
Anastomosis Groups and Mycovirome of Rhizoctonia Isolates Causing Sugar Beet Root and Crown Rot and Their Sensitivity to Flutolanil, Thifluzamide, and Pencycuron
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
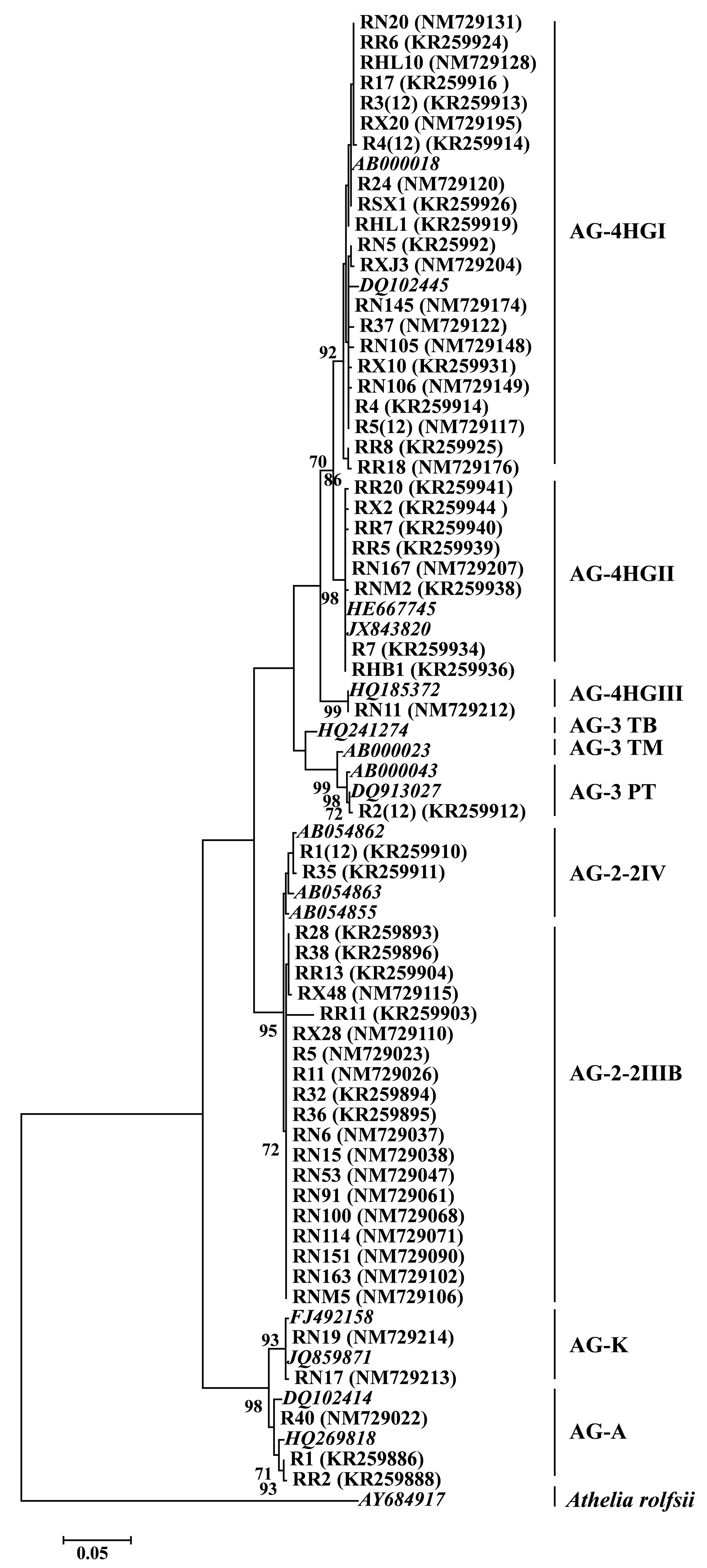
2.1. Rhizoctonia Isolation and Identification
2.2. Pathogenicity Test
2.3. RNA Extraction, Metatranscriptomic Sequencing, and Sequence Analysis
2.4. Open Reading Frame (ORF) Prediction and Phylogenetic Analysis
2.5. Confirmation of Identified Mycoviruses in Rhizoctonia Populations
2.6. Virus Name
2.7. Sensitivity of Rhizoctonia Isolates to Flutolanil, Thifluzamide, and Pencycuron In Vitro
2.8. Statistical Analysis
3. Results
3.1. AGs or Subgroups Determination of Rhizoctonia
3.2. Pathogenicity on Sugar Beet
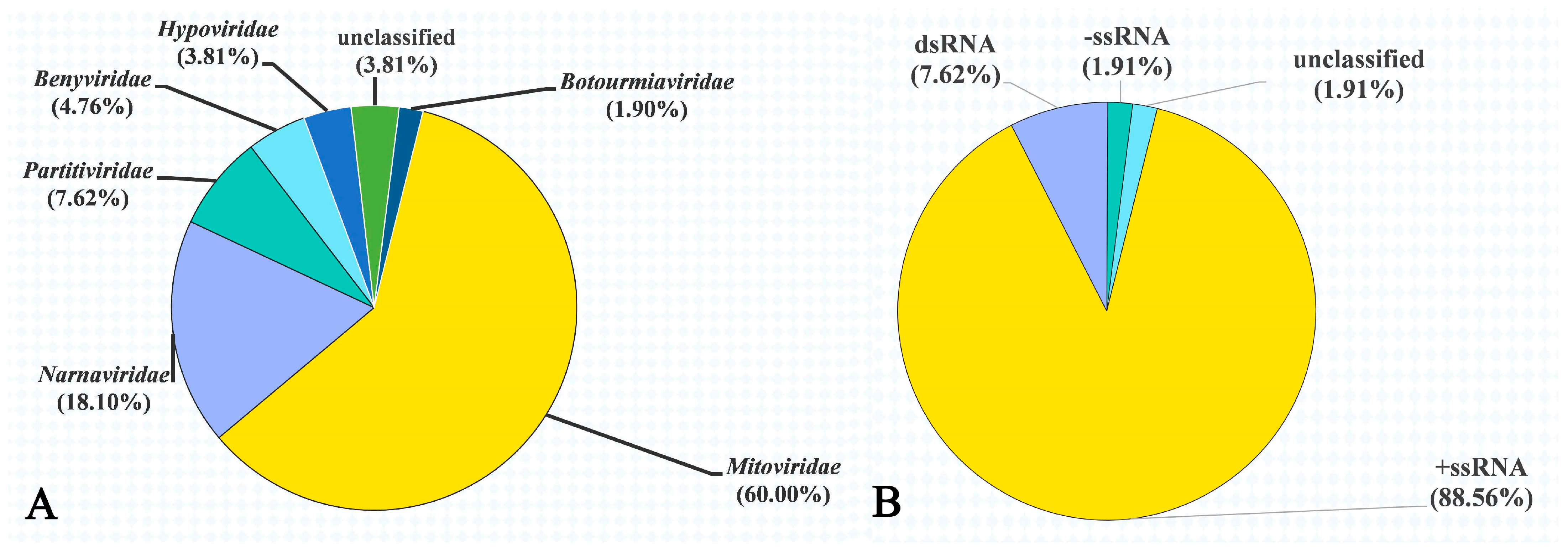
3.3. Mycovirus Diversity in Rhizoctonia Isolates
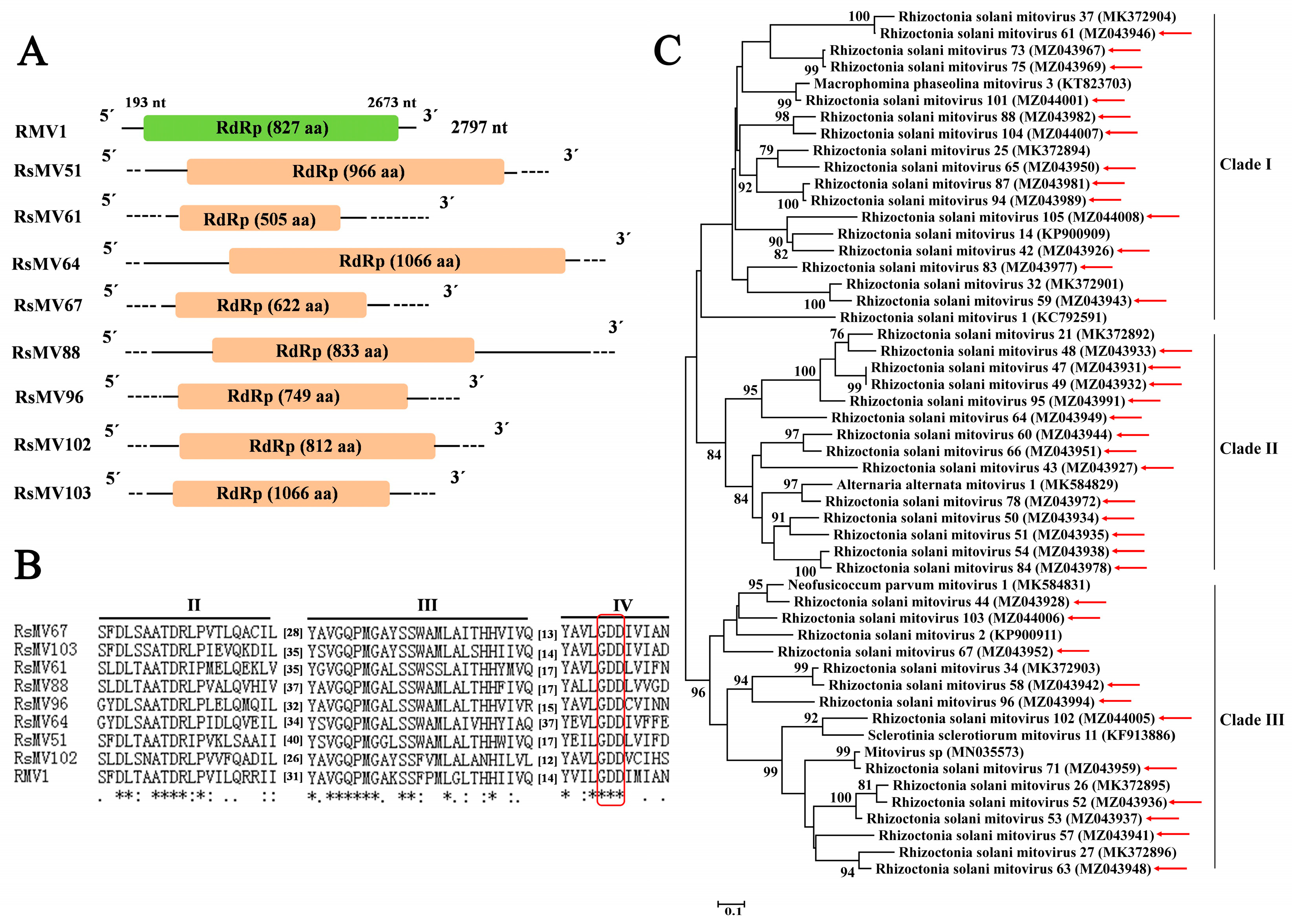
3.4. Sequences Related to Putative Members of the Family Mitoviridae
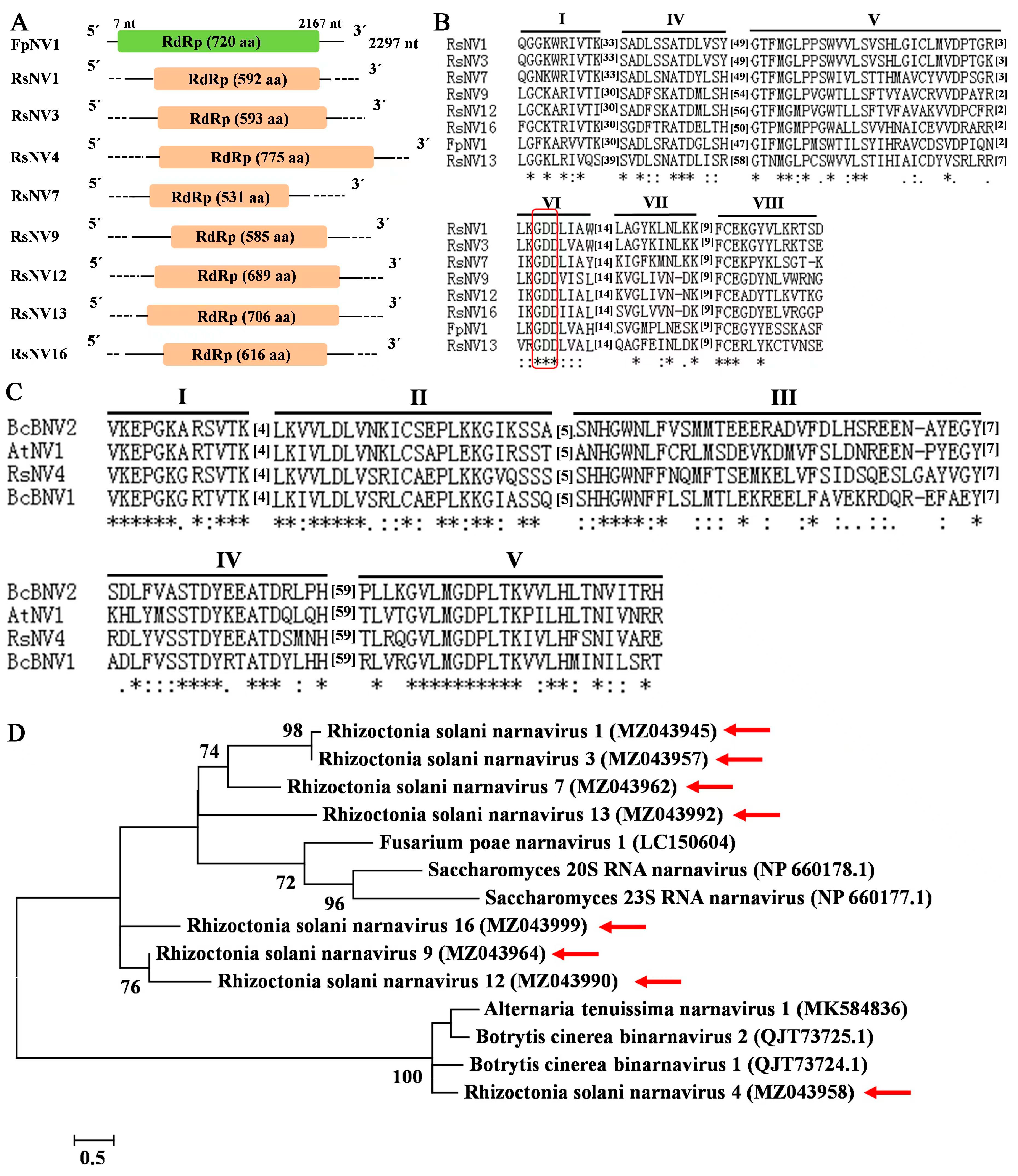
3.5. Sequences Related to Putative Members of the Family Narnaviridae
3.6. Sequences Related to Putative Members of the Family Partitiviridae
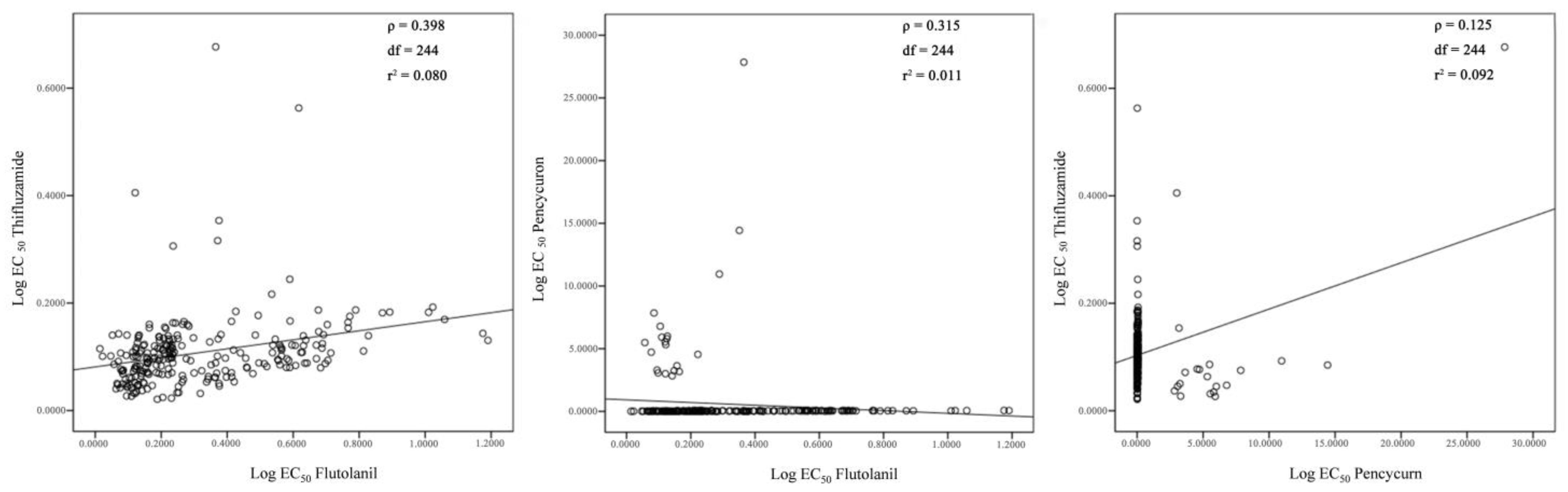
3.7. Sensitivity of 244 Rhizoctonia Isolates to Flutolanil, Thifluzamide, and Pencycuron
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geng, G.; Yang, J. Sugar beet production and industry in China. Sugar Tech. 2015, 17, 13–21. [Google Scholar] [CrossRef]
- Harveson, R.M.; Hanson, L.E.; Hein, G.L. (Eds.) Root diseases caused by fungi and oomycetes. In Compendium of Beet Diseases and Pests, 2nd ed.; American Phytopathological Society: Saint Paul, MN, USA, 2009; pp. 21–24. [Google Scholar]
- Campbell, L.G.; Windels, C.E.; Fugate, K.K.; Brantner, J.R. Postharvest losses associated with severity of Rhizoctonia root and crown rot of sugar beet at harvest. J. Sugar Beet Res. 2014, 51, 31–51. [Google Scholar] [CrossRef]
- Inokuti, E.M.; Thiery-Lanfranchi, D.; Edel-Hermann, V.; Gautheron, N.; Fayolle, L.; Michereff, S.J.; Steinberg, C. Genetic and pathogenic variability of Rhizoctonia solani causing root and crown rot on sugar beet in France. J. Plant Pathol. 2019, 101, 907–916. [Google Scholar] [CrossRef]
- Stojsin, V.; Budakov, D.; Jacobsen, B.; Bagi, F.; Grimme, E.; Neher, O. Analysis of Rhizoctonia solani isolates associated with sugar beet root and crown rot from Serbia. Afr. J. Biotechnol. 2011, 10, 19049–19055. [Google Scholar]
- Berdugo, C.; Hillnhütter, C.; Sikora, R.; Oerke, E.C. A resistance bioassay for Rhizoctonia root and crown rot and damping-off caused by the anastomosis groups AG-2-2IIIB and AG-4 in sugar beet. J. Agric. Sci. Technol. 2012, 2, 294–302. [Google Scholar]
- Taheri, P.; Tarighi, S. Genetic and virulence analysis of Rhizoctonia spp. associated with sugar beet root and crown rot in the Northeast region of Iran. Plant Dis. 2012, 96, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Nagendran, S.; Hammerschmidt, R.; McGrath, J.M. Identification of sugar beet germplasm EL51 as a source of resistance to post-emergence Rhizoctonia damping-off. Eur. J. Plant Pathol. 2009, 123, 461–471. [Google Scholar] [CrossRef]
- Hanson, L.E.; McGrath, J.M. Rhizoctonia seedling disease on sugar beet. Int. Sugar J. 2011, 113, 584–589. [Google Scholar]
- Zhao, C.; Li, Y.; Wu, S.; Wang, P.; Han, C.; Wu, X. Anastomosis group and pathogenicity of Rhizoctonia spp. associated with seedling damping-off of sugar beet in China. Eur. J. Plant Pathol. 2019, 153, 869–878. [Google Scholar] [CrossRef]
- Strausbaugh, C.A.; Eujayl, I.A.; Panella, L.W.; Hanson, L.E. Virulence, distribution and diversity of Rhizoctonia solani from sugar beet in Idaho and Oregon. Can. J. Plant Pathol. 2011, 33, 210–226. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, X.H. First report of sugar beet Rhizoctonia root and crown rot caused by Rhizoctonia solani AG-2-2IIIB in Shanxi province of China. Plant Dis. 2014, 98, 419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, B.; Zhou, H.; Xie, J.; Hao, J.J. First report of Rhizoctonia solani AG4-HG-I infecting sugar beet in China. Plant Dis. 2015, 99, 1185. [Google Scholar] [CrossRef]
- Khan, A.F.; Liu, Y.; Khan, M.F.R. Efficacy and safety of generic azoxystrobin at controlling Rhizoctonia solani in sugar beet. Crop Prot. 2017, 93, 77–81. [Google Scholar] [CrossRef]
- Arabiat, S.; Khan, M.F.R. Sensitivity of Rhizoctonia solani AG-2-2 from sugar beet to fungicides. Plant Dis. 2016, 100, 2427–2433. [Google Scholar] [CrossRef]
- Sharma, P.; Malvick, D.K.; Chanda, A.K. Sensitivity of Rhizoctonia solani anastomosis group 2-2 isolates from soybean and sugar beet to selected SDHI and QoI fungicides. Plant Dis. 2021, 105, 3573–3579. [Google Scholar] [CrossRef]
- Liu, S.; Xie, J.T.; Cheng, J.S.; Li, B.; Chen, T.; Fu, Y.P.; Li, G.Q.; Wang, M.Q.; Jin, H.A.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. USA 2016, 113, 12803–12808. [Google Scholar] [CrossRef]
- Bolton, M.D.; Panella, L.; Campbell, L.; Khan, M.F.R. Temperature, moisture, and fungicide effects in managing Rhizoctonia root and crown rot of sugar beet. Phytopathology 2010, 100, 689–697. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, A.F.; Wang, W.X.; Zhang, Y.; Gao, T.C. Baseline sensitivity and efficacy of thifluzamide in Rhizoctonia solani. Ann. Appl. Biol. 2012, 161, 247–254. [Google Scholar] [CrossRef]
- Li, S.D.; Hou, Y.P.; Peng, D.; Meng, L.; Wang, J.X.; Zhou, M.G.; Chen, C.J. Baseline sensitivity and control efficacy of flutolanil in Rhizoctonia solani. Australas. Plant Pathol. 2014, 43, 313–320. [Google Scholar] [CrossRef]
- Peng, D.; Li, S.D.; Chen, C.J.; Zhou, M.G. Combined application of Bacillus subtilis NJ-18 with fungicides for control of sharp eyespot of wheat. Biol. Control 2014, 70, 28–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, J.L.; Wang, J.X.; Zhou, M.G.; Chen, C.J. Baseline sensitivity and resistance risk assessment of Rhizoctonia cerealis to thifluzamide, a succinate dehydrogenase inhibitor. Pestic. Biochem. Physiol. 2015, 124, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, X.F.; Hua, H.H.; Han, C.G.; Wu, X.H. Sensitivity of Rhizoctonia spp. to flutolanil and characterization of the point mutation in succinate dehydrogenase conferring fungicide resistance. Eur. J. Plant Pathol. 2019, 155, 13–23. [Google Scholar] [CrossRef]
- Zhao, C.; Li, Y.T.; Liang, Z.J.; Gao, L.H.; Han, C.G.; Wu, X.H. Molecular mechanisms associated with the resistance of Rhizoctonia solani AG-4 isolates to the succinate dehydrogenase inhibitor, thifluzamide. Phytopathology 2022, 112, 567–578. [Google Scholar] [CrossRef]
- Evan, E.; Rush, M.C.; Blackwell, M. Effects of rate and timing of fungicide applications on incidence and severity of sheath blight and grain yield of rice. Plant Dis. 1991, 75, 1254–1261. [Google Scholar]
- Moszczynska, E.; Pytlarz-Kozicka, M. The effect of pencycuron on health of potato tubers. Przem. Chem. 2017, 96, 406–408. [Google Scholar]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.H.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Y.Q.; Dong, W.H.; Sun, A.L.; Chen, N.W.; Zhao, Z.F.; Li, Y.Q.; Li, C.Y.; Yang, G.H. Molecular characterization of a novel mycovirus isolated from Rhizoctonia solani AG-1 IA strain 9-11. Arch. Virol. 2021, 166, 3229–3232. [Google Scholar] [CrossRef]
- Zhang, M.L.; He, Z.R.; Huang, X.T.; Shu, C.W.; Zhou, E.X. Genome organizations and functional analyses of a novel gammapartitivirus from Rhizoctonia solani AG-1 IA strain D122. Viruses 2021, 13, 2254. [Google Scholar] [CrossRef]
- Li, S.W.; Li, Y.T.; Hu, C.H.; Han, C.G.; Zhou, T.; Zhao, C.; Wu, X.H. Full genome sequence of a new mitovirus from the phytopathogenic fungus Rhizoctonia solani. Arch. Virol. 2020, 165, 1719–1723. [Google Scholar] [CrossRef] [PubMed]
- Bartholomaus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed]
- Bartholomaus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Identification of a novel mycovirus isolated from Rhizoctonia solani (AG-2-2 IV) provides further information about genome plasticity within the order Tymovirales. Arch. Virol. 2017, 162, 555–559. [Google Scholar] [CrossRef]
- Picarelli, M.A.S.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica from Brazil. Front. Cell Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Novel mitoviruses in Rhizoctonia solani AG-3 PT infecting potato. Fungal Biol. 2016, 120, 338–350. [Google Scholar] [CrossRef]
- Li, Y.T.; Li, S.W.; Zhao, Y.M.; Zhou, T.; Wu, X.H.; Zhao, C. Six novel mycoviruses containing positive single-stranded RNA and double-stranded RNA genomes co-infect a single strain of the Rhizoctonia solani AG-3 PT. Viruses 2022, 14, 813. [Google Scholar] [CrossRef]
- Li, Y.T.; Li, S.W.; Liang, Z.J.; Cai, Q.N.; Zhou, T.; Zhao, C.; Wu, X.H. RNA-seq analysis of Rhizoctonia solani AG-4HGI strain BJ-1H infected by a new viral strain of Rhizoctonia solani partitivirus 2 reveals a potential mechanism for hypovirulence. Phytopathology 2022, 112, 1373–1385. [Google Scholar] [CrossRef]
- Li, Y.Q.; Chen, W.R.; Niu, Y.F.; Xu, P.; Zhang, L.F.; Yu, S.H.; Yang, G.H.; Mo, X.H. Complete nucleotide sequence of a novel alphapartitivirus from Rhizoctonia solani AG-4HGIII isolate SM03. Arch. Virol. 2022, 167, 953–957. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. Complete genome sequence of a novel mitovirus from the phytopathogenic fungus Rhizoctonia oryzae-sativae. Arch. Virol. 2017, 162, 1409–1412. [Google Scholar] [CrossRef]
- Zhang, T.; Li, W.; Chen, H.G.; Yu, H.S. Full genome sequence of a putative novel mitovirus isolated from Rhizoctonia cerealis. Arch. Virol. 2015, 160, 1815–1818. [Google Scholar] [CrossRef]
- Li, Y.Q.; Xu, P.; Zhang, L.F.; Xia, Z.Y.; Qin, X.Y.; Yang, G.H.; Mo, X.H. Molecular characterization of a novel mycovirus from Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2015, 160, 2371–2374. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and future perspectives in virus discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar] [CrossRef]
- He, Z.R.; Huang, X.T.; Fan, Y.; Yang, M.; Zhou, E.X. Metatranscriptomic analysis reveals rich mycoviral diversity in three major fungal pathogens of rice. Int. J. Mol. Sci. 2022, 23, 9192. [Google Scholar] [CrossRef]
- Li, Y.T.; Yang, N.B.; Mu, T.Y.; Wu, X.H.; Zhao, C. Diversity of mycoviruses present in strains of binucleate Rhizoctonia and multinucleate Rhizoctonia, causal agents for potato stem canker or black scurf. J. Fungi 2023, 9, 214. [Google Scholar] [CrossRef]
- Sun, H.Y.; Lu, C.Q.; Li, W.; Deng, Y.Y.; Chen, H.G. Homozygous and heterozygous point mutations in succinate dehydrogenase subunits b, c and d of Rhizoctonia cerealis conferring resistance to thifluzamide. Pest Manag. Sci. 2017, 73, 896–903. [Google Scholar] [CrossRef]
- Harveson, R.M. Evaluation of fumigation and Rhizomania-tolerant cultivars for control of a root disease complex of sugar beets. Plant Dis. 1994, 78, 1197. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Crucitti, D.; Chiapello, M.; Oliva, D.; Forgia, M.; Turina, M.; Carimi, F.; La Bella, F.; Pacifico, D. Identification and molecular characterization of novel mycoviruses in Saccharomyces and Non-Saccharomyces yeasts of oenological interest. Viruses 2021, 14, 52. [Google Scholar] [CrossRef]
- Ruiz-Padilla, A.; Rodríguez-Romero, J.; Gómez-Cid, I.; Pacifico, D.; Ayllón, M.A. Novel mycoviruses discovered in the mycovirome of a necrotrophic fungus. mBio 2021, 12, e03705-20. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Chiapello, M.; Rodriguez-Romero, J.; Ayllon, M.A.; Turina, M. Analysis of the virome associated to grapevine downy mildew lesions reveals new mycovirus lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, M.L.; Chen, Q.G.; Zhu, M.H.; Zhou, E.X. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456, 220–226. [Google Scholar] [CrossRef]
- Xu, S.S.; Liu, X.R.; Zhang, J.; Xiao, L.Y.; Zhao, L.J.; Li, C.Y.; Dong, W.H.; Yang, G.H. Morphological and molecular characterization of Rhizoctonia solani AG-3 associated with tobacco target leaf spot in China. J. Basic Microbiol. 2023, 63, 200–209. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Anastomosis group and pathogenicity of Rhizoctonia solani associated with stem canker and black scurf of potato in China. Eur. J. Pathol. 2015, 143, 99–111. [Google Scholar] [CrossRef]
- Nerva, L.; Vigani, G.; di Silvestro, D.; Ciuffo, M.; Forgia, M.; Chitarra, W.; Turina, M. Biological and molecular characterization of Chenopodium quinoa mitovirus 1 reveals a distinct small RNA response compared to those of cytoplasmic RNA viruses. J. Virol. 2019, 93, e01998-18. [Google Scholar] [CrossRef]
- Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel mitoviruses and a unique Tymo-like virus in hypovirulent and virulent strains of the Fusarium head blight fungus, Fusarium boothii. Viruses 2018, 10, 584. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Wu, S.S.; Liu, L.J.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef]
- Hillman, B.I.; Esteban, R. Family Narnaviridae. In Virus Taxonomy: Ninth Report of the International Committee for the Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 1025–1030. [Google Scholar]
- Lin, Y.; Zhou, J.; Zhou, X.; Shuai, S.; Zhou, R.; An, H.; Fang, S.; Zhang, S.; Deng, Q. A novel narnavirus from the plant-pathogenic fungus Magnaporthe oryzae. Arch. Virol. 2020, 165, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Kamer, G.; Argos, P. Primary structural comparison of RNA-dependent polymerases from plant, animal and bacterial viruses. Nucleic Acids Res. 1984, 12, 7269–7282. [Google Scholar] [CrossRef] [PubMed]
- Poch, O.; Sauvaget, I.; Delarue, M.; Tordo, N. Identification of four conserved motifs among the RNA-dependent polymerase encoding elements. EMBO J. 1989, 8, 3867–3874. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.H.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.T.; Nibert, M.L.; Consortium, I.R. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Wang, J.X.; Yang, B.; Wang, Q.R.; Zhou, J.J.; Yu, W.F. Molecular characterization of a debilitation-associated partitivirus infecting the pathogenic fungus Aspergillus flavus. Front. Microbiol. 2019, 10, 626. [Google Scholar] [CrossRef]
- Nerva, L.; Silvestri, A.; Ciuffo, M.; Palmano, S.; Varese, G.C.; Turina, M. Transmission of Penicillium aurantiogriseum partiti-like virus 1 to a new fungal host (Cryphonectria parasitica) confers higher resistance to salinity and reveals adaptive genomic changes. Environ. Microbiol. 2017, 19, 4480–4492. [Google Scholar] [CrossRef]
- Chen, Y.; Gai, X.T.; Chen, R.X.; Li, C.X.; Zhao, G.K.; Xia, Z.Y.; Zou, C.M.; Zhong, J. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Molecular characterization of an endornavirus from Rhizoctonia solani AG-3 PT infecting potato. Fungal Biol. 2014, 118, 924–934. [Google Scholar] [CrossRef]
- Campion, C.; Chatot, C.; Perraton, B.; Andrivon, D. Anastomosis groups, pathogenicity and sensitivity to fungicides of Rhizoctonia solani isolates collected on potato crops in France. Eur. J. Plant Pathol. 2003, 109, 983–992. [Google Scholar] [CrossRef]
- Brenneman, T.B.; Sumner, D.R.; Baird, R.E.; Burton, G.W.; Minton, N.A. Suppression of foliar and soilborne peanut diseases in bahiagrass rotations. Phytopathology 1995, 85, 948–952. [Google Scholar] [CrossRef]
- Koehler, A.M.; Shew, H.D. Seasonal dynamics and fungicide sensitivity of organisms causing brown patch of tall fescue in North Carolina. Mycologia 2017, 109, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Tsror, L. Biology, epidemiology and management of Rhizoctonia solani on potato. J. Phytopathol. 2010, 158, 649–658. [Google Scholar] [CrossRef]
- Virgen-Calleros, G.; Olalde-Portugal, V.; Carling, D.E. Anastomosis groups of Rhizoctonia solani on potato in central Mexico and potential for biological and chemical control. Am. J. Potato Res. 2000, 77, 219–224. [Google Scholar] [CrossRef]
- Kanetis, L.; Tsimouris, D.; Christoforou, M. Characterization of Rhizoctonia solani associated with black scurf in Cyprus. Plant Dis. 2016, 100, 1591–1598. [Google Scholar] [CrossRef]
- Niu, Y.H.; Yuan, Y.Z.; Mao, J.L.; Yang, Z.; Cao, Q.W.; Zhang, T.F.; Wang, S.Q.; Liu, D.L. Characterization of two novel mycoviruses from Penicillium digitatum and the related fungicide resistance analysis. Sci. Rep. 2018, 8, 5513. [Google Scholar] [CrossRef]
- Wang, S.Q.; Yang, Z.; Zhang, T.F.; Li, N.; Cao, Q.W.; Li, G.Q.; Yuan, Y.Z.; Liu, D.L. Molecular characterization of a chrysovirus isolated from the citrus pathogen Penicillium crustosum and related fungicide resistance analysis. Front. Cell Infect. Microbiol. 2019, 9, 156. [Google Scholar] [CrossRef]
- Ma, G.P.; Zhang, X.F.; Hua, H.H.; Zhou, T.; Wu, X.H. Molecular and biological characterization of a novel strain of Alternaria alternata chrysovirus 1 identified from the pathogen Alternaria tenuissima causing watermelon leaf blight. Virus Res. 2020, 280, 197904. [Google Scholar] [CrossRef]
- Liang, Z.J.; Hua, H.H.; Wu, C.Y.; Zhou, T.; Wu, X.H. A botybirnavirus isolated from Alternaria tenuissima confers hypervirulence and decreased sensitivity of its host fungus to difenoconazole. Viruses 2022, 14, 2093. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Origin | Number of Isolates in Different Anastomosis Groups (AGs) or Subgroups of Rhizoctonia | |||||||
|---|---|---|---|---|---|---|---|---|
| AG-A | AG-K | AG-2-2IIIB | AG-2-2IV | AG-3 PT | AG-4HGI | AG-4HGII | AG-4HGIII | |
| Beijing municipality | 0 | 0 | 0 | 1 (1) z | 0 | 0 | 0 | 0 |
| Gansu province | 0 | 0 | 0 | 0 | 0 | 1 (1) | 1 (1) | 0 |
| Hebei province | 0 | 0 | 0 | 0 | 0 | 0 | 4 (1) | 0 |
| Heilongjiang province | 5 (5) | 0 | 4 (3) | 1 (1) | 1 (1) | 18 (5) | 3 (2) | 0 |
| Inner Mongolia autonomous region | 0 | 2 (2) | 93 (12) | 0 | 0 | 50 (7) | 8 (3) | 1 (1) |
| Jilin province | 0 | 0 | 1 (1) | 0 | 0 | 0 | 0 | 0 |
| Shanxi province | 0 | 0 | 2 (2) | 0 | 0 | 7 (2) | 0 | 0 |
| Xinjiang Uygur autonomous region | 0 | 0 | 7 (4) | 0 | 0 | 32 (6) | 1 (1) | 0 |
| Total | 5 (5) | 2 (2) | 107 (22) | 2 (2) | 1 (1) | 108 (21) | 18 (8) | 1 (1) |
| Ratio (%) | 2.05 | 0.82 | 43.85 | 0.82 | 0.41 | 44.26 | 7.38 | 0.41 |
| AGs or Subgroups of Rhizoctonia | Disease Incidence (%) y | Disease Index y |
|---|---|---|
| AG-A | 29.10 ± 6.28 b | 19.98 ± 3.87 b |
| AG-K | 54.35 ± 24.55 b | 16.82 ± 7.80 b |
| AG-2-2IIIB | 80.83 ± 6.60 ab | 61.57 ± 4.59 a |
| AG-2-2IV | 95.70 ± 4.30 a | 68.20 ± 18.20 a |
| AG-3 PT z | 0.00 | 0.00 |
| AG-4HGI | 96.69 ± 1.42 a | 73.85 ± 3.39 a |
| AG-4HGII | 98.12 ± 1.26 a | 68.70 ± 10.29 a |
| AG-4HGIII z | 100.00 | 72.73 |
| Contig ID | Size (Amino Acid) | Mycovirus Name | Best Match | Identity (Amino Acid) | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|
| Contig 190 | 1001 | Rhizoctonia solani mitovirus 42 | Rhizoctonia solani mitovirus 14 | 57.91% | 70% | 0.0 | MZ043926 |
| Contig 195 | 879 | Rhizoctonia solani mitovirus 43 | Epicoccum nigrum mitovirus 1 | 53.20% | 66% | 0.0 | MZ043927 |
| Contig 249 | 697 | Rhizoctonia solani mitovirus 44 | Neofusicoccum parvum mitovirus 1 | 88.52% | 78% | 0.0 | MZ043928 |
| Contig 370 | 898 | Rhizoctonia solani mitovirus 47 | Macrophomina phaseolina mitovirus 3 | 41.43% | 80% | 0.0 | MZ043931 |
| Contig 466 | 1066 | Rhizoctonia solani mitovirus 48 | Rhizoctonia solani mitovirus 21 | 58.70% | 70% | 0.0 | MZ043932 |
| Contig 468 | 1041 | Rhizoctonia solani mitovirus 49 | Rhizoctonia solani mitovirus 21 | 76.89% | 71% | 0.0 | MZ043933 |
| Contig 508 | 967 | Rhizoctonia solani mitovirus 50 | Mitovirus sp. | 42.71% | 57% | 2 × 10−179 | MZ043934 |
| Contig 509 | 966 | Rhizoctonia solani mitovirus 51 z | Mitovirus sp. | 43.61% | 57% | 0.0 | MZ043935 |
| Contig 538 | 781 | Rhizoctonia solani mitovirus 52 | Rhizoctonia solani mitovirus 26 | 84.54% | 89% | 0.0 | MZ043936 |
| Contig 539 | 783 | Rhizoctonia solani mitovirus 53 | Rhizoctonia solani mitovirus 26 | 85.06% | 77% | 0.0 | MZ043937 |
| Contig 604 | 844 | Rhizoctonia solani mitovirus 54 | Mitovirus sp. | 48.70% | 51% | 5 × 10−168 | MZ043938 |
| Contig 851 | 811 | Rhizoctonia solani mitovirus 57 | Rhizoctonia solani mitovirus 8 | 72.80% | 81% | 0.0 | MZ043941 |
| Contig 997 | 869 | Rhizoctonia solani mitovirus 58 | Rhizoctonia solani mitovirus 34 | 80.92% | 76% | 0.0 | MZ043942 |
| Contig 1038 | 887 | Rhizoctonia solani mitovirus 59 | Rhizoctonia solani mitovirus 32 | 70.90% | 64% | 0.0 | MZ043943 |
| Contig 1104 | 850 | Rhizoctonia solani mitovirus 60 | Epicoccum nigrum mitovirus 1 | 75.54% | 70% | 0.0 | MZ043944 |
| Contig 1212 | 505 | Rhizoctonia solani mitovirus 61 z | Rhizoctonia solani mitovirus 37 | 77.25% | 83% | 0.0 | MZ043946 |
| Contig 1830 | 855 | Rhizoctonia solani mitovirus 63 | Rhizoctonia solani mitovirus 27 | 77.09% | 76% | 0.0 | MZ043948 |
| Contig 2377 | 1066 | Rhizoctonia solani mitovirus 64 z | Rhizoctonia solani mitovirus 1 | 72.18% | 35% | 0.0 | MZ043949 |
| Contig 3007 | 835 | Rhizoctonia solani mitovirus 65 | Rhizoctonia solani mitovirus 25 | 62.19% | 65% | 0.0 | MZ043950 |
| Contig 3241 | 838 | Rhizoctonia solani mitovirus 66 | Epicoccum nigrum mitovirus 1 | 85.00% | 71% | 0.0 | MZ043951 |
| Contig 3312 | 622 | Rhizoctonia solani mitovirus 67 z | Neofusicoccum parvum mitovirus 1 | 46.21% | 87% | 0.0 | MZ043952 |
| Contig 10147 | 804 | Rhizoctonia solani mitovirus 71 | Mitovirus sp. | 89.66% | 77% | 0.0 | MZ043959 |
| First_Contig 14 | 715 | Rhizoctonia solani mitovirus 73 | Macrophomina phaseolina mitovirus 3 | 42.95% | 82% | 5 × 10−175 | MZ043967 |
| First_Contig 20 | 715 | Rhizoctonia solani mitovirus 75 | Macrophomina phaseolina mitovirus 3 | 42.97% | 80% | 1 × 10−169 | MZ043969 |
| First_Contig 27 | 847 | Rhizoctonia solani mitovirus 78 | Alternaria alternata mitovirus 1 | 80.87% | 71% | 0.0 | MZ043972 |
| First_Contig 139 | 949 | Rhizoctonia solani mitovirus 83 | Rhizoctonia solani mitovirus 7 | 70.06% | 72% | 0.0 | MZ043977 |
| First_Contig 203 | 876 | Rhizoctonia solani mitovirus 84 | Epicoccum nigrum mitovirus 1 | 47.54% | 57% | 8 × 10−161 | MZ043978 |
| First_Contig 275 | 864 | Rhizoctonia solani mitovirus 87 | Rhizoctonia solani mitovirus 25 | 47.64% | 77% | 0.0 | MZ043981 |
| First_Contig 345 | 833 | Rhizoctonia solani mitovirus 88 z | Rhizoctonia solani mitovirus 33 | 62.34% | 21% | 2 × 10−122 | MZ043982 |
| First_Contig 740 | 730 | Rhizoctonia solani mitovirus 94 | Rhizoctonia solani mitovirus 25 | 45.35% | 93% | 0.0 | MZ043989 |
| First_Contig 919 | 809 | Rhizoctonia solani mitovirus 95 | Rhizoctonia solani mitovirus 21 | 55.36% | 90% | 0.0 | MZ043991 |
| First_Contig 1173 | 749 | Rhizoctonia solani mitovirus 96 z | Rhizoctonia solani mitovirus 11 | 41.36% | 79% | 2 × 10−164 | MZ043994 |
| Second_Contig 58 | 785 | Rhizoctonia solani mitovirus 101 | Macrophomina phaseolina mitovirus 3 | 86.19% | 93% | 0.0 | MZ044001 |
| Contig 11664 | 812 | Rhizoctonia solani mitovirus 102 z | Sclerotinia sclerotiorum mitovirus 11 | 74.05% | 38% | 0.0 | MZ044005 |
| Contig 163 | 731 | Rhizoctonia solani mitovirus 103 z | Rhizoctonia solani mitovirus 2 | 56.04% | 75% | 0.0 | MZ044006 |
| Contig 1141 | 803 | Rhizoctonia solani mitovirus 104 | Rhizoctonia solani mitovirus 25 | 48.38% | 66% | 3 × 10−175 | MZ044007 |
| Contig 1714 | 866 | Rhizoctonia solani mitovirus 105 | Rhizoctonia solani mitovirus 14 | 52.10% | 69% | 0.0 | MZ044008 |
| Contig ID | Size (Amino Acid) | Mycovirus Name | Best Match | Identity (Amino Acid) | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|
| Contig 1113 | 592 | Rhizoctonia solani narnavirus 1 | Fusarium poae narnavirus 1 | 30.83% | 45% | 4 × 10−27 | MZ043945 |
| Contig 8527 | 593 | Rhizoctonia solani narnavirus 3 | Fusarium poae narnavirus 1 | 30.71% | 44% | 3 × 10−28 | MZ043957 |
| Contig 8560 | 775 | Rhizoctonia solani narnavirus 4 | Alternaria tenuissima narnavirus 1 | 45.55% | 88% | 0.0 | MZ043958 |
| Contig 11100 | 531 | Rhizoctonia solani narnavirus 7 | Fusarium poae narnavirus 1 | 31.72% | 45% | 1 × 10−24 | MZ043962 |
| Contig 19187 | 585 | Rhizoctonia solani narnavirus 9 | Fusarium poae narnavirus 1 | 28.98% | 54% | 8 × 10−30 | MZ043964 |
| First_Contig 917 | 689 | Rhizoctonia solani narnavirus 12 | Fusarium poae narnavirus 1 | 28.68% | 60% | 1 × 10−36 | MZ043990 |
| First_Contig 1098 | 706 | Rhizoctonia solani narnavirus 13 | Fusarium poae narnavirus 1 | 28.29% | 64% | 3 × 10−18 | MZ043992 |
| First_Contig 5037 | 616 | Rhizoctonia solani narnavirus 16 | Fusarium poae narnavirus 1 | 28.96% | 55% | 3 × 10−34 | MZ043999 |
| Contig ID | Size (Amino Acid) | Mycovirus Name | Best Match | Identity (Amino Acid) | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|
| Contig 3529 | 623 | Rhizoctonia solani partitivirus 2 strain beet | Rhizoctonia solani dsRNA virus 2 | 99.52% | 95% | 0.0 | MZ043917 |
| Contig 3073 | 595 | Rhizoctonia solani partitivirus 15 | Rhizoctonia solani partitivirus 8 | 67.68% | 94% | 0.0 | MZ043918 |
| Contig 14630 | 630 | Rhizoctonia solani partitivirus 18 | Fusarium solani partitivirus 2 | 47.52% | 88% | 0.0 | MZ043921 |
| AGs or Subgroups of Rhizoctonia | Number of Isolates | Mean of EC50 (μg·mL−1) of Fungicide on Rhizoctonia y | ||
|---|---|---|---|---|
| Flutolanil | Thifluzamide | Pencycuron | ||
| AG-A | 5 | 0.1568 ± 0.1235 b | 0.2797 ± 0.2588 a | 8.8470 ± 10.6742 b |
| AG-K | 2 | 0.3201 ± 0.0440 ab | 0.0889 ± 0.0055 c | 12.6914 ± 2.4621 a |
| AG-2-2IIIB | 107 | 0.5071 ± 0.2288 a | 0.1200 ± 0.0714 b | 0.0372 ± 0.0200 d |
| AG-2-2IV | 2 | 0.3220 ± 0.0651 ab | 0.0828 ± 0.0189 c | 0.0124 ± 0.0061 d |
| AG-3 PT z | 1 | 0.2330 | 0.0979 | 0.0109 |
| AG-4HGI | 108 | 0.1754 ± 0.0833 b | 0.0958 ± 0.0328 c | 0.0623 ± 0.3142 d |
| AG-4HGII | 17 | 0.1309 ± 0.0393 b | 0.0728 ± 0.0402 c | 3.5671 ± 2.7024 c |
| AG-4HGIII z | 1 | 0.1288 | 0.0476 | 0.0238 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Li, S.; Ma, Z.; Wang, W.; Gao, L.; Han, C.; Yang, A.; Wu, X. Anastomosis Groups and Mycovirome of Rhizoctonia Isolates Causing Sugar Beet Root and Crown Rot and Their Sensitivity to Flutolanil, Thifluzamide, and Pencycuron. J. Fungi 2023, 9, 545. https://doi.org/10.3390/jof9050545
Zhao C, Li S, Ma Z, Wang W, Gao L, Han C, Yang A, Wu X. Anastomosis Groups and Mycovirome of Rhizoctonia Isolates Causing Sugar Beet Root and Crown Rot and Their Sensitivity to Flutolanil, Thifluzamide, and Pencycuron. Journal of Fungi. 2023; 9(5):545. https://doi.org/10.3390/jof9050545
Chicago/Turabian StyleZhao, Can, Siwei Li, Zhihao Ma, Wenjun Wang, Lihong Gao, Chenggui Han, Anpei Yang, and Xuehong Wu. 2023. "Anastomosis Groups and Mycovirome of Rhizoctonia Isolates Causing Sugar Beet Root and Crown Rot and Their Sensitivity to Flutolanil, Thifluzamide, and Pencycuron" Journal of Fungi 9, no. 5: 545. https://doi.org/10.3390/jof9050545