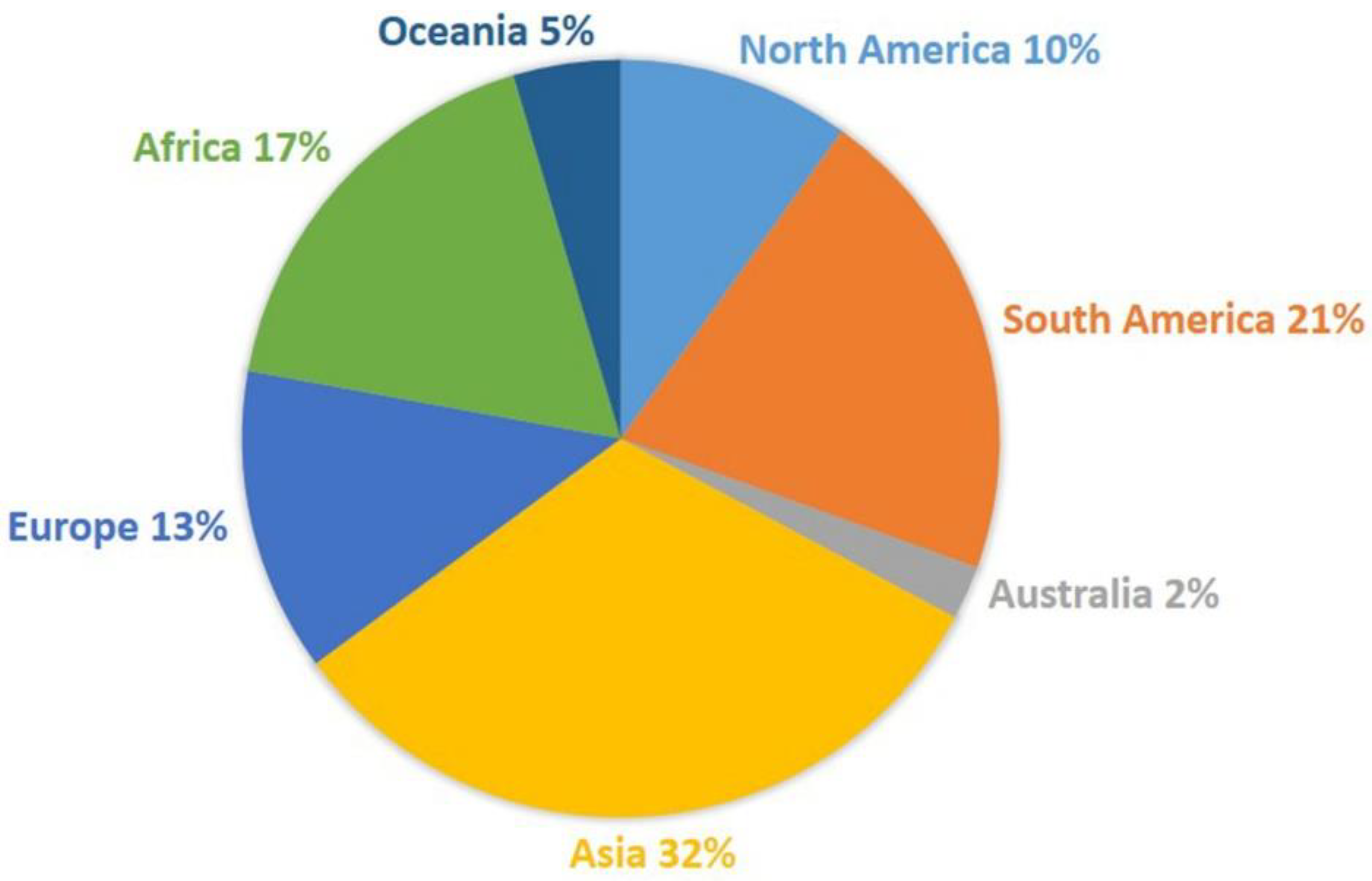
5. Current Status of Numbers of Species
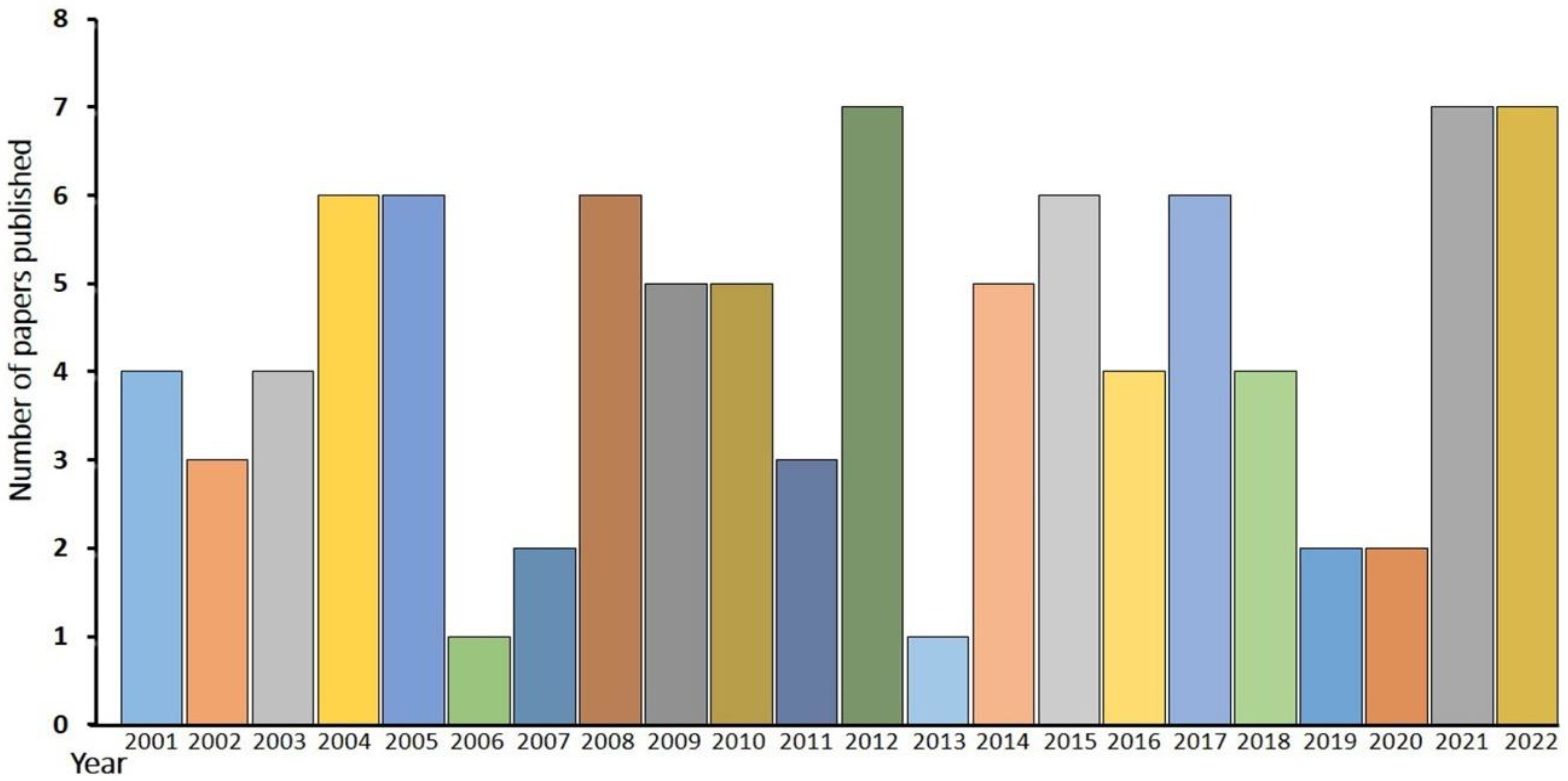
6. Trends in Published Literature
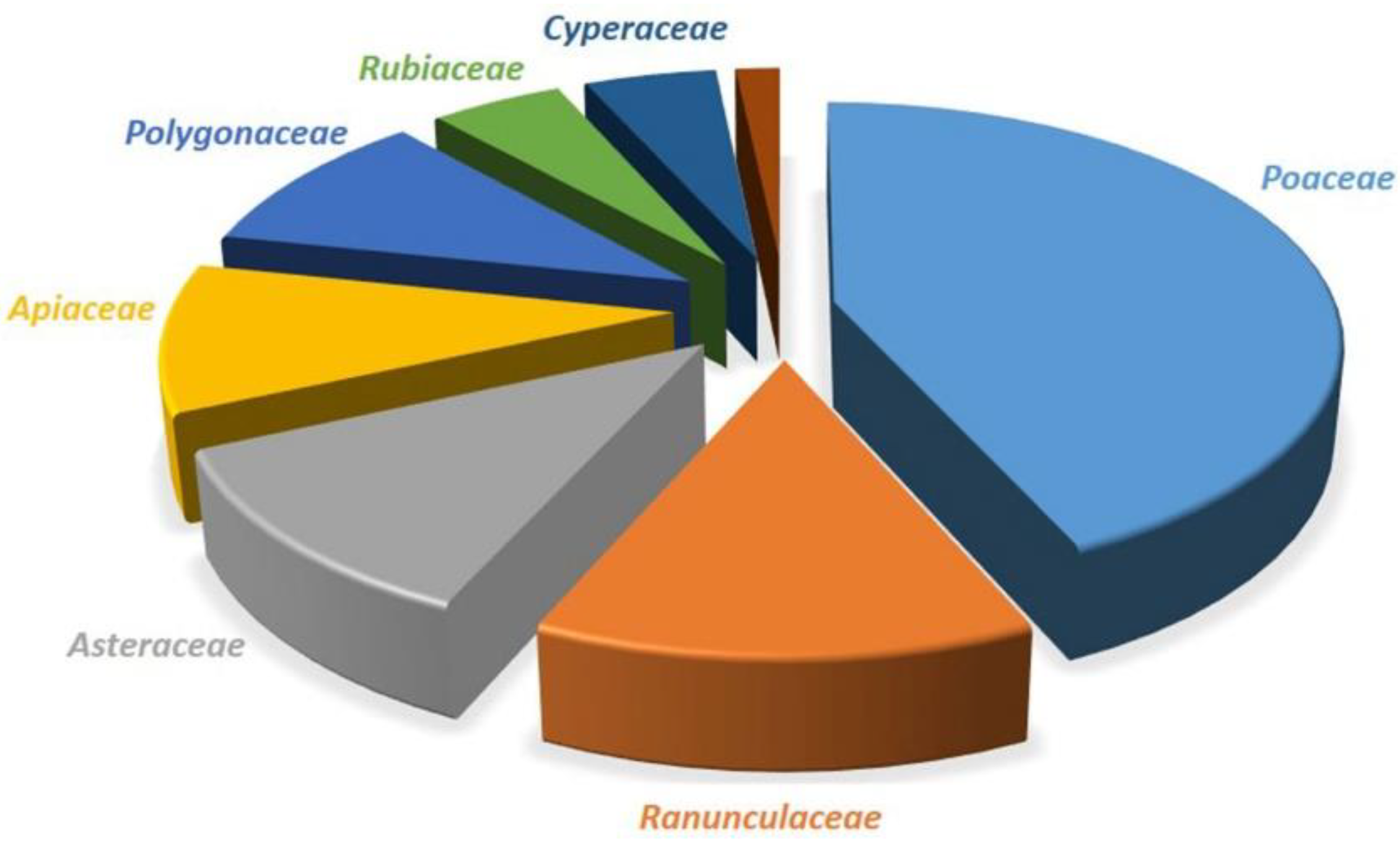
7. Threat to Biodiversity
8. Puccinia in the Present Century
9. Limitations of Current Knowledge
- Apart from the availability of 5000 species of Puccinia, only a few species are known to have DNA sequence data. The difficulty in culturing rust fungi is one of the possible reasons behind the reduced availability of molecular data. Further, DNA isolation directly from rust fungi present on a natural host and further processing are not easy tasks. In addition, the available sequences are not up to the mark. While others are too long, some sequences are too short, and some are incomplete. While some sequences are up to 300 nucleotides long, others are up to 800 nucleotides long, reflecting the intricacy of their taxonomic evaluation. Incomplete sequences in the Puccinia sequence dataset can result in two complexes in a single genus. Therefore, to investigate these complicated clades, entire gene sequences are required. The phylogenetic studies revealed the polyphyletic nature of the species of Puccinia, which require more DNA-based analyses for a better understanding of their taxonomic placement. The nonavailability of molecular data for all collections of Puccinia all over the globe is another limitation that highlights the requirement for fresh collections of Puccinia species and their molecular characterizations to generate molecular data so that their phylogenetic relationships can be explained more precisely. Although country-wide databases of rust fungi are available on various online platforms, the lack of a universal platform exclusively for global rust fungi is also a major limitation in the research on rust fungi, including Puccinia.
- When we talk about the day-by-day decreased interest of budding scientists/researchers in the field of the basic taxonomy of fungi, the reasons behind this are complex, such as insufficient funds, expensive outsourced mycological services, and, overall, the difficulty in publishing taxonomies in high-impact journals without modern techniques. These issues are leading to the decreased interest of mycologists in fungal taxonomy, which is ultimately decreasing the number of fungal taxonomists [32].
- Publication in high-impact journals has now become a criterion to assess the quality of research and the performance of researchers/scientists/academicians, or to appraise whether they should be promoted. However, taxonomy based on DNA-based molecular techniques has now become a minimum criterion to process any submitted manuscript, even for initial review. Luckily, few journals are still focusing on the novelty of the research, and most are now considering fungal manuscripts that are purely based on morpho-taxonomy.
- Despite being less expensive to support than applied research, basic fungus research is no longer prioritized for funding. This scenario is common in developing countries. The fundamental inventorying and identification of fungi is not everyone’s cup of tea, similar to obtaining funding for applied research (ideally in biotechnology). Due to a lack of sufficient funding, laboratories working on taxonomic studies of fungi continue to lack current equipment (e.g., that which is used in DNA isolation, amplification (PCR), and sequencing), and they are gradually turning their attention to the practical elements of the field. Additionally, not every mycologist can afford the service fees for the molecular techniques offered by many agencies/institutions of national and worldwide reputation, and particularly researchers working on a self-finance basis.
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cummins, G.B.; Hiratsuka, Y. Illustrated Genera of Rust Fungi, 3rd ed.; American Phytopathological Society (APS Press): St Paul, MN, USA, 2003. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Aime, M.C.; McTaggart, A.R.; Mondo, S.J.; Duplessis, S. Phylogenetics and phylogenomics of rust fungi. Adv. Genet. 2017, 100, 267–307. [Google Scholar]
- Aime, M.C.; Bell, C.D.; Wilson, A.W. Deconstructing the evolutionary complexity between rust fungi (Pucciniales) and their plant hosts. Stud. Mycol. 2018, 89, 143–152. [Google Scholar] [CrossRef]
- Aime, M.C.; Castlebury, L.A.; Abbasi, M.; Begerow, D.; Berndt, R.; Kirschner, R.; Marvanová, L.; Ono, Y.; Padamsee, M.; Scholler, M.; et al. Competing sexual and asexual generic names in Pucciniomycotina and Ustilaginomycotina (Basidiomycota) and recommendations for use. IMA Fungus 2018, 9, 75–89. [Google Scholar] [CrossRef]
- Aime, M.C.; McTaggart, A.R. A higher-rank classification for rust fungi, with notes on genera. Fungal Syst. Evol. 2021, 7, 21–47. [Google Scholar] [CrossRef]
- Gautam, A.K.; Verma, R.K.; Avasthi, S.; Sushma; Bohra, Y.; Devadatha, B.; Niranjan, M.; Suwannarach, N. Current insight into traditional and modern methods in fungal diversity estimates. J. Fungi 2022, 8, 226. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Dai, D.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.; Aptroot, A.; et al. Outline of fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Ordonez, M.E.; Groth, J.V. The Rust Fungi. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2009. [Google Scholar]
- Helfer, S. Rust fungi and global change. New Phytol. 2014, 201, 770–780. [Google Scholar] [CrossRef]
- Line, R.F.; Griffith, C.S. Research on the Epidemiology of Stem Rust of Wheat During the Cold War. In Stem Rust of Wheat. From Ancient Enemy to Modern Foe; Peterson, P.D., Ed.; American Phytopathological Society: St Paul, MN, USA, 2001; pp. 83–118. [Google Scholar]
- Hennen, J.F.; Buriticá, P. A brief summary of modern rust taxonomic and evolutionary theory. Rep. Tottori Mycol. Inst. 1980, 18, 243–256. [Google Scholar]
- Laundon, G.F. A new name for a New Zealand Phragmidium. Trans. Br. Mycol. Soc. 1976, 67, 177. [Google Scholar] [CrossRef]
- Holm, L. Etudes uredinologiques. l. Sur les ecidies de Oenotheracees. Sven. Bot. Tidskr. 1963, 57, 129–144. [Google Scholar]
- Laundon, G.F. Deliniation of aecial from uredial states. Trans. Br. Mycol. Soc. 1972, 5S, 344–346. [Google Scholar] [CrossRef]
- Arthur, J.C. Rusts on Compositae from Mexico. Bot. Gaz. 1905, 40, 196–208. [Google Scholar] [CrossRef]
- Arthur, J.C. The grass rusts of South America; based on the Holway collections. Proc. Am. Philos. Soc. 1925, 64, 131–223. [Google Scholar]
- Arthur, J.C. The Plant Rusts (Urediniales); Wiley: New York, NY, USA, 1929. [Google Scholar]
- Cummins, G.B. Illustrated Genera of Rust Fungi; Burgess: Minneapolis, Minnesota, 1959. [Google Scholar]
- Katsuya, K.; Green, G.J. Reproductive potentials of races 15B and 56 of wheat stem rust. Can. J. Bot. 1967, 45, 1077–1091. [Google Scholar] [CrossRef]
- Mont, R.M. Studies of Nonspecific Resistance to Stem Rust in Spring Wheat. Master’s Thesis, University of Minnesota, St. Paul, MN, USA, 1970; p. 61. [Google Scholar]
- Aime, M.C. Toward resolving family-level relationships in rust fungi (Uredinales). Mycoscience 2006, 47, 112–122. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Magurno, F. Outline of fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the raxml web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop 2010, New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Rambaut, A. FigTree version 1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 26 September 2022).
- Gautam, A.K.; Avasthi, S.; Verma, R.K.; Sushma; Niranjan, M.; Devadatha, B.; Jayawardena, R.S.; Suwannarach, N.; Karunarathna, S.C. A Global overview of diversity and phylogeny of the rust genus Uromyces. J. Fungi 2022, 8, 633. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S.; Verma, R.K.; Devadatha, B.; Jayawardena, R.S.; Sushma, R.K. Indian Pucciniales: Taxonomic outline with important descriptive notes. Mycosphere 2021, 12, 89–162. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S.; Verma, R.K.; Devadatha, B.; Sushma, R.K.; Bhadauria, R.; Prasher, I.B. Current status of research on rust fungi (Pucciniales) in India. Asian J. Mycol. 2021, 4, 40–80. [Google Scholar]
- Ji, J.; Li, Z.; Li, Y.; Kakishima, M. Phylogenetic approach for identification and life cycles of Puccinia (Pucciniaceae) species on Carex (Cyperaceae) from northeastern China. Phytotaxa 2022, 542, 221–255. [Google Scholar] [CrossRef]
- Getaneh, G.; Endale, H.; Teklu, N. Detection of barberry plants (Berberis holstii) as an alternate host of stem rust (Puccinia graminis) of wheat in Ethiopia. Pest Manag. J. Ethiop. 2016, 19, 17–26. [Google Scholar]
- Eshete, B.B. Status and challenges of wheat stem rust (Puccinia graminis f. sp. tritici) and threats of new races in Ethiopia. Int. J. Forest Horticul. 2018, 4, 22–31. [Google Scholar]
- Azmeraw, Y.; Admassu, B.; Abeyo, B.G.; Bacha, N. Virulence spectrum of Puccinia graminis f. sp. tritici in Northwest Ethiopia. Ethiop. J. Agric. Sci. 2020, 30, 87–97. [Google Scholar]
- Singh, R.P.; Hodson, D.P.; Jin, Y. Current status, likely migration and strategies to mitigate the threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. Perspect. Agricul. Vet. Sc. Nutrit. Nat. Res. 2006, 1, 1–13. [Google Scholar]
- Khanfri, S.; Boulif, M.; Lahlali, R. Yellow rust (Puccinia striiformis): A serious threat to wheat production worldwide. Not. Sci. Biol. 2018, 10, 410–423. [Google Scholar] [CrossRef]
- Li, F.; Upadhyaya, N.M.; Sperschneider, J.; Matny, O.; Nguyen-Phuc, H.; Mago, R.; Raley, C.; Miller, M.E.; Silverstein, K.A.T.; Henningsen, E.; et al. Emergence of the Ug99 lineage of the wheat stem rust pathogen through somatic hybridisation. Nat. Commun. 2019, 10, 5068. [Google Scholar] [CrossRef]
- Shahin, A.A.; Youssif, W.; EL-Naggar, D. Detection of Ug99 (TTKSK) of wheat stem rust fungus and new virulence races of Puccinia graminis f. sp. tritici in Egypt. Egypt. J. Phytopathol. 2020, 48, 14–28. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Sørensen, C.K.; Walter, S.; Justesen, A.F. Diversity of Puccinia striiformis on cereals and grasses. Annu. Rev. Phytopathol. 2011, 49, 197–217. [Google Scholar] [CrossRef]
- Anikster, A.Y.; Eilam, T.; Manisterski, J.; Leonard, K.J. Self-fertility and other distinguishing characteristics of a new morphotype of Puccinia coronata pathogenic on smooth brome grass. Mycologia 2003, 95, 87–97. [Google Scholar] [CrossRef]
- Carson, M.L. Virulence in oat crown rust (Puccinia coronata f. sp. avenae) in the United States from 2006 through 2009. Plant Dis. 2011, 95, 1528–1534. [Google Scholar] [CrossRef]
- Nazareno, E.S.; Li, F.; Smith, M.J.; Park, R.F.; Kianian, S.F.; Figueroa, M. Puccinia coronata f. sp. avenae: A threat to global oat production. Mol. Plant Pathol. 2018, 19, 1047–1060. [Google Scholar]
- Glen, M.; Alfenas, A.C.; Zauza, E.A.V.; Wingfield, M.J.; Mohammed, C. Puccinia psidii: A threat to the Australian environment and economy—A review. Australas. Plant Pathol. 2007, 36, 1–16. [Google Scholar] [CrossRef]
- Graça, R.N.; Ross-Davis, A.L.; Klopfenstein, N.B.; Kim, M.-S.; Peever, T.L.; Cannon, P.G.; Aun, C.P.; Mizubuti, E.S.G.; Alfenas, A.C. Rust disease of eucalypts, caused by Puccinia psidii, did not originate via host jump from guava in Brazil. Mol. Ecol. 2013, 22, 6033–6047. [Google Scholar] [CrossRef]
- Makinson, R.O.; Conn, B.J. Puccinia psidii (Pucciniaceae—Eucalyptus rust, guava rust, myrtle rust)—A threat to biodiversity in the Indo-Pacific region. Gard. Bull. Singap. 2014, 66, 173–188. [Google Scholar]
- Tommerup, I.C.; Alfenas, A.C.; Old, K.M.; Ridley, G.; Dick, M.A. Guava rust in Brazil—A threat to eucalyptus and other Myrtaceae. N. Z. J. For. Sci. 2003, 33, 420–428. [Google Scholar]
- Tobias, P.A.; Guest, D.I.; Külheim, C.; Hsieh, J.F.; Park, R.F. A curious case of resistance to a new encounter pathogen: Myrtle rust in Australia. Mol. Plant Pathol. 2016, 17, 783–788. [Google Scholar] [CrossRef]
- Beenken, L. Austropuccinia: A new genus name for the myrtle rust Puccinia psidii placed within the redefined family Sphaerophragmiaceae (Pucciniales). Phytotaxa 2017, 297, 53–61. [Google Scholar] [CrossRef]
- Henderson, D.M. A Checklist of Rust Fungi of British Isles; Royal Gerden Kew, Richmond Surrey; British Mycological Society: London, UK, 2000. [Google Scholar]
- Kamil, D.; Sharma, R.K.; Masheswari, C.U.; Prameela, D.T.; Jain, R.K. HCIO—Herbarium Cryptogamae Indiae Orientalis, Check List of Puccinia Species; Indian Agricultural Research Institute: New Delhi, India, 2013; p. 536. [Google Scholar]
- Maharachchikumbura, S.N.N.; Al-Sadi, A.M.; Al-Kharousi, M.; Al-Saady, N.A.; Hyde, K.D. A checklist of fungi in Oman. Phytotaxa 2016, 273, 219–261. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S. First checklist of rust fungi in the genus Puccinia from Himachal Pradesh, India. Plant Pathol. Quar. J. Fungal Biolog. 2016, 6, 106–120. [Google Scholar] [CrossRef]
- Talhinhas, P.; Carvalho, R.; Figueira, R.; Ramos, A.P. An annotated checklist of rust fungi (Pucciniales) occurring in Portugal. Sydowia 2019, 71, 65–84. [Google Scholar]
- Gautam, A.K.; Avasthi, S. A checklist of rust fungi from Himachal Pradesh, India. J. Threat. Taxa 2019, 11, 14845–14861. [Google Scholar] [CrossRef]
- Berndt, R.; Brodtbeck, T. Checklist and Host Index of the Rust Fungi (Uredinales) of Switzerland; ETH Zurich: Zurich, Switzerland, 2020; p. 247. [Google Scholar]
- Abbasi, M. A checklist of rust fungi (Pucciniales) in Iran. J. Crop Prot. 2021, 10, 175–259. [Google Scholar]
- Brar, G.S. Population structure of Puccinia striiformis f. sp. tritici, the cause of wheat stripe rust, in western Canada. Master’s Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2015. [Google Scholar]
- Li, T.Y.; Ma, Y.C.; Wu, X.X.; Chen, S.; Xu, X.F.; Wang, H.; Cao, Y.Y.; Xuan, Y.H. Race and virulence characterization of Puccinia graminis f. sp. tritici in China. PLoS ONE 2018, 13, 0197579. [Google Scholar]
- Sriram, S.; Chandran, N.K.; Kumar, R.; Reddy, K.M. First report of Puccinia horiana causing white rust of chrysanthemum in India. New Dis. Rep. 2015, 32, 8. [Google Scholar] [CrossRef]
- Göre, M.E. Geranium rust disease caused by Puccinia pelargonii-zonalis: First report in Turkey. Plant Pathol. 2008, 57, 786. [Google Scholar]
- Ono, Y.; Kakishima, M. Puccinia pulchella: A new Viola–Carex rust from Japan. Can. J. Bot. 2011, 59, 1543–1546. [Google Scholar] [CrossRef]
- Negash, T.; Shif, H. Garlic rust (Puccinia allii): Effect and management options—A review. Social Sci. Rev. 2018, 4, 32–37. [Google Scholar]
- Uppalapati, S.R.; Serba, D.D.; Ishiga, Y.; Szabo, L.J.; Mittal, S.; Bhandari, H.S.; Bouton, J.H.; Mysore, K.S.; Saha, M.C. Characterization of the rust fungus, Puccinia emaculata, and evaluation of genetic variability for rust resistance in switchgrass populations. Bioenerg. Res. 2013, 6, 458–468. [Google Scholar] [CrossRef]
- Muè Ller–Schaèrer, H.; Rieger, S. Epidemic spread of the rust fungus Puccinia lagenophorae and its impact on the competitive ability of senecio vulgaris in celeriac during early development. Biocontrol Sci. Technol. 1998, 8, 59–72. [Google Scholar] [CrossRef]
- Mondal, S.; Badigannavar, A.M. Peanut rust (Puccinia arachidis Speg.) disease: Its background and recent accomplishments towards disease resistance breeding. Protoplasma 2015, 252, 1409–1420. [Google Scholar] [CrossRef]
- Verma, R.K.; Gautam, A.K.; Singh, A.; Avasthi, S.; Prasher, I.B.; Nautiyal, M.C.; Singh, H. New record of rust disease caused by Puccinia oxalidis on Oxalis latifolia from India. MycoAsia 2020, 2020, 1. [Google Scholar] [CrossRef]
- Maciel, J.C.; Costa, M.R.; Ferreira, E.A.; Oliveira, I.T.; Alencar, B.T.B.; Zanuncio, J.C.; Santos, J.B. Puccinia oxalidis Dietel & Ellis (1895): First report controlling Oxalis latifolia kunth (Oxalidaceae) in systems of direct planting. Braz. J. Biol. 2021, 84, 249087. [Google Scholar]
- Index Fungorum. Available online: https://www.indexfungorum.org/names/Names.asp (accessed on 20 April 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Taxa | GenBank Accession Number | |
|---|---|---|
| ITS | LSU | |
| Helicobasidium purpureum CBS 163.24 | AY292433 | AY254181 |
| Puccinia adenocauli HMJAU 8630 | MK785267 | MK785292 |
| Puccinia aizazii BA34C | – | KY386659 |
| Puccinia aizazii ISL 46866 | NR158929 | – |
| Puccinia albispora TSHR 11044 | MT560796 | MT560813 |
| Puccinia amari BPI 064435 | KX190836 | KX190914 |
| Puccinia ampliaticoronata HMJAU 8734 | – | MW404927 |
| Puccinia ampliaticoronata HMJAU 8735 | MW404766 | – |
| Puccinia aridariae RSA 155 | – | DQ917725 |
| Puccinia baccharidis BPI 910214 | KY764097 | – |
| Puccinia calystegiae-soldanellae HamaITSA2 | AB125956 | – |
| Puccinia canaliculata var. tenuis TA 430 | – | OL437033 |
| Puccinia caricis-artemisiae HMJAU 8874 | MW447821 | MW414430 |
| Puccinia caricis-atractylodes HMJAU 8888 | MW447811 | MW414419 |
| Puccinia caricis BPI 871515 | – | DQ354514 |
| Puccinia caricis-hebeiensis HMJAU 8895 | MW447818 | MW414427 |
| Puccinia caricis-jilinensis HMJAU 8842 | MW447898 | MW414367 |
| Puccinia caricis-lactucae HMJAU 8902 | MW447781 | MW414384 |
| Puccinia caricis NYBG 69670 | MK518859 | MK518517 |
| Puccinia caricis-pediformis HMJAU 8836 | MW447855 | MW414323 |
| Puccinia caricis-rafaensis HMJAU 8798 | MW447892 | MW414360 |
| Puccinia caricis-ribicola HMJAU 8871 | MW447805 | MW414413 |
| Puccinia caricis-tenuiformis HMJAU 8851 | MW447858 | MW414326 |
| Puccinia caricis-violae HMJAU 8664 | MW447798 | MW414406 |
| Puccinia caulophylli HMJAU 8633 | MK785263 | MK785288 |
| Puccinia chardoniensis R177 | – | EU851149 |
| Puccinia chunjiei DAOM 240982 | NR111548 | - |
| Puccinia coleataeniae BPI 006819 | KX190843 | KX190919 |
| Puccinia convolvuli BPI 871465 | – | DQ354512 |
| Puccinia coronata BPI 844300 | – | DQ354526 |
| Puccinia coronata KUSF 30637 | MT393874 | MT393876 |
| Puccinia coronati-agrostidis HMJAU 8740 | MW404761 | MW404921 |
| Puccinia coronati-agrostidis PURN 114 | NR111528 | – |
| Puccinia coronati-brevispora PURN 652 | NR111526 | – |
| Puccinia coronati-calamagrostidis DAOM 204923 | HM131305 | – |
| Puccinia coronati-japonica PURF 16131 | NR111527 | – |
| Puccinia coronati-longispora PRC 194 | HM131233 | – |
| Puccinia crotonopsidis BPI 006810 | KX190844 | KX190920 |
| Puccinia cumminsii DAOM 114236 | KX190845 | - |
| Puccinia dampierae BRIP 57724 | – | KF690688 |
| Puccinia dianellae BRIP 57433 | KM249859 | – |
| Puccinia digitaticoronata HMJAU 8773 | MW404702 | MW404862 |
| Puccinia dimidipes BPI 195034 | MH144395 | - |
| Puccinia eleganticoronata HMJAU 8286 | MW404721 | MW404881 |
| Puccinia elymi-albispora HMJAU 8677 | MW404820 | MW404980 |
| Puccinia emaculata BPI 851570 | NR148108 | – |
| Puccinia gansensis DAOM 240065 | HM057115 | – |
| Puccinia geitonoplesii BRIP 55679 | KM249860 | – |
| Puccinia gilgiana BRIP 57719 | – | KF690691 |
| Puccinia graminicola PURN 10143 | MH707050 | MH704521 |
| Puccinia graminis ECS | – | AF522177 |
| Puccinia grevilleae BRIP 55600 | – | KX999878 |
| Puccinia haemodori BRIP 56965 | – | KF690692 |
| Puccinia hemerocallidis BPI 843967 | DQ354519 | – |
| Puccinia hordei AFTOLID 1402 | – | DQ354527 |
| Puccinia klugkistiana KUSF 31285 | MW740211 | MW740212 |
| Puccinia knersvlaktensis RSA 176 | – | DQ917726 |
| Puccinia komarovii IMI 502296 | KC466553 | – |
| Puccinia lagenophorae BRIP 57563 | – | KF690696 |
| Puccinia latimamma ZPR 1398 | MK518986 | MK518685 |
| Puccinia lycii RBerndt 294 | MH144384 | – |
| Puccinia menthae BPI 871110 | DQ354513 | – |
| Puccinia merrilliana BRIP 56913 | – | KX999885 |
| Puccinia microsora DAOM 106309 | MW009501 | – |
| Puccinia millegranae BPI 086067 | – | NG059683 |
| Puccinia mysuruensis HSZ 2119 | KC847089 | – |
| Puccinia novopanici BPI 747673 | NR148109 | – |
| Puccinia novopanici BPI 893095 | KX190888 | KX190947 |
| Puccinia oncospora HMJAU 8691 | MW404822 | MW404982 |
| Puccinia otzeniani RSA 164 | – | DQ917742 |
| Puccinia panici urvilleani JE 2017d | – | BPI841053 |
| Puccinia pascua PUR 11682 | MH707035 | MH704507 |
| Puccinia peradeniyae BPI 089014 | KX190906 | – |
| Puccinia peradeniyae BPI 871072 | – | GU057996 |
| Puccinia pileiformis HMJAU8701 | MW404792 | MW404952 |
| Puccinia poarum AFTOLID 1027 | – | DQ831028 |
| Puccinia polysora BPI 863756 | – | GU058024 |
| Puccinia protuberanticoronata HMJAU 8718 | MW404752 | MW404912 |
| Puccinia pseudodigitata BPI 085603 | KF661261 | – |
| Puccinia pseudomesnieriana BPI 085621 | KF661263 | – |
| Puccinia pseudostriiformis IRAN 11500 | AY956560 | – |
| Puccinia pseudostriiformis PSP91WA | KM507443 | – |
| Puccinia pseudostriiformis PUR 59844 | MT965634 | – |
| Puccinia psidii BRIP 57991 | – | KF318443 |
| Puccinia pulverulenta NZFRI 29019 | KM065015 | – |
| Puccinia rapipes Prap_4 | MK874620 | – |
| Puccinia rhagodiae BRIP 60078 | – | KX999890 |
| Puccinia saccardoi BRIP 57725 | – | KF690701 |
| Puccinia salihae BPI 881123 | – | HQ412645 |
| Puccinia setariae BPI 188745 | NR148110 | – |
| Puccinia smilacis BPI 871784 | DQ354533 | – |
| Puccinia sporoboli var. robusta BPI 871549 | – | GU058003 |
| Puccinia striiformoides BPI 199096 | HM057137 | – |
| Puccinia suaveolens KRM 0005945 | ON063373 | – |
| Puccinia taeniatheri IRAN 11491 | AY956557 | – |
| Puccinia tuberosa HMJAU 8949 | OK489429 | OK489421 |
| Puccinia ursiniae BRIP 57993 | – | – |
| Puccinia violae BPI 842321 | DQ354509 | – |
| Puccinia wiehei BPI 111530 | NR148111 | – |
| Puccinia windhoekensis NA 152 | – | DQ917710 |
| Puccinia xanthii BRIP 48819 | EU659694 | – |
| Puccinia xanthosiae BRIP 57729 | – | KF690706 |
| Puccinia xinyuanensis HMUT 2549 | NR173762 | NG079639 |
| Uromyces scaevolae BRIP 60096 | – | KJ622214 |
| Year Range | Number of Publications | |
|---|---|---|
| Decadal Trend | Centurial Trend | |
| 1794–1800 | 02 | 02 |
| 1801–1810 | 03 | 277 |
| 1811–1820 | 06 | |
| 1821–1830 | 09 | |
| 1831–1840 | 06 | |
| 1841–1850 | 03 | |
| 1851–1860 | 09 | |
| 1861–1870 | 09 | |
| 1871–1880 | 61 | |
| 1881–1890 | 50 | |
| 1891–1900 | 121 | |
| 1901–1910 | 144 | 627 |
| 1911–1920 | 85 | |
| 1921–1930 | 51 | |
| 1931–1940 | 71 | |
| 1941–1950 | 58 | |
| 1951–1960 | 78 | |
| 1961–1970 | 28 | |
| 1971–1980 | 44 | |
| 1981–1990 | 39 | |
| 1991–2000 | 29 | |
| 2001–2010 | 42 | 82 |
| 2011–2022 | 40 | |
| Rust Disease | Causal Organism | Host Plant | Reference |
|---|---|---|---|
| Rust disease of eucalyptus | Puccinia psidii | Eucalyptus spp. | [46] |
| Stripe rust of wheat | Puccinia striiformis | Triticum sp. | [38,59] |
| Stem rust of wheat | Puccinia graminis | Triticum sp. | [36,60] |
| Guava rust | Puccinia psidii | Psidium guajava | [48] |
| Crown rust of cultivated and wild oats | Puccinia coronata f.sp. avenae | Avena sativa | [44] |
| White rust of chrysanthemum | Puccinia horiana | Chrysanthemum sp. | [61] |
| Geranium rust | Puccinia pelargonic-zonalis | Pelargonium × hortorum | [62] |
| Viola rust | Puccinia viola and P. pulchella | Viola sp. | [63] |
| Garlic rust | Puccinia allii | Allium sativum | [64] |
| Switchgrass rust | Pucciniaemaculata | Panicumvirgatum | [65] |
| Senecio rust | Puccinia lagenophorae | Senecio vulgaris | [66] |
| Peanut rust | Puccinia arachidis | Arachis spp. | [67] |
| Oxalis rust | Puccinia oxalidis | Oxalis latifolia | [68,69] |
| Buckwheat rust | Puccinia fagopyri | Fagopyrum esculentum | [54] |
| Mint rust | Puccinia menthae | Mentha longifolia | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avasthi, S.; Gautam, A.K.; Niranjan, M.; Verma, R.K.; Karunarathna, S.C.; Kumar, A.; Suwannarach, N. Insights into Diversity, Distribution, and Systematics of Rust Genus Puccinia. J. Fungi 2023, 9, 639. https://doi.org/10.3390/jof9060639
Avasthi S, Gautam AK, Niranjan M, Verma RK, Karunarathna SC, Kumar A, Suwannarach N. Insights into Diversity, Distribution, and Systematics of Rust Genus Puccinia. Journal of Fungi. 2023; 9(6):639. https://doi.org/10.3390/jof9060639
Chicago/Turabian StyleAvasthi, Shubhi, Ajay Kumar Gautam, Mekala Niranjan, Rajnish Kumar Verma, Samantha C. Karunarathna, Ashwani Kumar, and Nakarin Suwannarach. 2023. "Insights into Diversity, Distribution, and Systematics of Rust Genus Puccinia" Journal of Fungi 9, no. 6: 639. https://doi.org/10.3390/jof9060639