
Arthrobotrys blastospora sp. nov. (Orbiliomycetes): A Living Fossil Displaying Morphological Traits of Mesozoic Carnivorous Fungi

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Fungal Isolation
2.3. Morphological Observation
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analysis
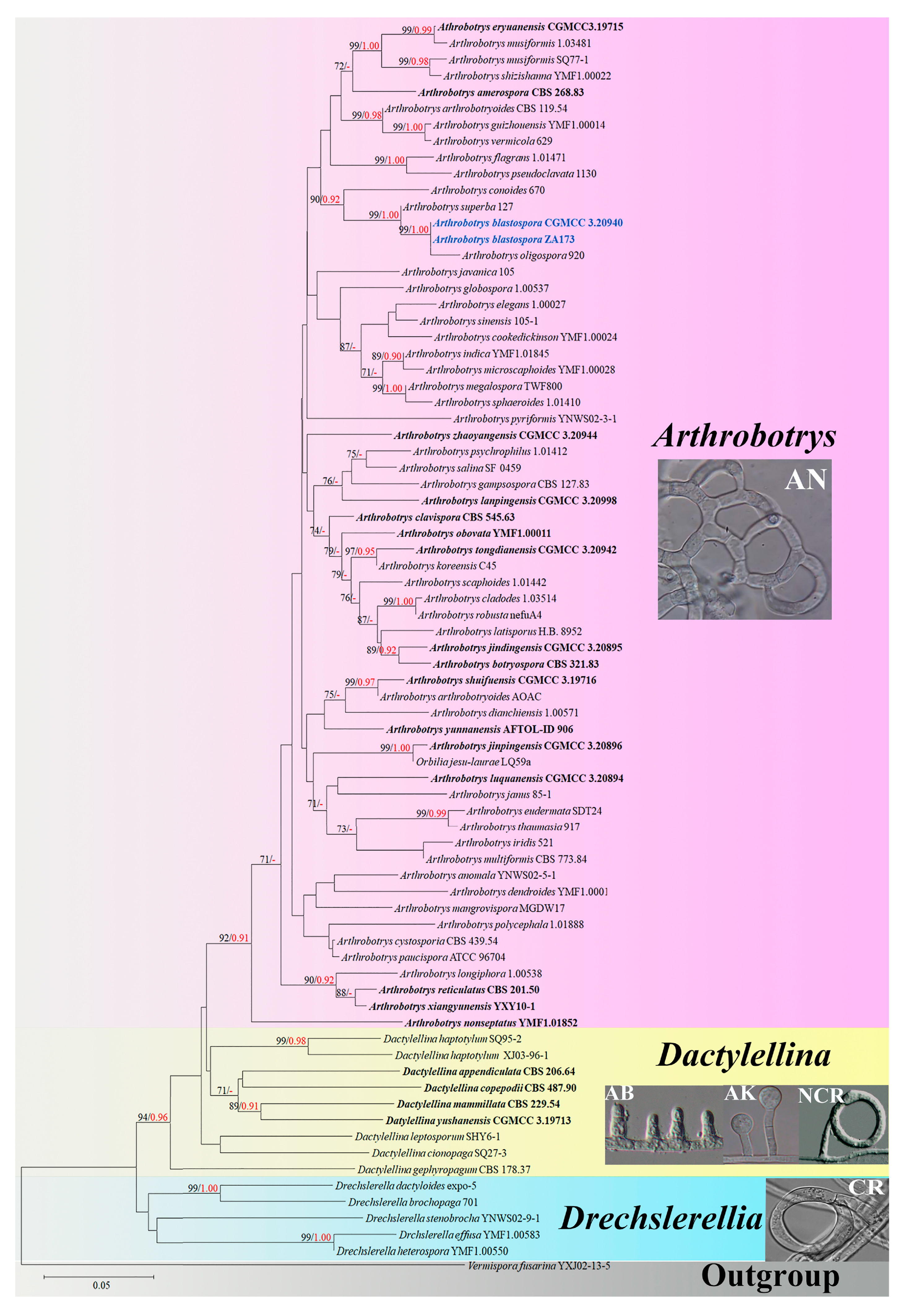
3. Results
3.1. Phylogenetic Analysis
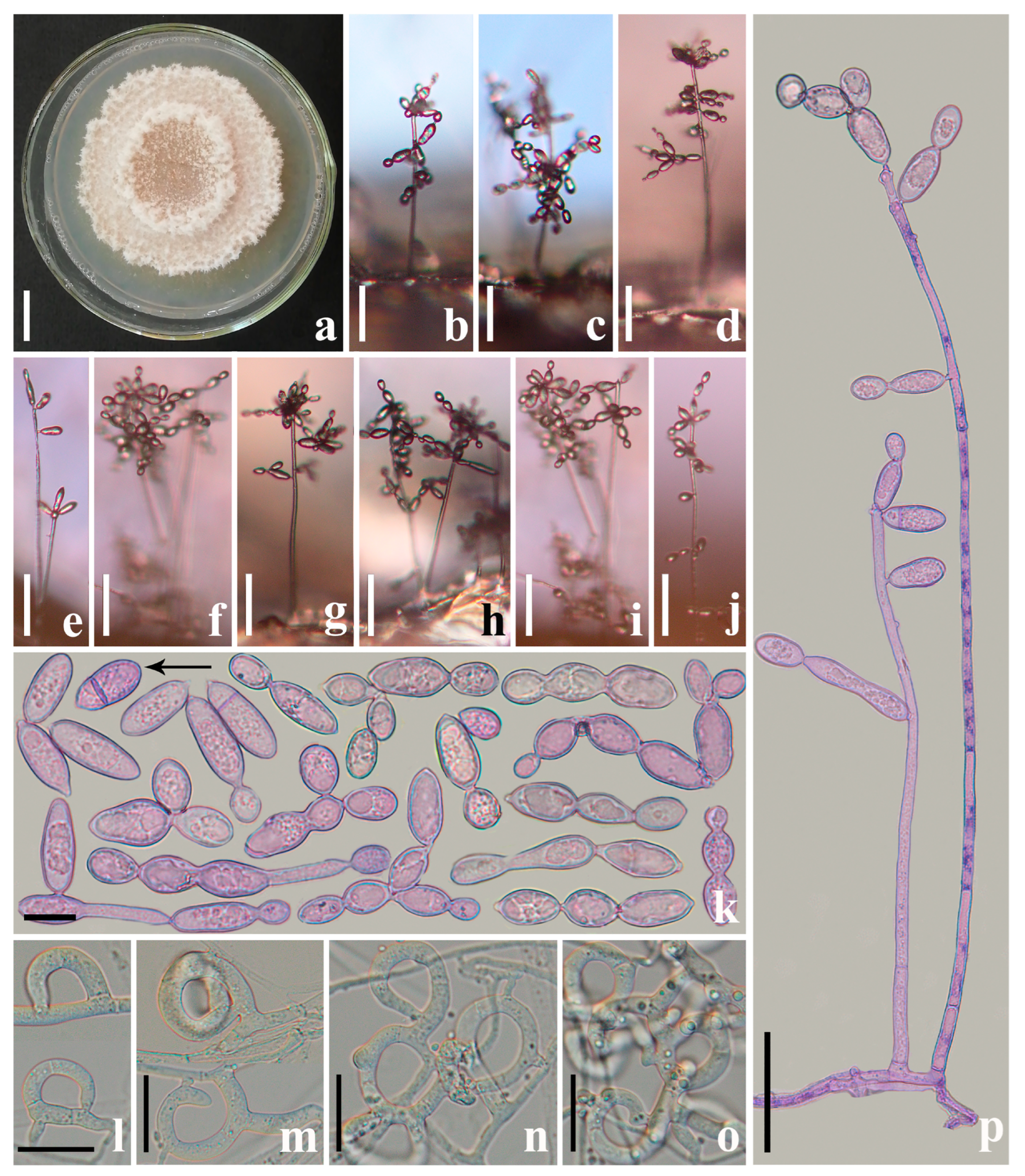
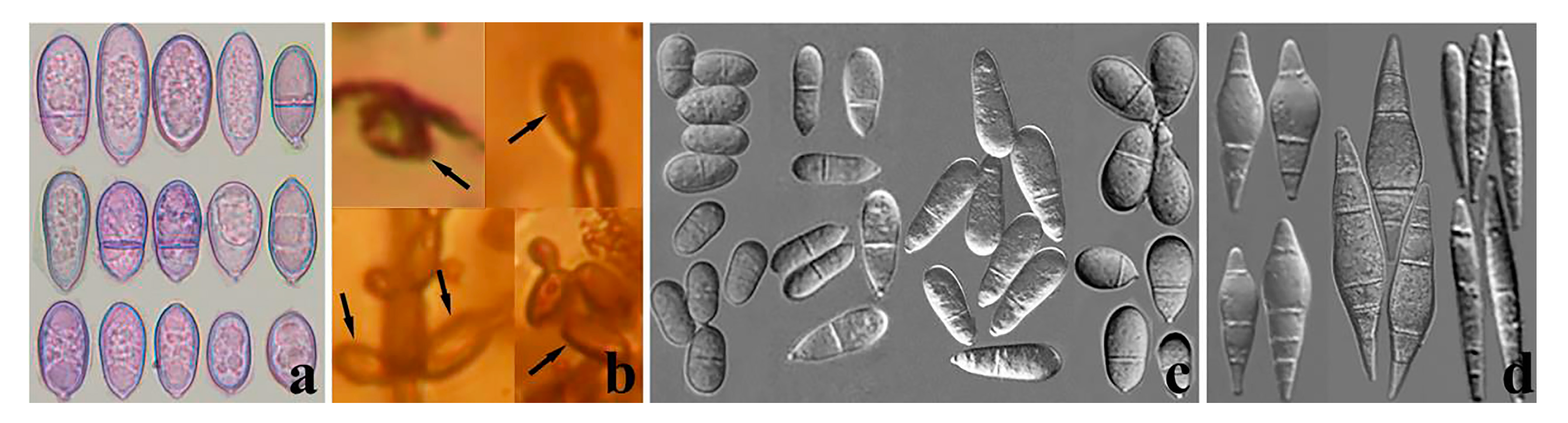
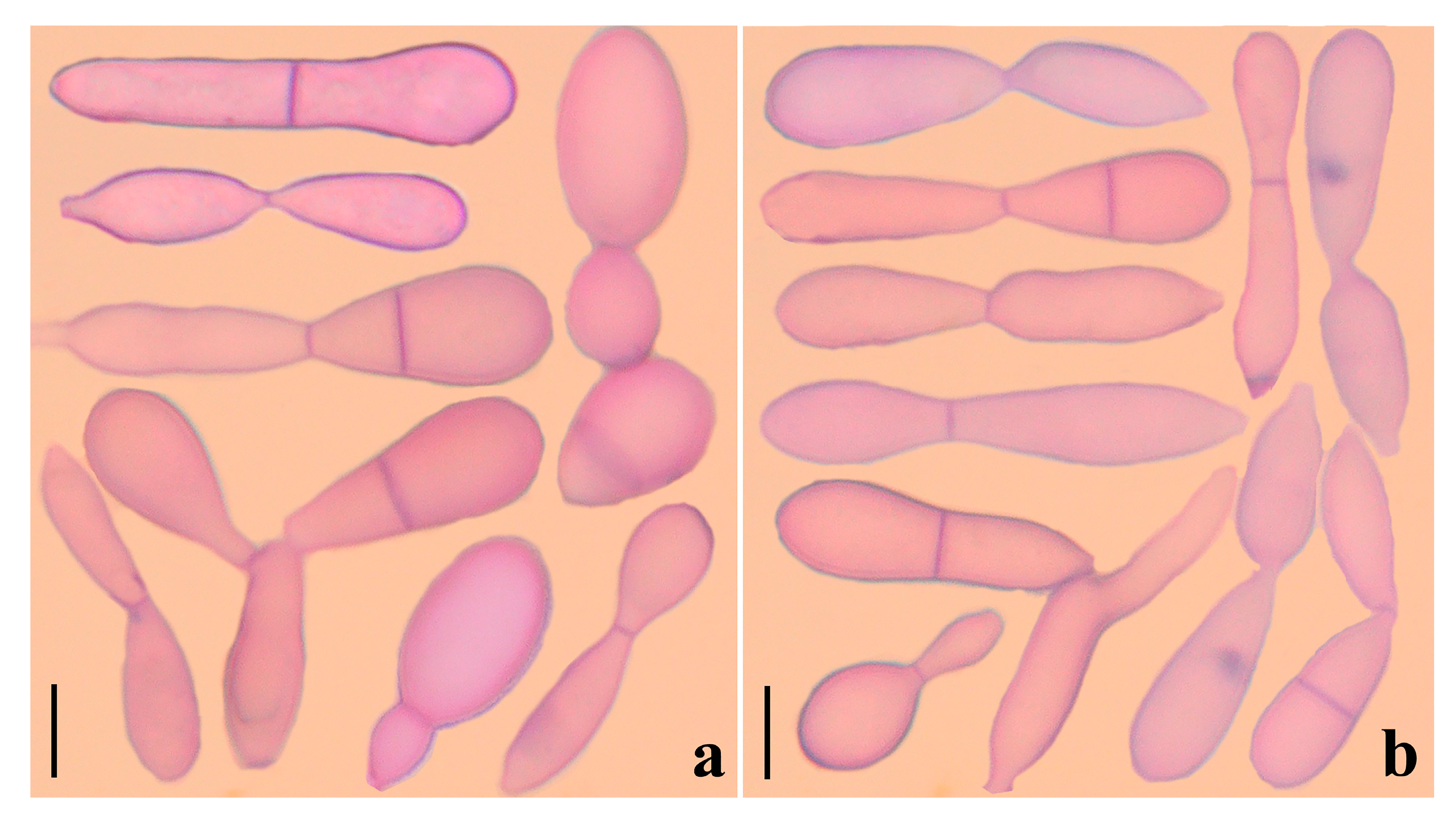
3.2. Taxonomy
4. Discussion
4.1. New Species of Arthrobotrys
4.2. Palaeoanellus Dimorphus Is an Ancient Ancestor of Modern Orbiliomycetes Carnivorous Fungi
4.3. The Possible Ancestral Position of Palaeoanellus Dimorphus
4.4. The Necessity of Strengthening the Research on Carnivorous Fungi in the Three Parallel Rivers
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, D.; Norman, C. What Don’t We Know? Science 2005, 309, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, F.H.; Lilje, O. Structure and function of fungal zoospores: Ecological implications. Fungal Ecol. 2009, 2, 53–59. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Wyngaert, S.V.D.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Genet. 2019, 17, 339–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, G.-J.; Solé, M.; Krauss, G.; Schlosser, D.; Wesenberg, D.; Bärlocher, F. Fungi in freshwaters: Ecology, physiology and biochemical potential. FEMS Microbiol. Rev. 2011, 35, 620–651. [Google Scholar] [CrossRef]
- Barron, G.L. Predators and parasites of microscopic animals. In Biology of Conidial Fungi; Cole, G.T., Kendrich, B., Eds.; ACADEMIC Press: New York, NY, USA, 2012; Volume 2, pp. 167–200. [Google Scholar]
- Barron, G.L. A new predatory hyphomycete capturing copepods. Can. J. Bot. 1990, 68, 691–696. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Gao, R.-H.; Zhang, K.-Q.; Cao, L. Dactylella tenuifusaria sp. nov., a rhizopod-capturing hyphomycete. Mycol. Res. 1996, 100, 236–238. [Google Scholar] [CrossRef]
- Liu, X.; Xiang, M.; Che, Y. The living strategy of nematophagous fungi. Mycoscience 2009, 50, 20–25. [Google Scholar] [CrossRef]
- Yang, E.; Xu, L.; Yang, Y.; Zhang, X.; Xiang, M.; Wang, C.; An, Z.; Liu, X. Origin and evolution of carnivorism in the Ascomycota (fungi). Proc. Natl. Acad. Sci. USA 2012, 109, 10960–10965. [Google Scholar] [CrossRef] [Green Version]
- Li, T.F.; Zhang, K.Q.; Liu, X.Z. Taxonomy of Nematophagous Fungi; Chinese Scientific and Technological Publications: Beijing, China, 2000. [Google Scholar]
- Ahrén, D.; Ursing, B.M.; Tunlid, A. Phylogeny of nematode-trapping fungi based on 18S rDNA sequences. FEMS Microbiol. Lett. 1998, 158, 179–184. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Hyde, K.D. Nematode-Trapping Fungi; Springer Science & Business: Berlin, Germany, 2014. [Google Scholar]
- Yang, Y.; Yang, E.; An, Z.; Liu, X.Z. Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hyde, K.D.; Jeewon, R.; Cai, L.; Vijaykrishna, D.; Zhang, K. Phylogenetics and evolution of nematode-trapping fungi (Orbiliales) estimated from nuclear and protein coding genes. Mycologia 2005, 97, 1034–1046. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Tzean, S.S. Phylogeny of the genus Arthrobotrys and allied nematode-trapping fungi based on rDNA sequences. Mycologia 1997, 89, 876–884. [Google Scholar] [CrossRef]
- Rubner, A. Revision of predacious hyphomycetes in the Dactylella-Monacrosporium complex. Stud. Mycol. 1996, 39, 1–134. [Google Scholar]
- Scholler, M.; Hagedorn, G.; Runner, A. A reevaluation of predatory orbiliaceous fungi. II. A new generic concept. Sydowia-Horn 1999, 51, 89–113. [Google Scholar]
- Flynn, L.J. Origin and evolution of the Diatomyidae, with clues to paleoecology from the fossil record. Bull. Carnegie Mus. Nat. Hist. 2007, 39, 173–181. [Google Scholar] [CrossRef]
- Gingerich, P.D.; Chaline, J. Origin and evolution of species: Evidence from the fossil record. In Modalités, Rythmes, Mécanismes de L’Évolution Biologique: Gradualisme Phylétique ou Équilibres Ponctués? Chaline, J., Ed.; Colloques Internationaux du Centre National de la Recherche Scientifique: Grand Rapids, MI, USA, 1983; pp. 125–130. [Google Scholar]
- Kukalova-Peck, J. Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record. J. Morphol. 1978, 156, 53–125. [Google Scholar] [CrossRef]
- Thaler, L. Origin and Evolution of Mice: An Appraisal of Fossil Evidence and Morphological Traits. Poxviruses 1986, 127, 3–11. [Google Scholar] [CrossRef]
- Jansson, H.-B.; Poinar, G.O. Some possible fossil nematophagous fungi. Trans. Br. Mycol. Soc. 1986, 87, 471–474. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Dorfelt, H.; Perrichot, V. Carnivorous Fungi from Cretaceous Amber. Science 2007, 318, 1743. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.R.; Dörfelt, H.; Perrichot, V. Palaeoanellus dimorphus gen. et sp. nov. (Deuteromycotina): A Cretaceous predatory fungus. Am. J. Bot. 2008, 95, 1328–1334. [Google Scholar] [CrossRef] [Green Version]
- Swe, A.; Li, J.; Zhang, K.Q.; Pointing, S.B.; Jeewon, R.; Hyde, K.D. Nematode-trapping fungi. Curr. Res. Environ. Appl. Mycol. 2011, 1, 1–26. [Google Scholar]
- Duddington, C. Notes on the technique of handling predacious fungi. Trans. Br. Mycol. Soc. 1955, 38, 97–103. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Swindell, S.R.; Plasterer, T.N. SEQMAN: Coting assembly. Meth. Mol. Biol. 1997, 70, 75–89. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Mo, M.H. Arthrobotrys et gengra cetera cognata. In Flora Fungorum Sinicorum; Science Press: Beijing, China, 2006; Volume 33. [Google Scholar]
- Zhang, F.; Boonmee, S.; Bhat, J.D.; Xiao, W.; Yang, X.-Y. New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China. J. Fungi 2022, 8, 671. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. Available online: http://mafft.cbrc.jp/alignment/server (accessed on 2 April 2022). [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Rezakhani, F.; Khodaparast, S.A.; Masigol, H.; Roja-Jimenez, K.; Grossart, H.-P.; Bakhshi, M. A preliminary report of aquatic hyphomycetes isolated from Anzali lagoon (Gilan province, North of Iran). Rostaniha 2019, 20, 123–143. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhu, H.; Zhang, K.-Q. Phylogenic analysis of adhesion related genes Mad1 revealed a positive selection for the evolution of trapping devices of nematode-trapping fungi. Sci. Rep. 2016, 6, 22609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, F.; Jiang, L.; Yang, Y.-Q.; Yang, X.-Y.; Xiao, W. Two new nematode-trapping fungi (Arthrobotrys, Orbiliaceae) from Yunnan, China. Phytotaxa 2022, 568, 255–266. [Google Scholar] [CrossRef]
- Wu, H.Y.; Kim, D.G.; Ryu, Y.H.; Zhou, X.B. Arthrobotrys koreensis, a new nematode-trapping species from Korea. Sydowia 2012, 64, 129–136. [Google Scholar]
- Swe, A.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Taxonomy and molecular phylogeny of Arthrobotrys mangrovispora, a new marine nematode-trapping fungal species. Bot. Mar. 2008, 51, 331–338. [Google Scholar] [CrossRef]
- Li, S.; Yu, Z.; Zhang, Y.; Qiao, M.; Guo, J.; Zhang, K. Arthrobotrys nonseptata, a new anamorph from an Orbilia species. Mycotaxon 2009, 109, 247–254. [Google Scholar] [CrossRef]
- Liu, S.; Su, H.; Su, X.; Zhang, F.; Liao, G.; Yang, X. Arthrobotrys xiangyunensis, a novel nematode-trapping taxon from a hot-spring in Yunnan Province, China. Phytotaxa 2014, 174, 89–96. [Google Scholar] [CrossRef]
- Mo, M.; Huang, X.; Zhou, W.; Huang, Y.; Hao, Y.E.; Zhang, K. Arthrobotrys yunnanensis sp. nov., the fourth anamorph of Orbilia auricolor. Fungal Divers. 2005, 18, 107–115. [Google Scholar]
- Zhang, F.; Zhou, X.-J.; Monkai, J.; Li, F.-T.; Liu, S.-R.; Yang, X.-Y.; Wen, X.; Hyde, K.D. Two new species of nematode-trapping fungi (Dactylellina, Orbiliaceae) from burned forest in Yunnan, China. Phytotaxa 2020, 452, 65–74. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.-O.; Beltrán-Tejera, E.; Pfister, D.H. Orbilia jesu-laurae (Ascomycota, Orbiliomycetes), a new species of neotropical nematode-trapping fungus from Puerto Rico, supported by morphology and molecular phylogenetics. Willdenowia 2020, 50, 241–251. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Phylogenetic Trees Made Easy: A How-to Manual; Sinauer Associates Sunderland: Sunderland, MA, USA, 2007. [Google Scholar]
- Rambaut, A. FigTree v1.3.1. 2010. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 6 June 2013).
- Thorn, R.G.; Scholler, M.; Gams, W. Mycological Research News. Mycol. Res. 2008, 112, 611–612. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Dörfelt, H.; Perrichot, V. Palaeomycology: Do we consider extinct lineages in the evaluation of fossil fungi? Mycol. Res. 2009, 113, 275–276. [Google Scholar]
- Nordbring-Hertz, B.; Jansson, H.; Tunlid, A. Nematophagous Fungi. eLS 2006, 1, 1–11. [Google Scholar] [CrossRef]
- Feng, B.; Zhao, Q.; Xu, J.; Qin, J.; Yang, Z.L. Drainage isolation and climate change-driven population expansion shape the genetic structures of Tuber indicum complex in the Hengduan Mountains region. Sci. Rep. 2016, 6, 21811. [Google Scholar] [CrossRef] [Green Version]
- Li, R.S.; Liu, W.; Chen, H. Three Parallel Rivers; China Translation and Publication Corporation: Shenzhen, China, 2001; pp. 44–67. [Google Scholar]
- Basnet, D.; Kandel, P.; Chettri, N.; Yang, Y.; Lodhi, M.S.; Htun, N.Z.; Uddin, K.; Sharma, E. Biodiversity Research Trends and Gaps from the Confluence of Three Global Biodiversity Hotspots in the Far-Eastern Himalaya. Int. J. Ecol. 2019, 2019, 1323419. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Wu, R.; Yang, F.; Wang, J.; Wu, W. Spatial trade-offs and synergies among ecosystem services within a global biodiversity hotspot. Ecol. Indic. 2018, 84, 371–381. [Google Scholar] [CrossRef]
- Li, J.W. A review of plant diversity and characteristics on three parallel rivers. Sci. Technol. Inf. 2009, 15, 59–64. [Google Scholar]
- Sun, H. Evolution of Arctic-Tertiary fora in Himalayan-Hengduan Mountains. Acta Bot. Yunnanica 2002, 24, 671–688. [Google Scholar]
- Sun, H.; Li, Z.M. Qinghai-Tibet Plateau uplift and its impact on Tethys fora. Adv. Earth Sci. 2003, 18, 852–862. [Google Scholar] [CrossRef]
- Wang, J.X.; Peng, X.F.; Yang, S.Y. Analysis of tourism resources and environmental vulnerability in three Parallel Rivers World Natural Heritage Site. J. Yunnan Norm. Univ. Nat. Sci. 2005, 25, 59–64. [Google Scholar]
- Zhang, D.C.; Zhang, Y.H.; Boufford, D.E.; Sun, H. Elevational patterns of species richness and endemism for some important taxa in the Hengduan Mountains, southwestern China. Biodivers. Conserv. 2009, 18, 699–716. [Google Scholar] [CrossRef]
- Fu, G.L. List of Rare and endangered Plants in China. Information System of Chinese Rare and Endangered Plant ISCREP. 1987. Available online: http://www.iplant.cn/rep/news/32 (accessed on 14 November 2022).
- Zhou, B. Revising the Yunnan Key Protected Wild Plants List. Acta Bot. Yunnanica 2010, 32, 221–226. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain Number | GenBank Accession Number | Reference | ||
|---|---|---|---|---|---|
| ITS | TEF | RPB2 | |||
| Arthrobotrys amerospora | CBS 268.83 | NR_159625 | — | — | [36] |
| Arthrobotrys anomala | YNWS02-5-1 | AY773451 | AY773393 | AY773422 | [13] |
| Arthrobotrys arthrobotryoides | CBS 119.54 | MH857262 | — | — | [36] |
| Arthrobotrys arthrobotryoides | AOAC | MF926580 | — | — | Unpublished |
| Arthrobotrys blastospora | CGMCC 3.20940 | OQ332405 | OQ341651 | OQ341649 | This study |
| Arthrobotrys blastospora | ZA173 | OM956088 | OQ341650 | OQ341648 | This study |
| Arthrobotrys botryospora | CBS 321.83 | NR_159626 | — | — | [36] |
| Arthrobotrys cladodes | 1.03514 | MH179793 | MH179616 | MH179893 | Unpublished |
| Arthrobotrys clavispora | CBS 545.63 | MH858353 | — | — | [36] |
| Arthrobotrys conoides | 670 | AY773455 | AY773397 | AY773426 | [13] |
| Arthrobotrys cookedickinson | YMF1.00024 | MF948393 | MF948550 | MF948474 | [12] |
| Arthrobotrys cystosporia | CBS 439.54 | MH857384 | — | — | [36] |
| Arthrobotrys dendroides | YMF1.00010 | MF948388 | MF948545 | MF948469 | [12] |
| Arthrobotrys dianchiensis | 1.00571 | MH179720 | — | MH179826 | [37] |
| Arthrobotrys elegans | 1.00027 | MH179688 | — | MH179797 | Unpublished |
| Arthrobotrys eryuanensis | CGMCC3.19715 | MT612105 | OM850307 | OM850301 | [32] |
| Arthrobotrys eudermata | SDT24 | AY773465 | AY773407 | AY773436 | [13] |
| Arthrobotrys flagrans | 1.01471 | MH179741 | MH179583 | MH179845 | Unpublished |
| Arthrobotrys gampsospora | CBS 127.83 | U51960 | — | — | [15] |
| Arthrobotrys globospora | 1.00537 | MH179706 | MH179562 | MH179814 | Unpublished |
| Arthrobotrys guizhouensis | YMF1.00014 | MF948390 | MF948547 | MF948471 | [12] |
| Arthrobotrys indica | YMF1.01845 | KT932086 | — | — | [38] |
| Arthrobotrys iridis | 521 | AY773452 | AY773394 | AY773423 | [13] |
| Arthrobotrys janus | 85-1 | AY773459 | AY773401 | AY773430 | [13] |
| Arthrobotrys javanica | 105 | EU977514 | — | — | Unpublished |
| Arthrobotrys jindingensis | CGMCC 3.20985 | OP236810 | OP272511 | OP272515 | [39] |
| Arthrobotrys jinpingensis | CGMCC 3.20896 | OM855569 | OM850311 | OM850305 | [32] |
| Arthrobotrys koreensis | C45 | JF304780 | — | — | [40] |
| Arthrobotrys lanpingensis | CGMCC3.20998 | OM855566 | OM850308 | OM850302 | [32] |
| Arthrobotrys latispora | H.B. 8952 | MK493125 | — | — | Unpublished |
| Arthrobotrys longiphora | 1.00538 | MH179707 | — | MH179815 | Unpublished |
| Arthrobotrys luquanensis | CGMCC3.20894 | OM855567 | OM850309 | OM850303 | [32] |
| Arthrobotrys mangrovispora | MGDW17 | EU573354 | — | — | [41] |
| Arthrobotrys megalospora | TWF800 | MN013995 | — | — | Unpublished |
| Arthrobotrys microscaphoides | YMF1.00028 | MF948395 | MF948552 | MF948476 | [12] |
| Arthrobotrys multiformis | CBS 773.84 | MH861834 | — | — | [36] |
| Arthrobotrys musiformis | SQ77-1 | AY773469 | AY773411 | AY773440 | [13] |
| Arthrobotrys musiformis | 1.03481 | MH179783 | MH179607 | MH179883 | Unpublished |
| Arthrobotrys nonseptata | YMF1.01852 | FJ185261 | — | — | [42] |
| Arthrobotrys obovata | YMF1.00011 | MF948389 | MF948546 | MF948470 | [12] |
| Arthrobotrys oligospora | 920 | AY773462 | AY773404 | AY773433 | [13] |
| Arthrobotrys paucispora | ATCC 96704 | EF445991 | — | — | [13] |
| Arthrobotrys polycephala | 1.01888 | MH179760 | MH179592 | MH179862 | Unpublished |
| Arthrobotrys pseudoclavata | 1130 | AY773446 | AY773388 | AY773417 | [13] |
| Arthrobotrys psychrophila | 1.01412 | MH179727 | MH179578 | MH179832 | Unpublished |
| Arthrobotrys pyriformis | YNWS02-3-1 | AY773450 | AY773392 | AY773421 | [13] |
| Arthrobotrys reticulata | CBS 201.50 | MH856589.1 | — | — | [36] |
| Arthrobotrys robusta | nefuA4 | MZ326655 | — | — | Unpublished |
| Arthrobotrys salina | SF 0459 | KP036623 | — | — | Unpublished |
| Arthrobotrys scaphoides | 1.01442 | MH179732 | MH179580 | MH179836 | Unpublished |
| Arthrobotrys shizishanna | YMF1.00022 | MF948392 | MF948549 | MF948473 | [12] |
| Arthrobotrys shuifuensis | CGMCC3.19716 | MT612334 | OM850306 | OM850300 | [32] |
| Arthrobotrys sinensis | 105-1 | AY773445 | AY773387 | AY773416 | [13] |
| Arthrobotrys sphaeroides | 1.01410 | MH179726 | MH179577 | MH179831 | Unpublished |
| Arthrobotrys superba | 127 | EU977558 | — | — | Unpublished |
| Arthrobotrys thaumasia | 917 | AY773461 | AY773403 | AY773432 | [13] |
| Arthrobotrys tongdianensis | CGMCC 3.20942 | OP236809 | OP272509 | OP272513 | [39] |
| Arthrobotrys vermicola | 629 | AY773454 | AY773396 | AY773425 | [13] |
| Arthrobotrys xiangyunensis | YXY10-1 | MK537299 | — | — | [43] |
| Arthrobotrys yunnanensis | AFTOL-ID 906 | DQ491512 | — | — | [44] |
| Arthrobotrys zhaoyangensis | CGMCC3.20944 | OM855568 | OM850310 | OM850304 | [32] |
| Dactylellina appendiculata | CBS 206.64 | AF106531 | DQ358227 | DQ358229 | [36] |
| Datylellinacionopogum | SQ27-3 | AY773467 | AY773409 | AY773438 | [13] |
| Dactylellina copepodii | CBS 487.90 | U51964 | DQ999835 | DQ999816 | [13] |
| Datylellinagephyropagum | CBS178.37 | U51974 | DQ999847 | DQ999802 | [13] |
| Datylellina haptotylum | SQ95-2 | AY773470 | AY773412 | AY773441 | [13] |
| Datylellina haptotylum | XJ03-96-1 | DQ999827 | DQ999849 | DQ999804 | [13] |
| Dactylellina leptosporum | SHY6-1 | AY773466 | AY773408 | AY773437 | [13] |
| Dactylellina mammillata | CBS229.54 | AY902794 | DQ999843 | DQ999817 | [13] |
| Dactylellina yushanensis | CGMCC3.19713 | MK372061 | MN915113 | MN915112 | [45] |
| Drechslerella brochopaga | 701 | AY773456 | AY773398 | AY773427 | [13] |
| Drechslerella dactyloides | expo-5 | AY773463 | AY773405 | AY773434 | [13] |
| Drechslerella effusa | YMF1.00583 | MF948405 | MF948557 | MF948484 | Unpublished |
| Drechslerella heterospora | YMF1.00550 | MF948400 | MF948554 | MF948480 | Unpublished |
| Drechslerella stenobrocha | YNWS02-9-1 | AY773460 | AY773402 | AY773431 | [13] |
| Orbilia jesu-laurae | LQ59a | MN816816 | — | — | [46] |
| Vermispora fusarina | YXJ02-13-5 | AY773447 | AY773389 | AY773418 | [13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Boonmee, S.; Yang, Y.-Q.; Zhou, F.-P.; Xiao, W.; Yang, X.-Y. Arthrobotrys blastospora sp. nov. (Orbiliomycetes): A Living Fossil Displaying Morphological Traits of Mesozoic Carnivorous Fungi. J. Fungi 2023, 9, 451. https://doi.org/10.3390/jof9040451
Zhang F, Boonmee S, Yang Y-Q, Zhou F-P, Xiao W, Yang X-Y. Arthrobotrys blastospora sp. nov. (Orbiliomycetes): A Living Fossil Displaying Morphological Traits of Mesozoic Carnivorous Fungi. Journal of Fungi. 2023; 9(4):451. https://doi.org/10.3390/jof9040451
Chicago/Turabian StyleZhang, Fa, Saranyaphat Boonmee, Yao-Quan Yang, Fa-Ping Zhou, Wen Xiao, and Xiao-Yan Yang. 2023. "Arthrobotrys blastospora sp. nov. (Orbiliomycetes): A Living Fossil Displaying Morphological Traits of Mesozoic Carnivorous Fungi" Journal of Fungi 9, no. 4: 451. https://doi.org/10.3390/jof9040451