
The Growth and Conidiation of Purpureocillium lavendulum Are Co-Regulated by Nitrogen Sources and Histone H3K14 Acetylation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Whole Genome Sequencing of P. lavendulum
2.3. Construction of the Histone Mutant Strain
2.4. Western Blotting Analysis
2.5. Knockout of Plgcn5 in P. lavendulum
2.6. ChIP-Seq Analysis
3. Results
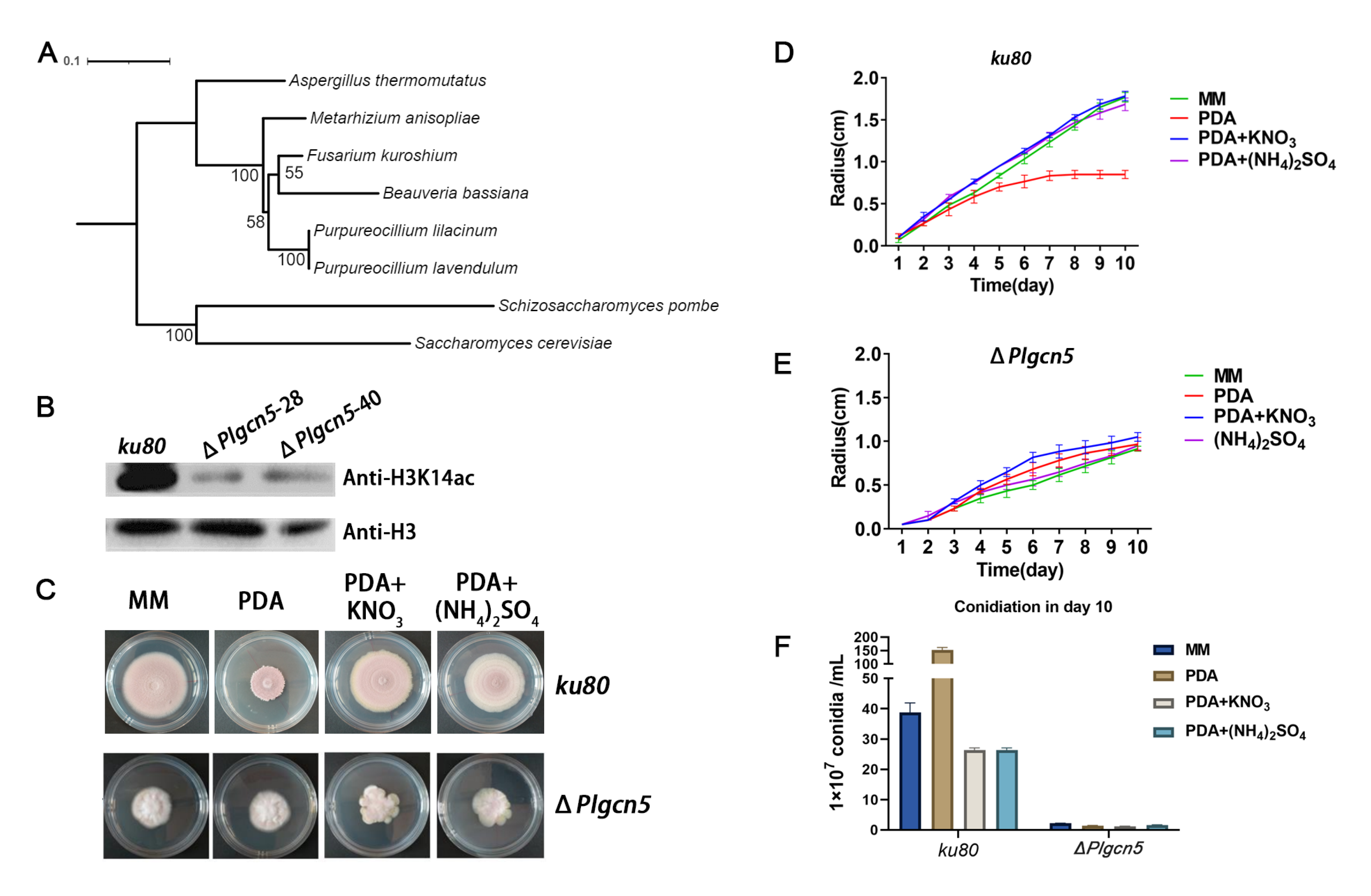
3.1. The Whole Genome Sequence of P. lavendulum
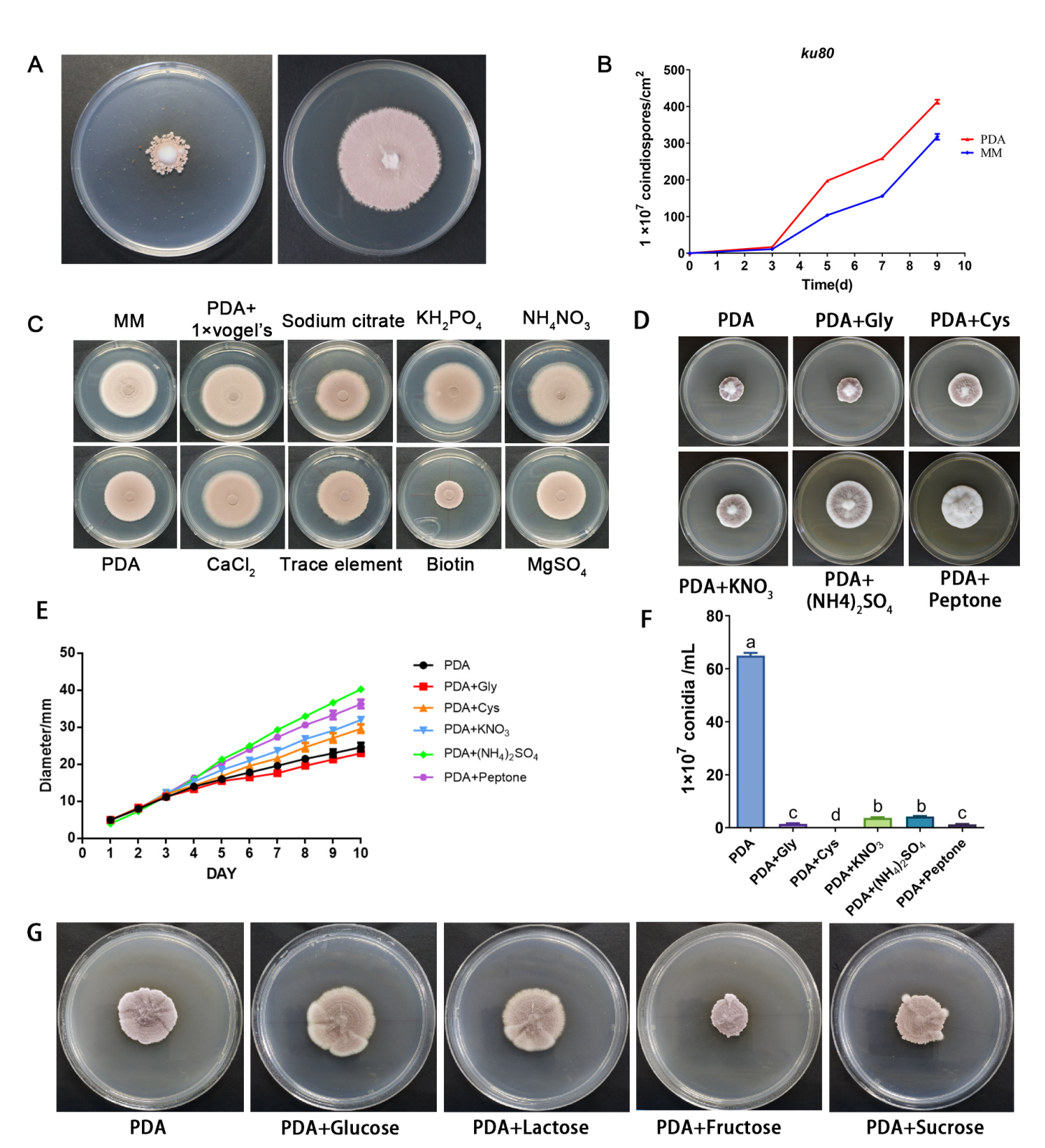
3.2. Effect of Nitrogen Sources on Vegetative Growth of P. lavendulum
3.3. The Modification of Histone H3K14 Affects the Growth and Conidiation Process of P. lavendulum
3.4. Knockout of Histone Acetyltransferase GCN5 Attenuated H3K14 Modification and Affected the Growth of P. lavendulum
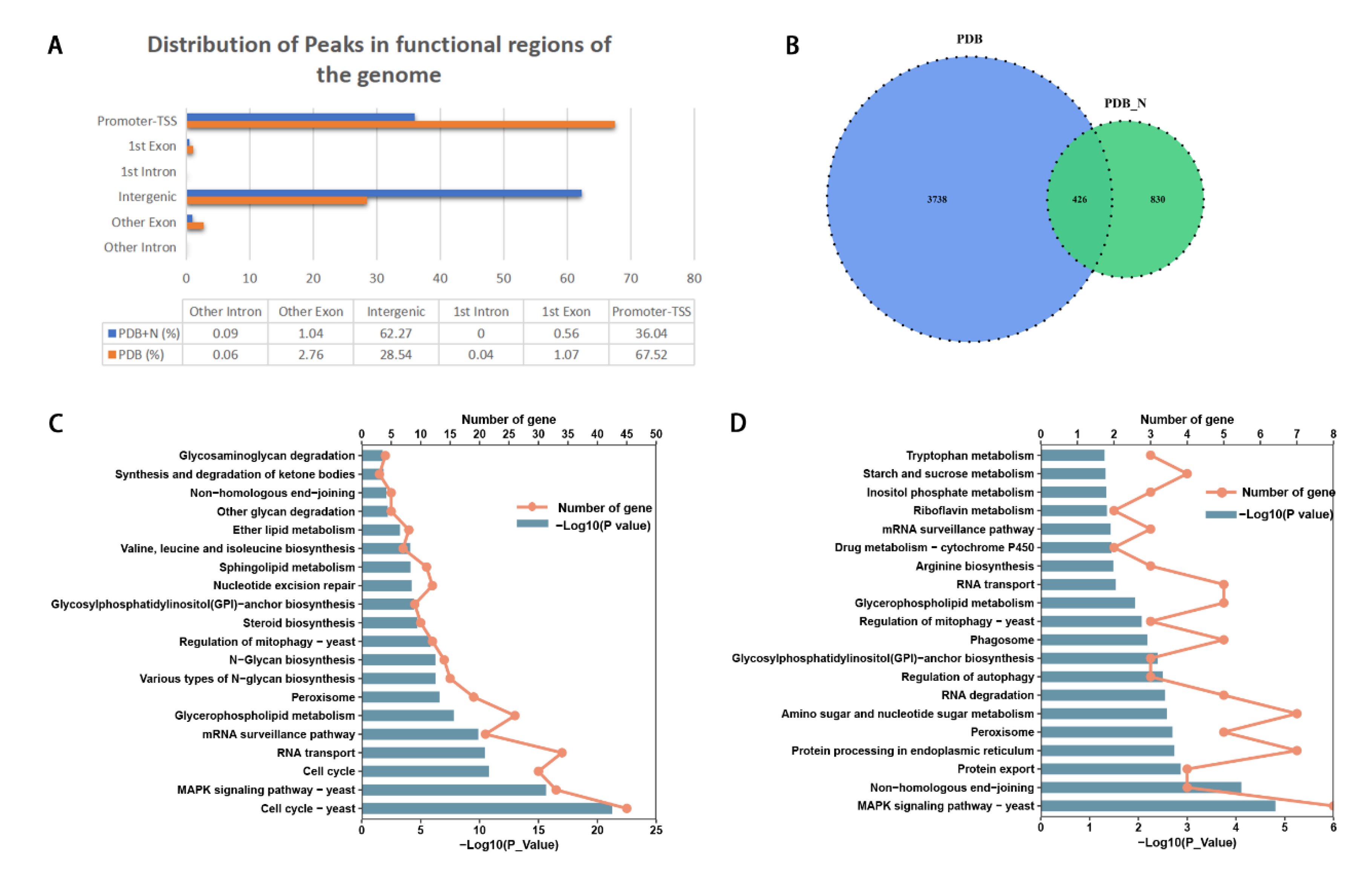
3.5. Histone H3K14 Acetylation Regulated Genes Involved in the Growth and Conidiation of P. lavendulum
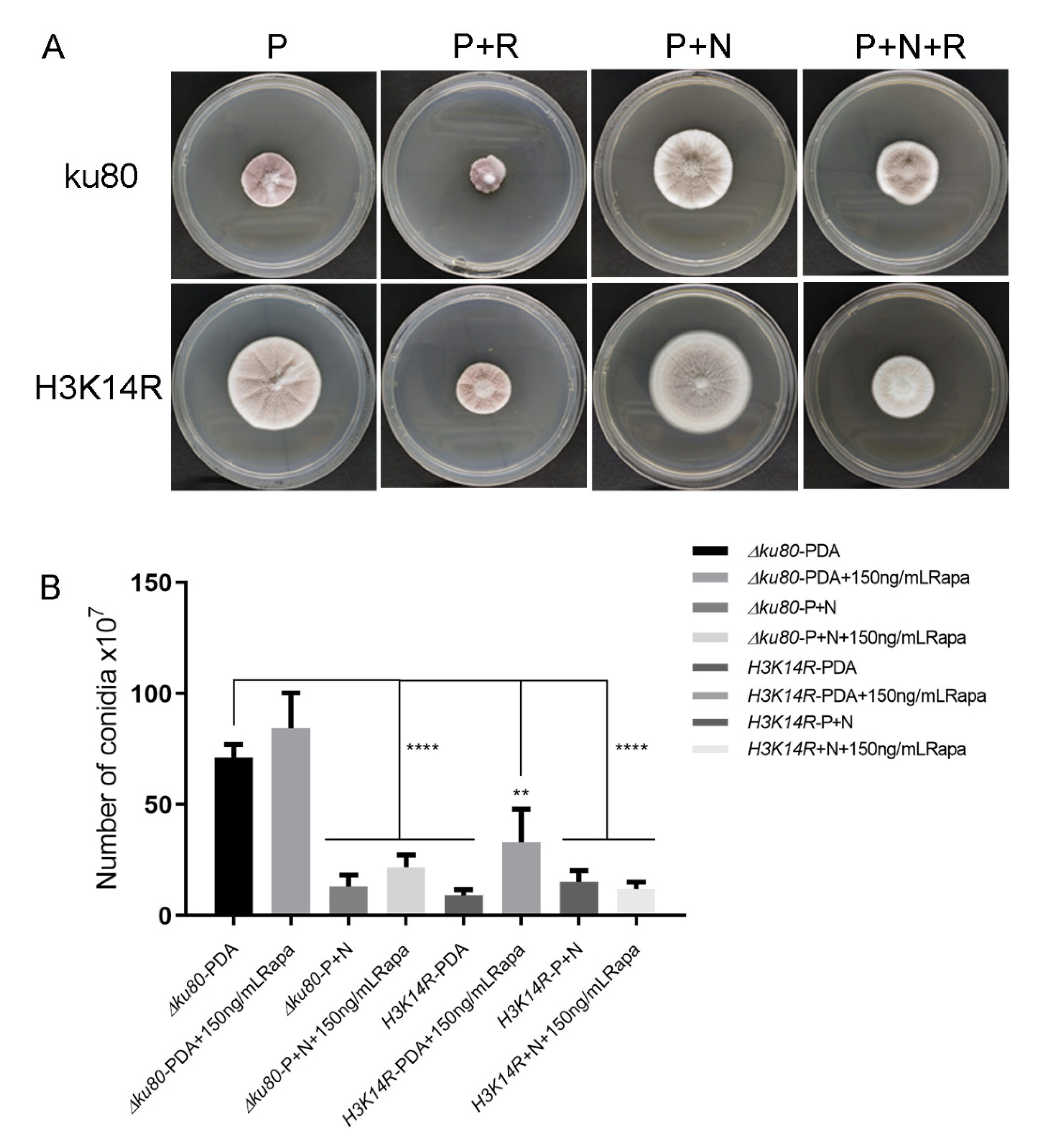
3.6. The TOR Signaling Pathway Is Not Directly Involved in the Nitrogen-H3K14ac Regulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivasubramaniam, N.; Hariharan, G.; Zakeel, M.C.M. Sustainable Management of Plant-Parasitic Nematodes: An Overview from Conventional Practices to Modern Techniques. In Management of Phytonematodes: Recent Advances and Future Challenges; Ansari, R.A., Rizvi, R., Mahmood, I., Eds.; Springer: Singapore, 2020; pp. 353–399. [Google Scholar]
- Xu, J. Assessing global fungal threats to humans. mLife 2022, 1, 223–240. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Li, H.; Wang, R.; Zhang, K.-Q.; Xu, J. Fungi–Nematode Interactions: Diversity, Ecology, and Biocontrol Prospects in Agriculture. J. Fungi 2020, 6, 206. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.S. Predatory Activity of Nematode Trapping Fungi against the Larvae of Trichostrongylus axei and Ostertagia ostertagi: A Possible Method of Biological Control. J. Helminthol. 1973, 47, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yang, H.-Y.; Cao, Y.-R.; Hui, Q.-Q.; Fan, H.-F.; Zhang, C.-C.; Han, J.-J.; Guo, Z.-Y.; Xu, J.; Zhang, K.-Q.; et al. Functional Characterization of Core Regulatory Genes Involved in Sporulation of the Nematophagous Fungus Purpureocillium lavendulum. mSphere 2020, 5, e00932-20. [Google Scholar] [CrossRef]
- Hsueh, Y.-P.; Mahanti, P.; Schroeder, F.C.; Sternberg, P.W. Nematode-Trapping Fungi Eavesdrop on Nematode Pheromones. Curr. Biol. 2013, 23, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Rubner, A. Revision of Predacious Hyphomycetes in the Dactylella-Monacrosporium Complex; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 1996. [Google Scholar]
- Etxebeste, O.; Garzia, A.; Espeso, E.A.; Ugalde, U. Aspergillus nidulans asexual development: Making the most of cellular modules. Trends Microbiol. 2010, 18, 569. [Google Scholar] [CrossRef] [Green Version]
- Fiałkowska, E.; Górska-Andrzejak, J.; Pajdak-Stós, A. The role of environmental factors in the conidiation of the predacious rotiferovorous fungus Zoophagus insidians (Zoopagomycota). Fungal. Ecol. 2023, 61, 101197. [Google Scholar] [CrossRef]
- Park, H.-S.; Yu, J.-H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41. [Google Scholar] [CrossRef]
- Keating, S.T.; El-Osta, A. Epigenetics and metabolism. Circ. Res. 2015, 116, 715–736. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, D.; Marcos, A.T.; Gacek, A.; Ramos, M.S.; Gutiérrez, G.; Reyesdomínguez, Y.; Strauss, J. The Histone Acetyltransferase GcnE (GCN5) Plays a Central Role in the Regulation of Aspergillus Asexual Development. Genetics 2014, 197, 1175. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-H.; Park, S.-H.; Kim, S.-S.; Lee, M.-W.; Yu, J.-H.; Shin, K.-S. Functional Characterization of the GNAT Family Histone Acetyltransferase Elp3 and GcnE in Aspergillus fumigatus. Int. J. Mol. Sci. 2023, 24, 2179. [Google Scholar] [CrossRef]
- Liu, L.; Cao, Y.-R.; Zhang, C.-C.; Fan, H.-F.; Guo, Z.-Y.; Yang, H.-Y.; Chen, M.; Han, J.-J.; Xu, J.; Zhang, K.-Q.; et al. An Efficient Gene Disruption System for the Nematophagous Fungus Purpureocillium lavendulum. Fungal Biol. 2019, 123, 274–282. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, X.; Qian, Y.; Chen, X.; Liu, R.; Zeng, G.; Zhao, H.; Fang, W. A high-throughput gene disruption methodology for the entomopathogenic fungus Metarhizium robertsii. PLoS ONE 2014, 9, e107657. [Google Scholar] [CrossRef]
- Lai, Y.; Cao, X.; Chen, J.; Wang, L.; Wei, G.; Wang, S. Coordinated regulation of infection-related morphogenesis by the KMT2-Cre1-Hyd4 regulatory pathway to facilitate fungal infection. Sci. Adv. 2020, 6, eaaz1659. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Wu, H.; Liu, Z.; Shen, R.; Gao, H.; Yang, J.; Zhang, K. Proteomic and transcriptional analyses of Arthrobotrys oligospora cell wall related proteins reveal complexity of fungal virulence against nematodes. Appl. Microbiol. Biotechnol. 2013, 97, 8683–8692. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic. Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Li, W.; Qi, S.; Gai, K.; Chen, Y.; Suo, J.; Cao, Y.; He, Y.; Wang, Y.; He, Q. The highly expressed methionine synthase gene of Neurospora crassa is positively regulated by its proximal heterochromatic region. Nucleic Acids Res. 2014, 42, 6183–6195. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.-M.; Zhang, Y.; Xu, J.; Zhang, K.-Q.; Cao, Y.-R. Characterization of the complete mitochondrial genome of the nematophagous fungus Purpureocillium lavendulum. Mitochondrial. DNA B 2021, 6, 33–35. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.R.; Staiger, C.J.; Hamer, J.E. Inactivation of the mitogen-activated protein kinase Mps1 from the rice blast fungus prevents penetration of host cells but allows activation of plant defense responses. Proc. Natl. Acad. Sci. USA. 1998, 95, 12713–12718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussink, H.J.; Osmani, S.A. A mitogen-activated protein kinase (MPKA) is involved in polarized growth in the filamentous fungus, Aspergillus nidulans. FEMS Microbiol. Lett. 1999, 173, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Campbell, M.; Murphy, J.; Lam, S.; Xu, J.R. The BMP1 gene is essential for pathogenicity in the gray mold fungus Botrytis cinerea. Mol. Plant. Microbe Interact. 2000, 13, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, Y.S.; Lee, D.G.; Idnurm, A.; Ianiri, G.; Bahn, Y.S. The TOR Pathway Plays Pleiotropic Roles in Growth and Stress Responses of the Fungal Pathogen Cryptococcus neoformans. Genetics 2019, 212, 1241–1258. [Google Scholar] [CrossRef]
- Martin, D.E.; Hall, M.N. The expanding TOR signaling network. Curr. Opin. Cell Biol. 2005, 17, 158–166. [Google Scholar] [CrossRef]
- Skromne, I.; Sánchez, O.; Aguirre, J. Starvation stress modulates the expression of the Aspergillus nidulans brlA regulatory gene. Microbiology 1995, 141, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Barron, G.L. Predatory fungi, wood decay, and the carbon cycle. Biodiversity 2003, 4, 3–9. [Google Scholar] [CrossRef]
- Müller, B.; Russo, V. Nitrogen starvation or glucose limitation induces conidiation in constantly shaken liquid cultures of Neurospora crassa. Fungal Genet. Rep. 1989, 36, 11. [Google Scholar] [CrossRef] [Green Version]
- Krasniewski, I.; Molimard, P.; Feron, G.; Vergoignan, C.; Durand, A.; Cavin, J.-F.; Cotton, P. Impact of solid medium composition on the conidiation in Penicillium camemberti. Process Biochem. 2006, 41, 1318–1324. [Google Scholar] [CrossRef]
- Pascual, S.; Melgarejo, P.; Magan, N. Induction of submerged conidiation of the biocontrol agent Penicillium oxalicum. Appl. Microbiol. Biotechnol. 1997, 48, 389–392. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Nützmann, H.W.; Reyes-Dominguez, Y.; Scherlach, K.; Schroeckh, V.; Horn, F.; Gacek, A.; Schümann, J.; Hertweck, C.; Strauss, J.; Brakhage, A.A. Bacteria-induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl. Acad. Sci. USA. 2011, 108, 14282–14287. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Wang, J.J.; Fu, B.; Ying, S.H.; Feng, M.G. Gcn5-dependent histone H3 acetylation and gene activity is required for the asexual development and virulence of Beauveria bassiana. Environ. Microbiol. 2018, 20, 1484–1497. [Google Scholar] [CrossRef]
- Dixon, K.P.; Xu, J.-R.; Smirnoff, N.; Talbot, N.J. Independent Signaling Pathways Regulate Cellular Turgor during Hyperosmotic Stress and Appressorium-Mediated Plant Infection by Magnaporthe grisea. Plant Cell 1999, 11, 2045–2058. [Google Scholar] [CrossRef] [Green Version]
- Takano, Y.; Kikuchi, T.; Kubo, Y.; Hamer, J.E.; Mise, K.; Furusawa, I. The Colletotrichum lagenarium MAP kinase gene CMK1 regulates diverse aspects of fungal pathogenesis. Mol. Plant. Microbe Interact. 2000, 13, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Guertin, D.A.; Guntur, K.V.; Bell, G.W.; Thoreen, C.C.; Sabatini, D.M. Functional genomics identifies TOR-regulated genes that control growth and division. Curr. Biol. 2006, 16, 958–970. [Google Scholar] [CrossRef] [Green Version]
- Rohde, J.R.; Cardenas, M.E. Nutrient Signaling through TOR Kinases Controls Gene Expression and Cellular Differentiation in Fungi. In TOR: Target of Rapamycin; Thomas, G., Sabatini, D.M., Hall, M.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, P.; Han, J.-J.; Zhang, C.-C.; Tang, P.-P.; Qi, F.-N.; Zhang, K.-Q.; Liang, L.-M. The Growth and Conidiation of Purpureocillium lavendulum Are Co-Regulated by Nitrogen Sources and Histone H3K14 Acetylation. J. Fungi 2023, 9, 325. https://doi.org/10.3390/jof9030325
Tang P, Han J-J, Zhang C-C, Tang P-P, Qi F-N, Zhang K-Q, Liang L-M. The Growth and Conidiation of Purpureocillium lavendulum Are Co-Regulated by Nitrogen Sources and Histone H3K14 Acetylation. Journal of Fungi. 2023; 9(3):325. https://doi.org/10.3390/jof9030325
Chicago/Turabian StyleTang, Ping, Jing-Jing Han, Chen-Chen Zhang, Ping-Ping Tang, Feng-Na Qi, Ke-Qin Zhang, and Lian-Ming Liang. 2023. "The Growth and Conidiation of Purpureocillium lavendulum Are Co-Regulated by Nitrogen Sources and Histone H3K14 Acetylation" Journal of Fungi 9, no. 3: 325. https://doi.org/10.3390/jof9030325