
Engineering a Phosphoketolase Pathway to Supplement Cytosolic Acetyl-CoA in Aspergillus niger Enables a Significant Increase in Citric Acid Production
 ,
,
Abstract
:1. Introduction

2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. RNA Isolation and RT-PCR Analysis of acs, ack, pkA and pkB
2.3. Plasmid Construction
2.4. Construction of Strains
2.5. Citric Acid Shake-Flask Fermentation and Bioreactor Fermentation
2.6. Intracellular Acetyl-CoA Concentration Measurement
2.7. HPLC Analysis of Citric Acid and Acetic Acid
2.8. Determination of the Cell Dry Weight and Residual Sugar
2.9. Enzyme Activity Assays of PK and PTA
2.10. Statistical Analysis
3. Results
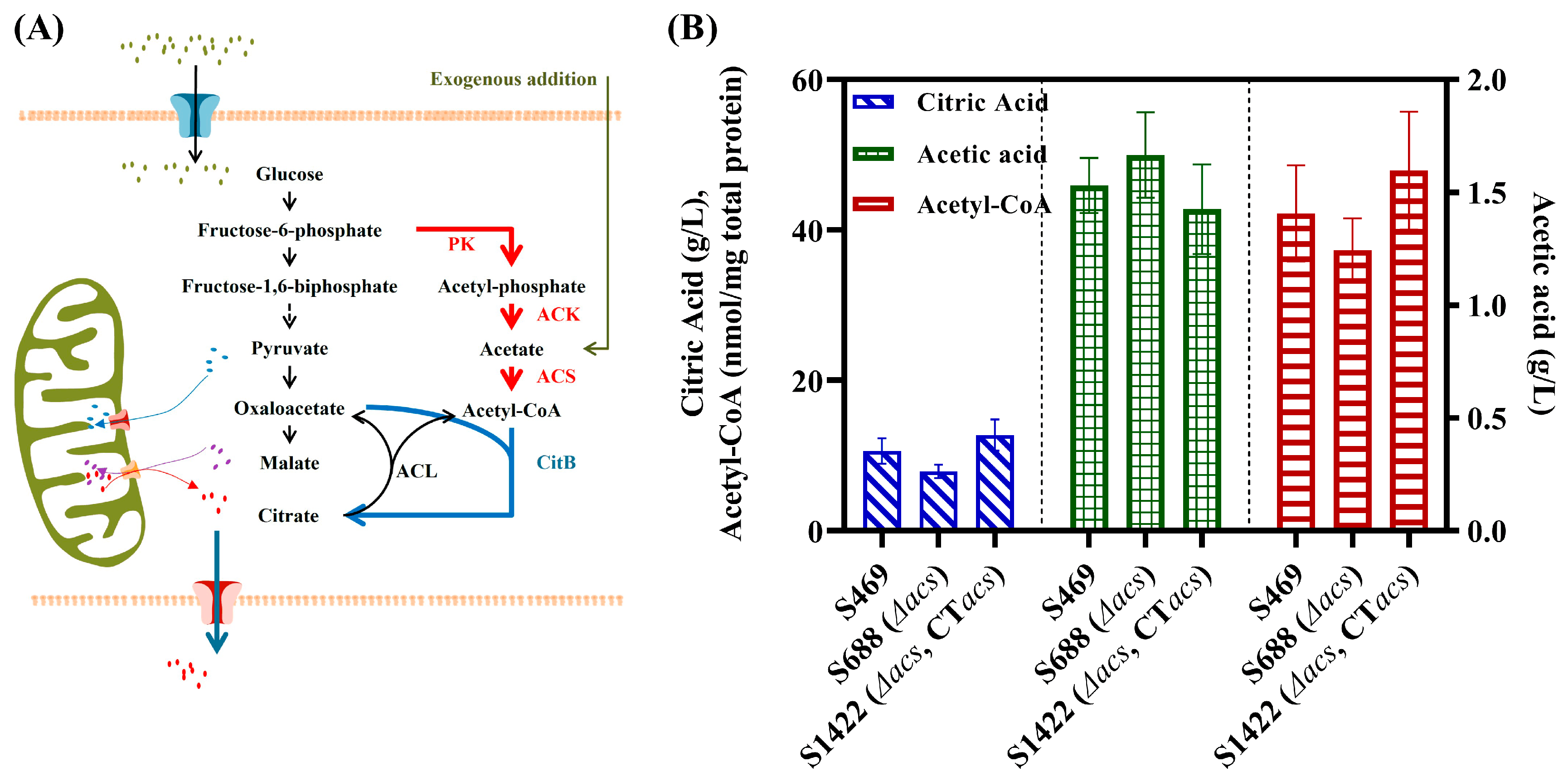
3.1. Role of ACS in the Biosynthesis of Acetyl-CoA and Citric Acid in A. niger
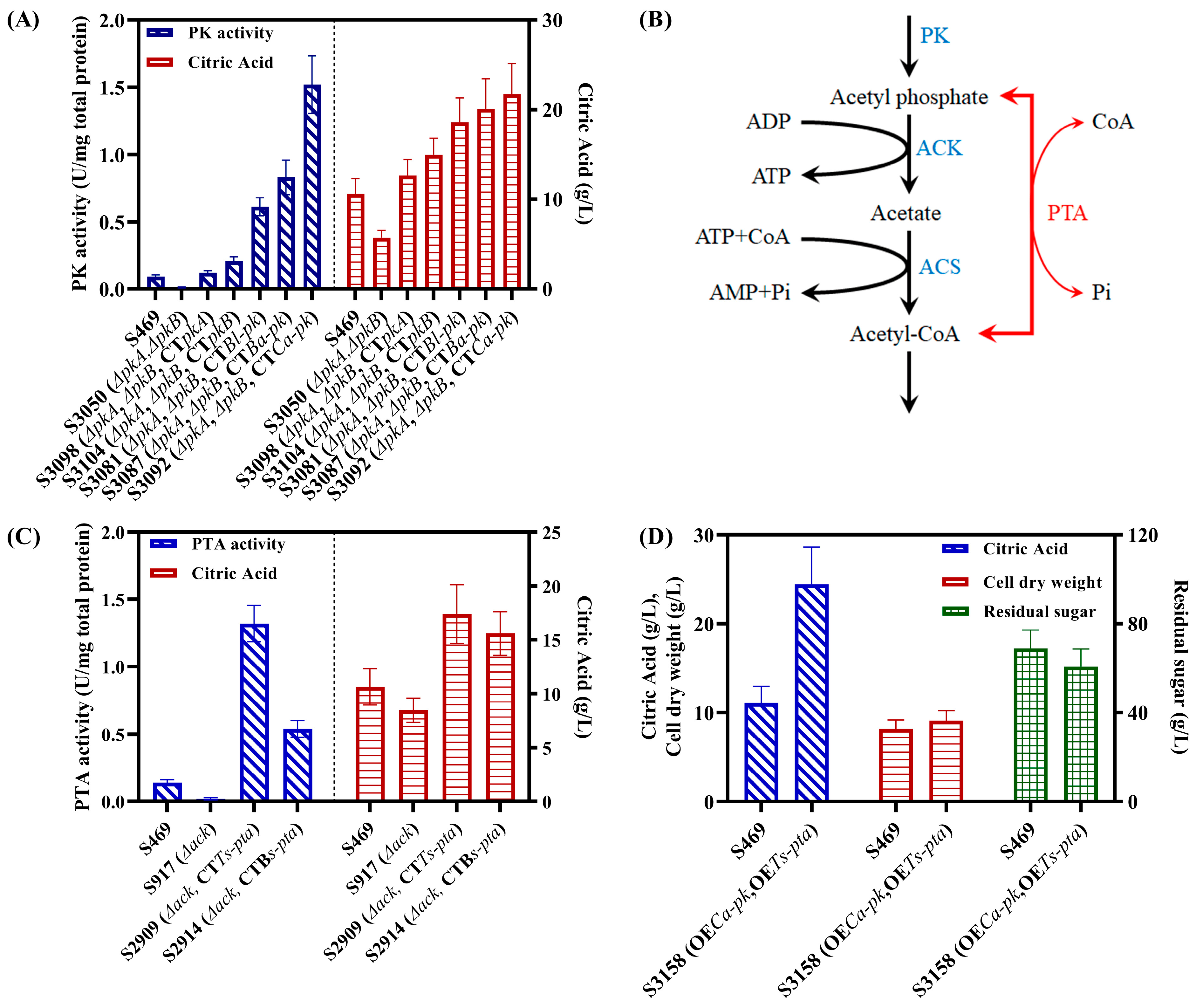
3.2. Roles of PKs and ACK in the Biosynthesis of Acetyl-CoA and Citric Acid in A. niger
3.3. Heterologous PK-PTA Pathway Can Significantly Increase the Citrate Production of A. niger
3.4. Citric Acid Fermentation in a 2 L Bioreactor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Upton, D.J.; McQueen-Mason, S.J.; Wood, A.J. An accurate description of Aspergillus niger organic acid batch fermentation through dynamic metabolic modelling. Biotechnol. Biofuels 2017, 10, 258. [Google Scholar] [CrossRef]
- Hu, W.; Li, W.J.; Yang, H.Q.; Chen, J.H. Current strategies and future prospects for enhancing microbial production of citric acid. Appl. Microbiol. Biotechnol. 2019, 103, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Karaffa, L.; Kubicek, C.P. Citric acid and itaconic acid accumulation: Variations of the same story? Appl. Microbiol. Biotechnol. 2019, 103, 2889–2902. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C. Citric acid from Aspergillus niger: A comprehensive overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.Y.; Zheng, X.M.; Tong, Y.; Shi, Y.C.; Sun, J.B. Systems metabolic engineering for citric acid production by Aspergillus niger in the post-genomic era. Microb. Cell Fact. 2019, 18, 28. [Google Scholar] [CrossRef]
- Nakamura, E.; Kadooka, C.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. Citrate exporter enhances both extracellular and intracellular citric acid accumulation in the koji fungi Aspergillus luchuensis mut. kawachii and Aspergillus oryzae. J. Biosci. Bioeng. 2021, 131, 68–76. [Google Scholar] [CrossRef]
- Steiger, M.G.; Rassinger, A.; Mattanovich, D.; Sauer, M. Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger. Metab. Eng. 2019, 52, 224–231. [Google Scholar] [CrossRef]
- Hou, L.; Liu, L.; Zhang, H.; Zhang, L.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Functional analysis of the mitochondrial alternative oxidase gene (aox1) from Aspergillus niger CGMCC 10142 and its effects on citric acid production. Appl. Microbiol. Biotechnol. 2018, 102, 7981–7995. [Google Scholar] [CrossRef]
- Ruijter, G.J.; Panneman, H.; Xu, D.; Visser, J. Properties of Aspergillus niger citrate synthase and effects of citA overexpression on citric acid production. FEMS Microbiol. Lett. 2000, 184, 35–40. [Google Scholar] [CrossRef]
- de Jongh, W.A.; Nielsen, J. Enhanced citrate production through gene insertion in Aspergillus niger. Metab. Eng. 2008, 10, 87–96. [Google Scholar] [CrossRef]
- Meijer, S.; Nielsen, M.L.; Olsson, L.; Nielsen, J. Gene deletion of cytosolic ATP: Citrate lyase leads to altered organic acid production in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; He, X.H.; Geng, H.R.; Liu, H. Physiological characterization of ATP-citrate lyase in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2014, 41, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.H.; Li, A.; Brickwedde, A.; Wilms, L.; Caspers, M.; Overkamp, K.; Punt, P.J. Rewiring a secondary metabolite pathway towards itaconic acid production in Aspergillus niger. Microb. Cell Fact. 2016, 15, 130. [Google Scholar] [CrossRef] [PubMed]
- Vesth, T.C.; Nybo, J.L.; Theobald, S.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F.; Hoof, J.B.; Brandl, J.; Salamov, A.; Riley, R.; et al. Investigation of inter- and intraspecies variation through genome sequencing of Aspergillus section Nigri. Nat. Genet. 2018, 50, 1688–1695. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, G.; Andersen, M.R.; Grotkjaer, T.; Regueira, T.B.; Hofmann, G.; Nielsen, J.; Olsson, L. Systems analysis unfolds the relationship between the phosphoketolase pathway and growth in Aspergillus nidulans. PLoS ONE 2008, 3, e3847. [Google Scholar] [CrossRef]
- Xu, Y.; Shan, L.; Zhou, Y.; Xie, Z.; Ball, A.S.; Cao, W.; Liu, H. Development of a Cre-loxP-based genetic system in Aspergillus niger ATCC 1015 and its application to construction of efficient organic acid-producing cell factories. Appl. Microbiol. Biotechnol. 2019, 103, 8105–8114. [Google Scholar] [CrossRef]
- Cao, W.; Yan, L.; Li, M.; Liu, X.; Xu, Y.; Xie, Z.; Liu, H. Identification and engineering a C4-dicarboxylate transporter for improvement of malic acid production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2020, 104, 9773–9783. [Google Scholar] [CrossRef]
- Cao, W.; Zhang, L.; Wu, L.; Zhang, M.; Liu, J.; Xie, Z.; Liu, H. Identification and genetic characterization of mitochondrial citrate transporters in Aspergillus niger. Front. Microbiol. 2022, 13, 1009491. [Google Scholar] [CrossRef]
- Hu, R.; Lin, L.; Liu, T.; Ouyang, P.; He, B.; Liu, S. Reducing sugar content in hemicellulose hydrolysate by DNS method: A revisit. J. Biobased Mater. Bio. 2008, 2, 156–161. [Google Scholar] [CrossRef]
- Xue, X.; Bi, F.; Liu, B.; Li, J.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Improving citric acid production of an industrial Aspergillus niger CGMCC 10142: Identification and overexpression of a high-affinity glucose transporter with different promoters. Microb. Cell Fact. 2021, 20, 168. [Google Scholar] [CrossRef]
- Bergman, A.; Siewers, V.; Nielsen, J.; Chen, Y. Functional expression and evaluation of heterologous phosphoketolases in Saccharomyces cerevisiae. AMB Expr. 2016, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Kamineni, A.; Consiglio, A.L.; MacEwen, K.; Chen, S.; Chifamba, G.; Shaw, A.J.; Tsakraklides, V. Increasing lipid yield in Yarrowia lipolytica through phosphoketolase and phosphotransacetylase expression in a phosphofructokinase deletion strain. Biotechnol. Biofuels 2021, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, J.; Yang, J.; Jiang, Y.; Yang, S. Strategies for optimizing acetyl-CoA formation from glucose in bacteria. Trends Biotechnol. 2022, 40, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Bogorad, I.W.; Lin, T.S.; Liao, J.C. Synthetic non-oxidative glycolysis enables complete carbon conservation. Nature 2013, 502, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.L.; Flamm, C.; Merkle, D.; Stadler, P.F. Chemical transformation motifs-modelling pathways as integer hyperflows. IEEE/ACM Trans. Comput. Biol. Bioinform. 2019, 16, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Krüsemann, J.L.; Lindner, S.N.; Dempfle, M.; Widmer, J.; Arrivault, S.; Debacker, M.; He, H.; Kubis, A.; Chayot, R.; Anissimova, M.; et al. Artificial pathway emergence in central metabolism from three recursive phosphoketolase reactions. FEBS J. 2018, 285, 4367–4377. [Google Scholar] [CrossRef]
- Hellgren, J.; Godina, A.; Nielsen, J.; Siewers, V. Promiscuous phosphoketolase and metabolic rewiring enables novel non-oxidative glycolysis in yeast for high-yield production of acetyl-CoA derived products. Metab. Eng. 2020, 62, 150–160. [Google Scholar] [CrossRef]
- Pel, H.J.; de Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype/Description | Source | |
|---|---|---|
| Strains | ||
| S469 | Tet-On::cre, hygs | Our laboratory [16] |
| S687 | Tet-On::cre, Δacs, hygr | This study |
| S688 | Tet-On::cre, Δacs, hygs | This study |
| S916 | Tet-On::cre, Δack, hygr | This study |
| S917 | Tet-On::cre, Δack, hygs | This study |
| S2738 | Tet-On::cre, ΔpkA, hygr | This study |
| S2740 | Tet-On::cre, ΔpkB, hygr | This study |
| S2908 | Tet-On::cre, ΔpkB, hygs | This study |
| S3008 | Tet-On::cre, ΔpkB, ΔpkA, hygr | This study |
| S3050 | Tet-On::cre, ΔpkB, ΔpkA, hygs | This study |
| S1422 | Tet-On::cre, Δacs, amyA::PgpdA::acs::TtrpC, hygr | This study |
| S1724 | Tet-On::cre, Δack, amyA::PgpdA::ack::TtrpC, hygr | This study |
| S3081 | Tet-On::cre, ΔpkB, ΔpkA, amyA::PgpdA::Bl-pk::TtrpC, hygr | This study |
| S3087 | Tet-On::cre, ΔpkB, ΔpkA, amyA::PgpdA::Ba-pk::TtrpC, hygr | This study |
| S3092 | Tet-On::cre, ΔpkB, ΔpkA, amyA::PgpdA::Ca-pk::TtrpC, hygr | This study |
| S3098 | Tet-On::cre, ΔpkB, ΔpkA, amyA::PgpdA::pkA::TtrpC, hygr | This study |
| S3104 | Tet-On::cre, ΔpkB, ΔpkA, amyA::PgpdA::pkB::TtrpC, hygr | This study |
| S2909 | Tet-On::cre, Δack, amyA::PgpdA::Ts-pta::TtrpC, hygr | This study |
| S2914 | Tet-On::cre, Δack, amyA::PgpdA::Bs-pta::TtrpC, hygr | This study |
| S3158 | Tet-On::cre, PgpdA::Ca-pk::TtrpC, PgpdA::Ts-pta::TtrpC, hygr | This study |
| Plasmids | ||
| pLH454 | loxP-hph-loxP, hygr, kanr, PgpdA, TtrpC | Our laboratory [16] |
| pLH594 | loxP-hph-loxP, hygr, pptr, kanr | Our laboratory [16] |
| pLH924 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA | Our laboratory [17] |
| pLH525 | loxP-hph-loxP, hygr, pptr, kanr, Δacs | This study |
| pLH670 | loxP-hph-loxP, hygr, pptr, kanr, Δack | This study |
| pLH1549 | loxP-hph-loxP, hygr, pptr, kanr, ΔpkA | This study |
| pLH1551 | loxP-hph-loxP, hygr, pptr, kanr, ΔpkB | This study |
| pLH821 | loxP-hph-loxP, hygr, kanr, PgpdA::acs::TtrpC | This study |
| pLH834 | loxP-hph-loxP, hygr, kanr, PgpdA::ack::TtrpC | This study |
| pLH1587 | loxP-hph-loxP, hygr, kanr, PgpdA::Bl-pk::TtrpC | This study |
| pLH1588 | loxP-hph-loxP, hygr, kanr, PgpdA::Ba-pk::TtrpC | This study |
| pLH1610 | loxP-hph-loxP, hygr, kanr, PgpdA::Ca-pk::TtrpC | This study |
| pLH1628 | loxP-hph-loxP, hygr, kanr, PgpdA::pkA::TtrpC | This study |
| pLH1630 | loxP-hph-loxP, hygr, kanr, PgpdA::pkB::TtrpC | This study |
| pLH1589 | loxP-hph-loxP, hygr, kanr, PgpdA::Ts-pta::TtrpC | This study |
| pLH1590 | loxP-hph-loxP, hygr, kanr, PgpdA::Bs-pta::TtrpC | This study |
| pLH822 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::acs::TtrpC | This study |
| pLH835 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::ack::TtrpC | This study |
| pLH1591 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::Bl-pk::TtrpC | This study |
| pLH1592 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::Ba-pk::TtrpC | This study |
| pLH1611 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::Ca-pk::TtrpC | This study |
| pLH1629 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::pkA::TtrpC | This study |
| pLH1631 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::pkB::TtrpC | This study |
| pLH1593 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::Ts-pta::TtrpC | This study |
| pLH1594 | loxP-hph-loxP, hygr, pptr, kanr, ΔamyA, PgpdA::Bs-pta::TtrpC | This study |
| pLH1080 | loxP-hph-loxP, hygr, kanr, PgdhA, TtrpC | This study |
| pLH1804 | loxP-hph-loxP, hygr, kanr, PgdhA::Ca-pk::TtrpC | This study |
| pLH1805 | loxP-hph-loxP, hygr, pptr, kanr, PgdhA::Ca-pk::TtrpC, PgpdA::Ts-pta::TtrpC | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, S.; Li, W.; Wang, G.; Xie, Z.; Cao, W.; Gao, W.; Liu, H. Engineering a Phosphoketolase Pathway to Supplement Cytosolic Acetyl-CoA in Aspergillus niger Enables a Significant Increase in Citric Acid Production. J. Fungi 2023, 9, 504. https://doi.org/10.3390/jof9050504
Liu J, Zhang S, Li W, Wang G, Xie Z, Cao W, Gao W, Liu H. Engineering a Phosphoketolase Pathway to Supplement Cytosolic Acetyl-CoA in Aspergillus niger Enables a Significant Increase in Citric Acid Production. Journal of Fungi. 2023; 9(5):504. https://doi.org/10.3390/jof9050504
Chicago/Turabian StyleLiu, Jiao, Shanshan Zhang, Wenhao Li, Guanyi Wang, Zhoujie Xie, Wei Cao, Weixia Gao, and Hao Liu. 2023. "Engineering a Phosphoketolase Pathway to Supplement Cytosolic Acetyl-CoA in Aspergillus niger Enables a Significant Increase in Citric Acid Production" Journal of Fungi 9, no. 5: 504. https://doi.org/10.3390/jof9050504