
Uncovering Phytotoxic Compounds Produced by Colletotrichum spp. Involved in Legume Diseases Using an OSMAC–Metabolomics Approach

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Plant Hosts, and Crossing Inoculations
2.2. Culture Medium and Growth Conditions for Fungal Organic Extract Production
2.3. Reagents and Materials
2.4. Extraction of Secondary Metabolites from Different Culture Media
2.5. UHPLC-QTOF-HRMS Apparatus and Conditions
2.6. Metabolite Annotation and Identification
2.7. Bioassays
2.8. Bioassay Data Analysis
3. Results
3.1. Cross Inoculations
3.2. Targeted Analysis of Selected Phytotoxic Metabolites
3.3. Impact of Cultural Conditions on Secondary Metabolite Production by Colletotrichum Species
3.3.1. Colletotrichum truncatum C428 (from Lentil)
3.3.2. Colletotrichum truncatum C431 (from Soybean)
3.3.3. Colletotrichum trifolii C436 (from Red Clover)
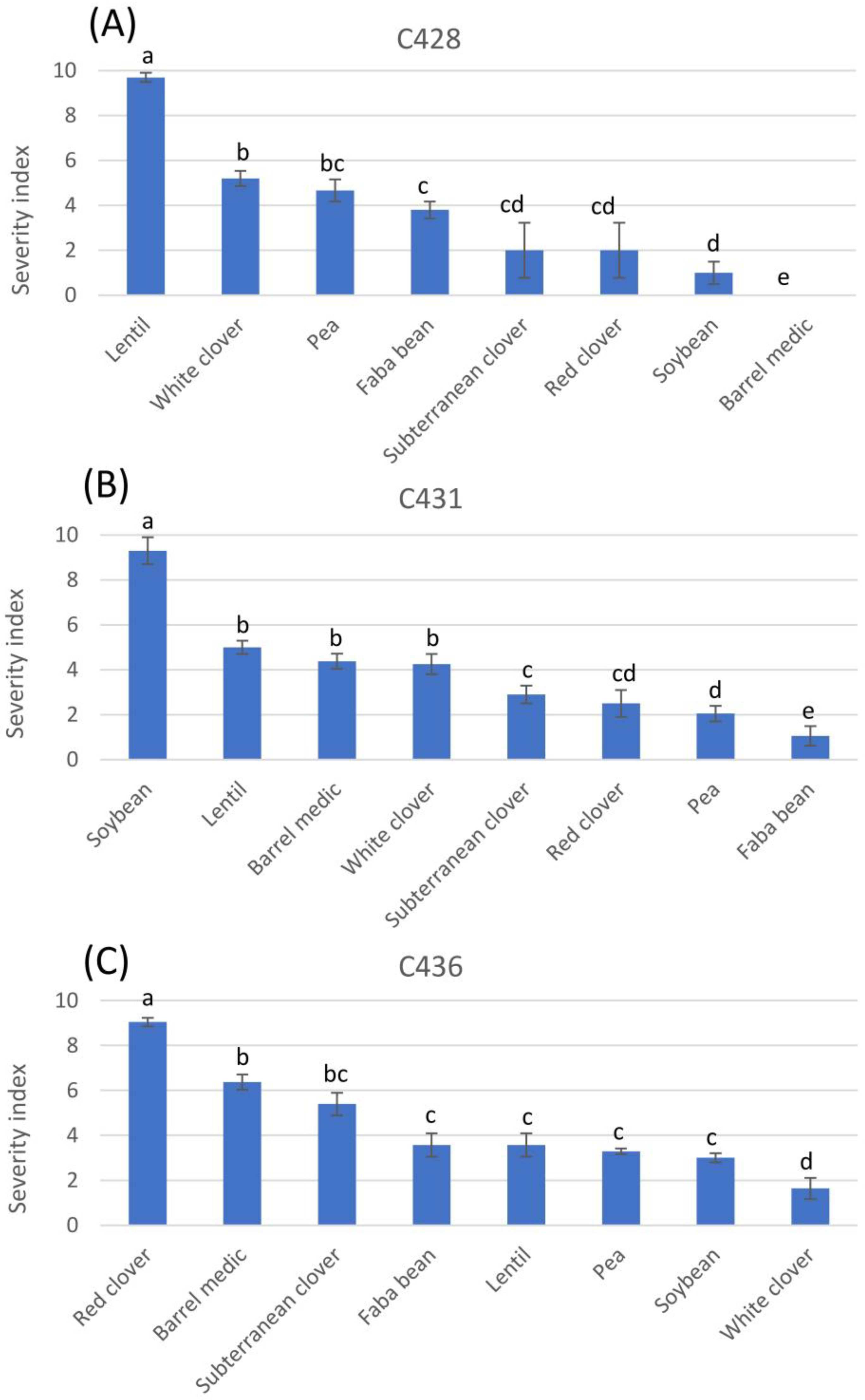
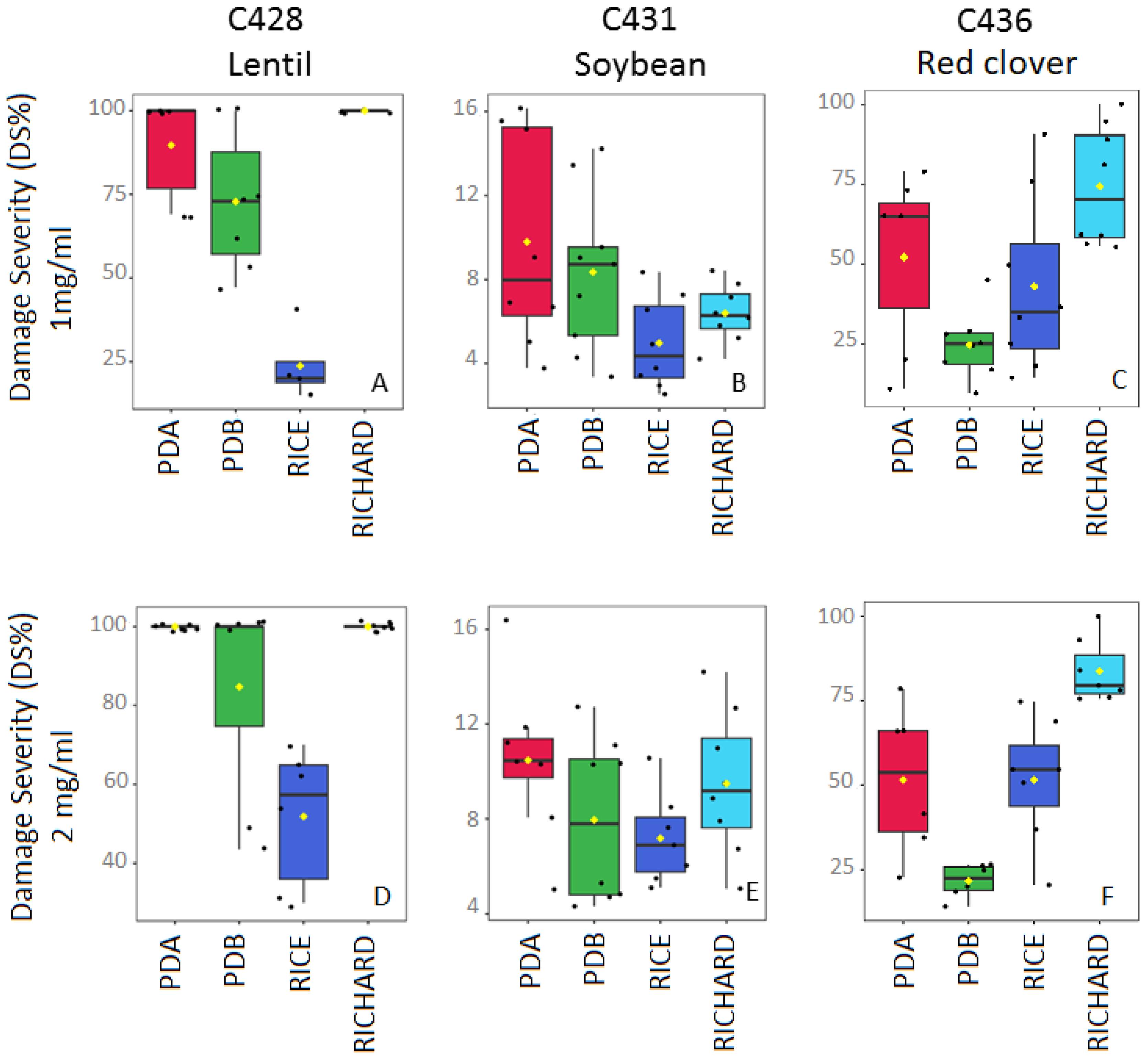
3.4. Effects of Culture Media on the Phytotoxicity of Organic Extracts
3.4.1. Colletotrichum truncatum C428 (from Lentil)
3.4.2. Colletotrichum truncatum C431 (from Soybean)
3.4.3. Colletotrichum trifolii C436 (from Red Clover)
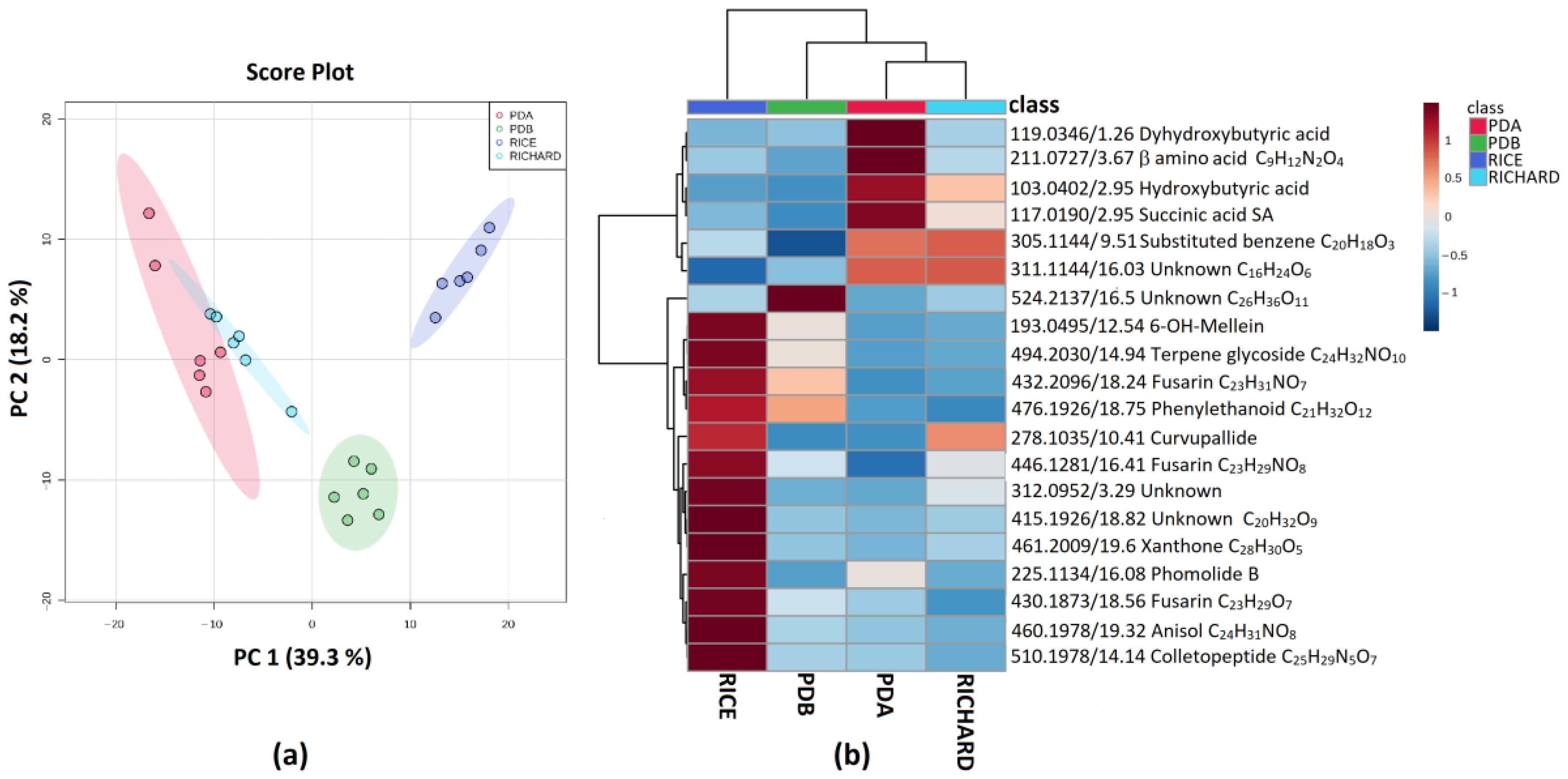
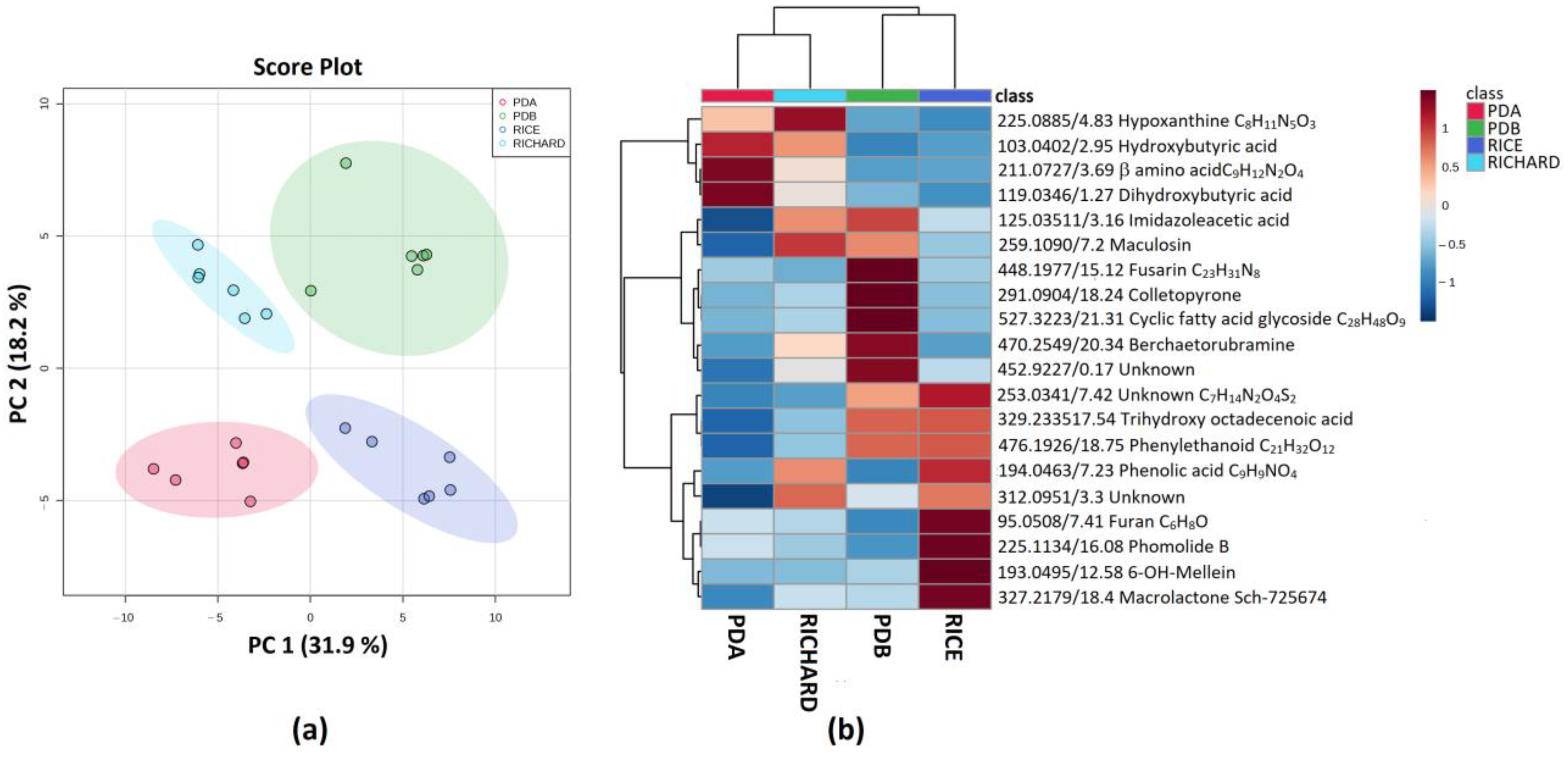
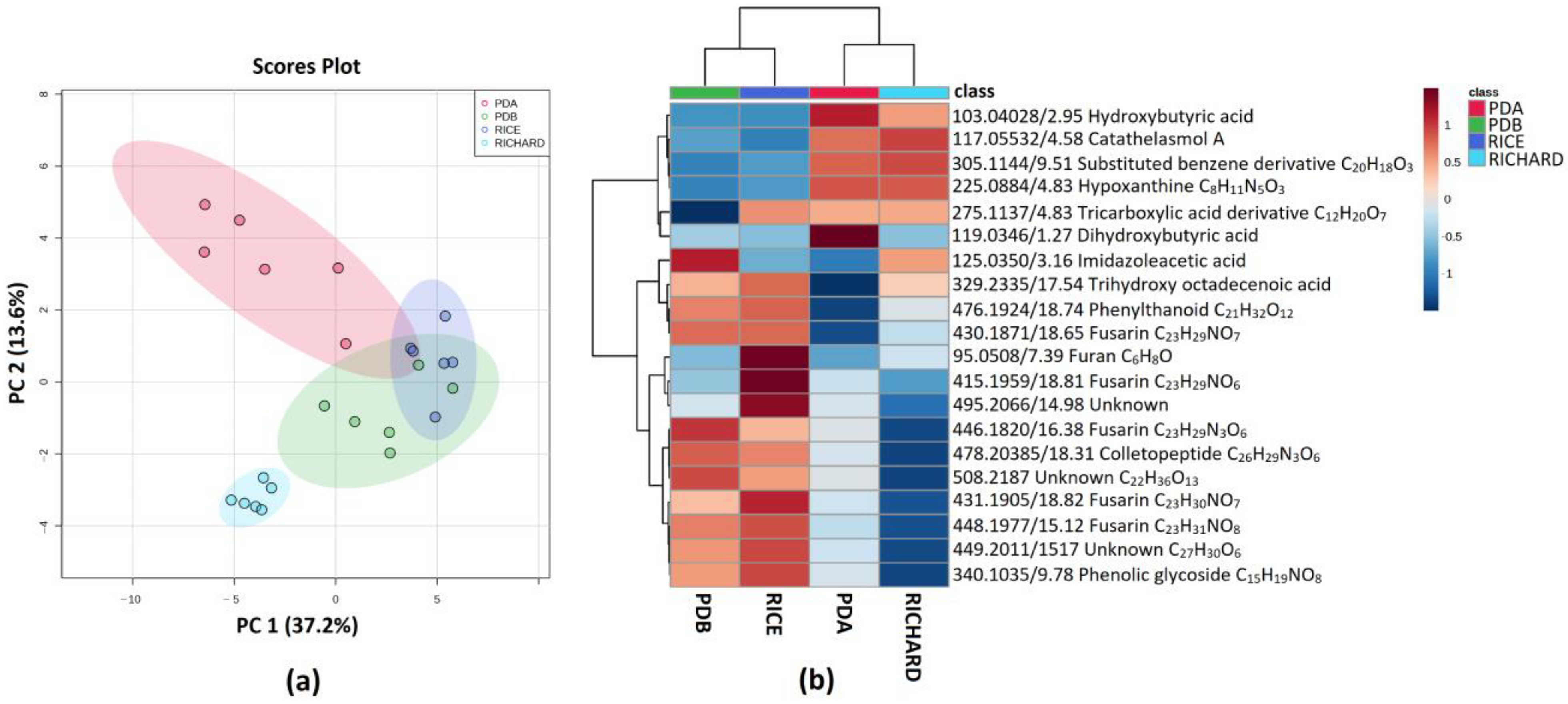
3.5. Secondary Metabolite Profiles as a Tool for Chemotaxonomy of Colletotrichum Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef]
- Cannon, P.; Damm, U.; Johnston, P.; Weir, B. Colletotrichum: Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef]
- García-Pajón, C.; Collado, I.G. Secondary metabolites isolated from Colletotrichum species. Nat. Prod. Rep. 2003, 20, 426–431. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Boufleur, T.R.; Ciampi-Guillardi, M.; Tikami, Í.; Rogério, F.; Thon, M.R.; Sukno, S.A.; Massola Junior, N.S.; Baroncelli, R. Soybean anthracnose caused by Colletotrichum species: Current status and future prospects. Mol. Plant Pathol. 2021, 22, 393–409. [Google Scholar]
- Begum, M.; Sariah, M.; Puteh, A.; Abidin, M.Z. Pathogenicity of Colletotrichum truncatum and its influence on soybean seed quality. Int. J. Agric. Biol. 2008, 10, 393–398. [Google Scholar]
- Tullu, A.; Buchwaldt, L.; Lulsdorf, M.; Banniza, S.; Barlow, B.; Slinkard, A.; Sarker, A.; Tar’an, B.; Warkentin, T.; Vandenberg, A. Sources of resistance to anthracnose (Colletotrichum truncatum) in wild Lens species. Genet. Resour. Crop Evol. 2006, 53, 111–119. [Google Scholar] [CrossRef]
- O’Connell, R.; Uronu, A.; Waksman, G.; Nash, C.; Keon, J.; Bailey, J. Hemibiotrophic infection of Pisum sativum by Colletotrichum truncatum. Plant Pathol. 1993, 42, 774–783. [Google Scholar] [CrossRef]
- Schubiger, F.; Streckeisen, P.; Boller, B. Resistance to southern anthracnose (Colletotrichum trifolii) in cultivars of red clover (Trifolium pratense). Czech J. Genet. Plant Breed. 2003, 39, 309. [Google Scholar]
- Mould, M.J.; Boland, G.; Robb, J. Ultrastructure of the Colletotrichum trifolii-Medicago sativa pathosystem. I. Pre-penetration events. Physiol. Mol. Plant Pathol. 1991, 38, 179–194. [Google Scholar] [CrossRef]
- Gossen, B.; Anderson, K.; Buchwaldt, L. Host specificity of Colletotrichum truncatum from lentil. Can. J. Plant Pathol. 2009, 31, 65–73. [Google Scholar] [CrossRef]
- Pradhan, A.; Ghosh, S.; Sahoo, D.; Jha, G. Fungal effectors, the double edge sword of phytopathogens. Curr. Genet. 2021, 67, 27–40. [Google Scholar]
- Evidente, A.; Cimmino, A.; Masi, M. Phytotoxins produced by pathogenic fungi of agrarian plants. Phytochem. Rev. 2019, 18, 843–870. [Google Scholar] [CrossRef]
- Möbius, N.; Hertweck, C. Fungal phytotoxins as mediators of virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef]
- Cox, R.J. Polyketides, proteins and genes in fungi: Programmed nano-machines begin to reveal their secrets. Org. Biomol. Chem. 2007, 5, 2010–2026. [Google Scholar] [CrossRef]
- Scharf, D.H.; Heinekamp, T.; Brakhage, A.A. Human and plant fungal pathogens: The role of secondary metabolites. PLoS Pathog. 2014, 10, e1003859. [Google Scholar] [CrossRef]
- Pusztahelyi, T.; Holb, I.J.; Pócsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Moraga, J.; Gomes, W.; Pinedo, C.; Cantoral, J.M.; Hanson, J.R.; Carbú, M.; Garrido, C.; Durán-Patrón, R.; Collado, I.G. The current status on secondary metabolites produced by plant pathogenic Colletotrichum species. Phytochem. Rev. 2019, 18, 215–239. [Google Scholar] [CrossRef]
- Stoessl, A.; Stothers, J. Colletruncoic acid methyl ester, a unique meroterpenoid from Colletotrichum truncatum. Z. Für Nat. C 1986, 41, 677–680. [Google Scholar] [CrossRef]
- Masi, M.; Castaldi, S.; Sautua, F.; Pescitelli, G.; Carmona, M.A.; Evidente, A. Truncatenolide, a bioactive disubstituted nonenolide produced by Colletotrichum truncatum, the causal agent of Aanthracnose of soybean in Argentina: Fungal antagonism and SAR Sstudies. J. Agr. Food Chem. 2022, 70, 9834–9844. [Google Scholar] [CrossRef]
- Agudo-Jurado, F.J.; Reveglia, P.; Rubiales, D.; Evidente, A.; Barilli, E. Status of phytotoxins isolated from necrotrophic fungi causing diseases on grain legumes. Int. J. Mol. Sci. 2023, 24, 5116. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Kim, W.; Peever, T.L.; Park, J.-J.; Park, C.-M.; Gang, D.R.; Xian, M.; Davidson, J.A.; Infantino, A.; Kaiser, W.J.; Chen, W. Use of metabolomics for the chemotaxonomy of legume-associated Ascochyta and allied genera. Sci. Rep. 2016, 6, 20192. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Farinella, V.F.; Kawafune, E.S.; Tangerina, M.M.; Domingos, H.V.; Costa-Lotufo, L.V.; Ferreira, M.J. OSMAC strategy integrated with molecular networking for accessing griseofulvin derivatives from endophytic fungi of Moquiniastrum polymorphum (Asteraceae). Molecules 2021, 26, 7316. [Google Scholar] [CrossRef]
- Li, G.; Jian, T.; Liu, X.; Lv, Q.; Zhang, G.; Ling, J. Application of metabolomics in fungal research. Molecules 2022, 27, 7365. [Google Scholar] [CrossRef]
- Ball, B.; Langille, M.; Geddes-McAlister, J. Fun (gi) omics: Advanced and diverse technologies to explore emerging fungal pathogens and define mechanisms of antifungal resistance. Mbio 2020, 11, e01020. [Google Scholar] [CrossRef]
- Khan, A.I.; Bhandari, R.R.; Pokhrel, A.; Yadav, R.N. A study on root exudation pattern and effect of plant growth promoting fungi during biotic and abiotic stress in pigeon pea. World J. Agric. Res. 2018, 6, 122–131. [Google Scholar]
- Masi, M.; Zonno, M.C.; Cimmino, A.; Reveglia, P.; Berestetskiy, A.; Boari, A.; Vurro, M.; Evidente, A. On the metabolites produced by Colletotrichum gloeosporioides a fungus proposed for the Ambrosia artemisiifolia biocontrol; spectroscopic data and absolute configuration assignment of colletochlorin A. Nat. Prod. Res. 2017, 32, 1537–1547. [Google Scholar] [CrossRef]
- Cimmino, A.; Mathieu, V.; Masi, M.; Baroncelli, R.; Boari, A.; Pescitelli, G.; Ferderin, M.N.; Lisy, R.; Evidente, M.; Tuzi, A.; et al. Higginsianins A and B, two diterpenoid α-pyrones produced by Colletotrichum higginsianum, with in vitro cytostatic activity. J. Nat. Prod. 2016, 79, 116–125. [Google Scholar] [CrossRef]
- Masi, M.; Cimmino, A.; Boari, A.; Tuzi, A.; Zonno, M.C.; Baroncelli, R.; Vurro, M.; Evidente, A. Colletochlorins E and F, new phytotoxic tetrasubstituted pyran-2-one and dihydrobenzofuran, isolated from Colletotrichum higginsianum with potential herbicidal activity. J. Agr. Food Chem. 2017, 65, 1124–1130. [Google Scholar] [CrossRef]
- Masi, M.; Reveglia, P.; Baaijens-Billones, R.; Górecki, M.; Pescitelli, G.; Savocchia, S.; Evidente, A. Phytotoxic metabolites from three Neofusicoccum species causal agents of Botryosphaeria dieback in Australia, luteopyroxin, neoanthraquinone, and Lluteoxepinone, a disubstituted furo-α-pyrone, a hexasubstituted anthraquinone, and a trisubstituted oxepi-2-one from Neofusicoccum luteum. J. Nat. Prod. 2020, 83, 453–460. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Berestetskiy, A.; Vurro, M.; Evidente, A. Chenopodolans A–C: Phytotoxic furopyrans produced by Phoma chenopodiicola, a fungal pathogen of Chenopodium album. Phytochemistry 2013, 96, 208–213. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Cimmino, A.; Evidente, A. Isolation of phytotoxic phenols and characterization of a new 5-hydroxymethyl-2-isopropoxyphenol from Dothiorella vidmadera, a causal agent of grapevine trunk disease. J. Agr. Food Chem. 2018, 66, 1760–1764. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Evidente, A. Spencertoxin and spencer acid, new phytotoxic derivatives of diacrylic acid and dipyridinbutan-1,4-diol produced by Spencermartinsia viticola, a causal agent of grapevine Botryosphaeria dieback in Australia. Arab. J. Chem. 2018, 13, 1803–1808. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Met. 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Ruttkies, C.; Neumann, S.; Posch, S. Improving MetFrag with statistical learning of fragment annotations. BMC Bioinform. 2019, 20, 376. [Google Scholar] [CrossRef]
- van Santen, J.A.; Poynton, E.F.; Iskakova, D.; McMann, E.; Alsup, T.A.; Clark, T.N.; Fergusson, C.H.; Fewer, D.P.; Hughes, A.H.; McCadden, C.A. The Natural Products Atlas 2.0: A database of microbially-derived natural products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef]
- Sorokina, M.; Merseburger, P.; Rajan, K.; Yirik, M.A.; Steinbeck, C. COCONUT online: Collection of open natural products database. J. Cheminform. 2021, 13, 2. [Google Scholar] [CrossRef]
- Rutz, A.; Sorokina, M.; Galgonek, J.; Mietchen, D.; Willighagen, E.; Gaudry, A.; Graham, J.G.; Stephan, R.; Page, R.; Vondrášek, J. The LOTUS initiative for open knowledge management in natural products research. Elife 2022, 11, e70780. [Google Scholar] [CrossRef]
- Alseekh, S.; Aharoni, A.; Brotman, Y.; Contrepois, K.; D’Auria, J.; Ewald, J.; Ewald, J.C.; Fraser, P.D.; Giavalisco, P.; Hall, R.D. Mass spectrometry-based metabolomics: A guide for annotation, quantification and best reporting practices. Nat. Met. 2021, 18, 747–756. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Andolfi, A.; Cimmino, A.; Villegas-Fernández, A.; Tuzi, A.; Santini, A.; Melck, D.; Rubiales, D.; Evidente, A. Lentisone, a new phytotoxic anthraquinone produced by Ascochyta lentis, the causal agent of Ascochyta blight in Lens culinaris. J. Agric. Food Chem. 2013, 61, 7301–7308. [Google Scholar] [CrossRef]
- Abraham, W.-R.; Meyer, H.; Abate, D. Curvupallides, a new class of alkaloids from the fungus Curvularia pallescens. Tetrahedron 1995, 51, 4947–4952. [Google Scholar] [CrossRef]
- Du, X.; Lu, C.; Li, Y.; Zheng, Z.; Su, W.; Shen, Y. Three new antimicrobial metabolites of Phomopsis sp. J. Antibiot. 2008, 61, 250–253. [Google Scholar] [CrossRef]
- Feng, L.; Wang, J.; Liu, S.; Zhang, X.-J.; Bi, Q.-R.; Hu, Y.-Y.; Wang, Z.; Tan, N.-H. Colletopeptides A-D, anti-inflammatory cyclic tridepsipeptides from the plant endophytic fungus Colletotrichum sp. S8. J. Nat. Prod. 2019, 82, 1434–1441. [Google Scholar]
- Kleigrewe, K.; Aydin, F.; Hogrefe, K.; Piecuch, P.; Bergander, K.; Würthwein, E.-U.; Humpf, H.-U. Structure elucidation of new fusarins revealing insights in the rearrangement mechanisms of the Fusarium mycotoxin fusarin C. J. Agric. Food Chem. 2012, 60, 5497–5505. [Google Scholar] [CrossRef]
- List, P.; Reith, H. Basische Pilzinhaltsstoffe, X. Imidazolderivate im Faltentintling, Coprinus atramentarius Bull. no. Jahresband 1960, 319, 17–21. [Google Scholar]
- Lee, H.-B.; Choi, Y.-C.; Kim, S.-U. Isolation and identification of maculosins from Streptomyces rochei 87051-3. Appl. Biol. Chem. 1994, 37, 339–342. [Google Scholar]
- Park, S.H.; Strobel, G.A. Cellular protein receptors of maculosin, a host specific phytotoxin of spotted knapweed (Centaurea maculosa L.). Biochim. Biophys. Acta Gen. Subj. 1994, 1199, 13–19. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Girtsman, T.; Mou, T.; Antczak, C.; Djaballah, H. Azaphilones from an acid mine extremophile strain of a Pleurostomophora sp. J. Nat. Prod. 2015, 78, 2917–2923. [Google Scholar] [CrossRef]
- Liu, W.; Chen, S.; Li, J.; Yang, X.; Yan, C.; Liu, H. A new β-tetralonyl glucoside from the Santalum album derived endophytic fungus Colletotrichum sp. GDMU-1. Nat. Prod. Res. 2019, 33, 354–359. [Google Scholar] [CrossRef]
- Evidente, A.; Capretti, P.; Giordano, F.; Surico, G. Identification and phytotoxicity of 3-nitropropanoic acid produced in vitro by Melanconis thelebola. Experientia 1992, 48, 1169–1172. [Google Scholar] [CrossRef]
- Francis, K.; Smitherman, C.; Nishino, S.F.; Spain, J.C.; Gadda, G. The biochemistry of the metabolic poison propionate 3-nitronate and its conjugate acid, 3-nitropropionate. IUBMB Life 2013, 65, 759–768. [Google Scholar] [CrossRef]
- Ayer, W.A.; Attah-Poku, S.K.; Browne, L.M.; Orszanska, H. The chemistry of the blue stain fungi. Part 3. Some metabolites of Ceratocystis minor (Hedgcock) Hunt. Can. J. Chem. 1987, 65, 765–769. [Google Scholar] [CrossRef]
- Kornsakulkarn, J.; Thongpanchang, C.; Lapanun, S.; Srichomthong, K. Isocoumarin glucosides from the scale insect fungus Torrubiella tenuis BCC 12732. J. Nat. Prod. 2009, 72, 1341–1343. [Google Scholar] [CrossRef]
- Chini, A.; Cimmino, A.; Masi, M.; Reveglia, P.; Nocera, P.; Solano, R.; Evidente, A. The fungal phytotoxin lasiojasmonate A activates the plant jasmonic acid pathway. J. Exp. Bot. 2018, 69, 3095–3102. [Google Scholar] [CrossRef]
- Fonseca, S.; Radhakrishnan, D.; Prasad, K.; Chini, A. Fungal production and manipulation of plant hormones. Curr. Med. Chem. 2018, 25, 253–267. [Google Scholar] [CrossRef]
- Tianpanich, K.; Prachya, S.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Radical scavenging and antioxidant activities of isocoumarins and a phthalide from the endophytic fungus Colletotrichum sp. J. Nat. Prod. 2011, 74, 79–81. [Google Scholar] [CrossRef]
- Wei, M.-Y.; Wang, C.-Y.; Liu, Q.-A.; Shao, C.-L.; She, Z.-G.; Lin, Y.-C. Five sesquiterpenoids from a marine-derived fungus Aspergillus sp. isolated from a gorgonian Dichotella gemmacea. Mar. Drugs 2010, 8, 941–949. [Google Scholar] [CrossRef]
- MacMillan, J.; Simpson, T.J. Fungal products. Part V. The absolute stereochemistry of colletodiol and the structures of related metabolites of Colletotrichum capsici. J. Chem. Soc. Perkin Trans. 1 1973, 1487–1493. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Zhi-jun, Y.; Yu, Y.; Zhi-qiang, W.; Tian, Y.; Dai-jie, C. Cytotoxic metabolites of endophytic fungus Colletotrichum sp. from Aristolochia spp. Nat. Prod. Res. Devel. 2012, 24, 329–332. [Google Scholar]
- Sakurai, M.; Kohno, J.; Nishio, M.; Yamamoto, K.; Okuda, T.; Kawano, K.; Nakanishi, N. TMC-205 a new transcriptional up-regulator of SV40 promoter produced by an unidentified fungus fermentation, isolation, physico-chemical properties, structure determination and biological activities. J. Antibiot. 2001, 54, 628–634. [Google Scholar] [CrossRef]
- Dietl, A.-M.; Meir, Z.; Shadkchan, Y.; Osherov, N.; Haas, H. Riboflavin and pantothenic acid biosynthesis are crucial for iron homeostasis and virulence in the pathogenic mold Aspergillus fumigatus. Virulence 2018, 9, 1036–1049. [Google Scholar] [CrossRef]
- Wu, Q.; Guo, Y.; Guo, Z.; Chu, Y.; Wang, T.; Tan, R. Two new cytosporones from the culture of endophytic Phomopsis sp. Chem. Nat. Comp. 2013, 48, 938–941. [Google Scholar] [CrossRef]
- Okamoto, K.; Sakai, T.; Shimada, A.; Shirai, R.; Sakamoto, H.; Yoshida, S.; Ojima, F.; Ishiguro, Y.; Kawagishi, H. Antimicrobial chlorinated orcinol derivatives from mycelia of Hericium erinaceum. Phytochemistry 1993, 34, 1445–1446. [Google Scholar] [CrossRef]
- Buchwaldt, L.; Anderson, K.; Morrall, R.; Gossen, B.; Bernier, C. Identification of lentil germ plasm resistant to Colletotrichum truncatum and characterization of two pathogen races. Phytopathology 2004, 94, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Chongo, G.; Gossen, B.; Bernier, C. Infection by Colletotrichum truncatum in resistant and susceptible lentil genotypes. Can. J. Plant Pathol. 2002, 24, 81–85. [Google Scholar] [CrossRef]
- Latunde-Dada, A.O. Colletotrichum: Tales of forcible entry, stealth, transient confinement and breakout. Mol. Plant Pathol. 2001, 2, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Cerkauskas, R.; Sinclair, J. Use of paraquat to aid detection of fungi in soybean tissues. Phytopathology 1980, 70, 1036–1038. [Google Scholar] [CrossRef]
- Reveglia, P.; Masi, M.; Evidente, A. Melleins—Intriguing natural compounds. Biomolecules 2020, 10, 772. [Google Scholar] [CrossRef]
- Ruijter, G.; Kubicek, C.; Visser, J. Production of organic acids by fungi. In Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 213–230. [Google Scholar]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and Phomentrioloxins B and C produced by diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y. Oxalic acid metabolism contributes to full virulence and pycnidial development in the poplar canker fungus Cytospora chrysosperma. Phytopathology 2020, 110, 1319–1325. [Google Scholar] [CrossRef]
- Tian, X.; Ding, H.; Ke, W.; Wang, L. Quorum sensing in fungal species. Ann. Rev. Microbiol. 2021, 75, 449–469. [Google Scholar] [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef]
- Evidente, A. Bioactive Lipodepsipeptides Produced by Bacteria and Fungi. Int. J. Mol. Sci. 2022, 23, 12342. [Google Scholar] [CrossRef]
- Desjardins, A.; Proctor, R. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P. Mycotoxins affecting animals, foods, humans, and plants: Types, occurrence, toxicities, action mechanisms, prevention, and detoxification strategies—A revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Janevska, S.; Tudzynski, B. Secondary metabolism in Fusarium fujikuroi: Strategies to unravel the function of biosynthetic pathways. App. Microbiol. Biotechnol. 2018, 102, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A. Fungal bioactive macrolides. Nat. Prod. Rep. 2022, 39, 1591–1621. [Google Scholar] [CrossRef]
- Omura, S. Macrolide Antibiotics: Chemistry, Biology, and Practice, 2nd ed.; Academic Press: Tokyo, Japan, 2002. [Google Scholar]
- Lumaret, J.-P.; Errouissi, F.; Floate, K.; Rombke, J.; Wardhaugh, K. A review on the toxicity and non-target effects of macrocyclic lactones in terrestrial and aquatic environments. Curr. Pharm. Biotechnol. 2012, 13, 1004–1060. [Google Scholar] [CrossRef]
- Amusa, N. Production, partial purification and bioassay of toxic metabolites of three plant pathogenic species of Colletotrichum in Nigeria. Mycopathologia 1994, 128, 161–166. [Google Scholar] [CrossRef]
- Kulkarni, S. Studies on the cultural characteristics of Colletotrichum truncatum the causal organism of green gram anthracnose. J. Pharm. Phytochem. 2019, 8, 2366–2369. [Google Scholar]
- Marak, T.; Umbrey, Y.; Mahapatra, S.; Das, S. Cultural, morphological and bio-chemical variability of different isolates of Colletotrichum truncatum causing anthracnose of greengram. Arch. Phytopathol. 2019, 52, 141–154. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Cimmino, A.; Cinelli, T.; Evidente, M.; Masi, M.; Mugnai, L.; Silva, M.A.; Michereff, S.J.; Surico, G.; Evidente, A. Phytotoxic fungal exopolysaccharides produced by fungi involved in grapevine trunk diseases. Nat. Prod. Commun. 2016, 11, 1481–1484. [Google Scholar] [CrossRef]
- Paper, J.M.; Scott-Craig, J.S.; Adhikari, N.D.; Cuomo, C.A.; Walton, J.D. Comparative proteomics of extracellular proteins in vitro and in planta from the pathogenic fungus Fusarium graminearum. Proteomics 2007, 7, 3171–3183. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; de Boves Harrington, P. Chemometric applications in metabolomic studies using chromatography-mass spectrometry. Trends Anal. Chem. 2021, 135, 116165. [Google Scholar] [CrossRef]
- Brereton, R.G.; Lloyd, G.R. Partial least squares discriminant analysis: Taking the magic away. J. Chemom. 2014, 28, 213–225. [Google Scholar] [CrossRef]
- Bittremieux, W.; Wang, M.; Dorrestein, P.C. The critical role that spectral libraries play in capturing the metabolomics community knowledge. Metabolomics 2022, 18, 94. [Google Scholar] [CrossRef]
- Guo, J.; Yu, H.; Xing, S.; Huan, T. Addressing big data challenges in mass spectrometry-based metabolomics. Chem. Commun. 2022, 58, 9979–9990. [Google Scholar] [CrossRef]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of molecular networking and in-silico MS/MS fragmentation for natural products dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Colby, S.M.; Nuñez, J.R.; Hodas, N.O.; Corley, C.D.; Renslow, R.R. Deep learning to generate in silico chemical property libraries and candidate molecules for small molecule identification in complex samples. Anal. Chem. 2019, 92, 1720–1729. [Google Scholar] [CrossRef] [PubMed]
- Dührkop, K.; Nothias, L.-F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C. Systematic classification of unknown metabolites using high-resolution fragmentation mass spectra. Nat. Biotechnol. 2021, 39, 462–471. [Google Scholar] [CrossRef]
- Rutz, A.; Dounoue-Kubo, M.; Ollivier, S.; Bisson, J.; Bagheri, M.; Saesong, T.; Ebrahimi, S.N.; Ingkaninan, K.; Wolfender, J.-L.; Allard, P.-M. Taxonomically informed scoring enhances confidence in natural products annotation. Front. Plant Sci. 2019, 10, 1329. [Google Scholar] [CrossRef]
- Gauglitz, J.M.; West, K.A.; Bittremieux, W.; Williams, C.L.; Weldon, K.C.; Panitchpakdi, M.; Di Ottavio, F.; Aceves, C.M.; Brown, E.; Sikora, N.C. Enhancing untargeted metabolomics using metadata-based source annotation. Nat. Biotechnol. 2022, 40, 1774–1779. [Google Scholar] [CrossRef]
- Allwood, J.W.; Clarke, A.; Goodacre, R.; Mur, L.A. Dual metabolomics: A novel approach to understanding plant–pathogen interactions. Phytochemistry 2010, 71, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ma, R.; Chen, X.L. Advances of metabolomics in fungal pathogen–plant interactions. Metabolites 2019, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Perez de Souza, L.; Alseekh, S.; Naake, T.; Fernie, A. Mass spectrometry-based untargeted plant metabolomics. Curr. Protoc. Plant Biol. 2019, 4, e20100. [Google Scholar] [CrossRef] [PubMed]
- Crandall, S.G.; Gold, K.M.; Jiménez-Gasco, M.D.M.; Filgueiras, C.C.; Willett, D.S. A multi-omics approach to solving problems in plant disease ecology. PLoS ONE 2020, 15, e0237975. [Google Scholar] [CrossRef] [PubMed]
- van Der Hooft, J.J.; Mohimani, H.; Bauermeister, A.; Dorrestein, P.C.; Duncan, K.R.; Medema, M.H. Linking genomics and metabolomics to chart specialized metabolic diversity. Chem. Soc. Rev. 2020, 49, 3297–3314. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Host Plant | Fungal Code |
|---|---|---|
| C. truncatum | Lentil (Lens culinaris) | C428 |
| C. truncatum | Soybean (Glycine max) | C431 |
| C. trifolii | Red clover (Trifolium pretense) | C436 |
| Legume | Plant Specie | Genotype |
|---|---|---|
| Soybean | Glycine max | Creator |
| Faba bean | Vicia faba | Baraca |
| Lentil | Lens culinaris | Pardina |
| Pea | Pisum sativum | Messire |
| Barrel medic | Medicago truncatula | Paraggio |
| Red clover | Trifolium pratense | B1401 |
| Subterranean clover | Trifolium subterraneum | E08 |
| White clover | Trifolium repens | Anteria |
| Compound | C. truncatum C428 | C. truncatum C431 | C. trifolii C436 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PDA | PDB | RICE | RICHARD | PDA | PDB | RICE | RICHARD | PDA | PDB | RICE | RICHARD | |
| Colletochlorin E | n.d. | n.d. | n.d. | 0.01 ± 0.004 | n.d. | n.d. | n.d. | ++ | n.d. | n.d. | n.d. | n.d. |
| Colletopyrone | 0.01 ± 0.001 | 8.13 ± 2.78 | 0.94 ± 0.11 | + | 0.04 ± 0.003 | 14.62 ± 1.32 | 0.04 ± 0.01 | + | + | n.d. | 0.18 ± 0.02 | + |
| 4-Hydroxybenzaldehyde | 0.81 ± 0.66 | 0.05 ± 0.01 | 0.09 ± 0.02 | 0.59 ± 0.04 | 0.19 ± 0.05 | 0.06 ± 0.02 | 0.11 ± 0.02 | 0.74 ± 0.29 | 0.15 ± 0.03 | 0.09 ± 0.006 | 0.19 ± 0.03 | 0.49 ± 0.06 |
| Resorcine | 0.04 ± 0.02 | n.d. | ++ | n.d. | n.d. | n.d. | n.d. | n.d. | ++ | n.d. | n.d. | ++ |
| Orcinol | n.d. | ++ | n.d. | n.d. | 0.02 ± 0.002 | + | n.d. | n.d. | n.d. | + | ++ | n.d. |
| Higginsianin A | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | + | n.d. | n.d. | n.d. |
| Higginsianin B | 0.01 ± 0.002 | ++ | ++ | ++ | ++ | + | ++ | ++ | ++ | ++ | ++ | + |
| 4-Hydroxyphenyl acetic acid | 0.68 ± 0.14 | + | 0.06 ± 0.02 | 0.18 ± 0.03 | 0.09 ± 0.05 | + | 0.21 ± 0.02 | 0.06 ± 0.01 | 0.79 ± 0.60 | n.d. | 0.03 ± 0.002 | 0.29 ± 0.02 |
| 6-Hydroxymellein | + | 0.06 ± 0.004 | 0.30 ± 0.01 | ++ | ++ | 0.06 ± 0.004 | 0.41 ± 0.02 | 0.03 ± 0.003 | + | + | ++ | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reveglia, P.; Agudo-Jurado, F.J.; Barilli, E.; Masi, M.; Evidente, A.; Rubiales, D. Uncovering Phytotoxic Compounds Produced by Colletotrichum spp. Involved in Legume Diseases Using an OSMAC–Metabolomics Approach. J. Fungi 2023, 9, 610. https://doi.org/10.3390/jof9060610
Reveglia P, Agudo-Jurado FJ, Barilli E, Masi M, Evidente A, Rubiales D. Uncovering Phytotoxic Compounds Produced by Colletotrichum spp. Involved in Legume Diseases Using an OSMAC–Metabolomics Approach. Journal of Fungi. 2023; 9(6):610. https://doi.org/10.3390/jof9060610
Chicago/Turabian StyleReveglia, Pierluigi, Francisco J. Agudo-Jurado, Eleonora Barilli, Marco Masi, Antonio Evidente, and Diego Rubiales. 2023. "Uncovering Phytotoxic Compounds Produced by Colletotrichum spp. Involved in Legume Diseases Using an OSMAC–Metabolomics Approach" Journal of Fungi 9, no. 6: 610. https://doi.org/10.3390/jof9060610