
A Correlation between 3′-UTR of OXA1 Gene and Yeast Mitochondrial Translation
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids and Media
2.2. DNA Manipulations
2.3. Mitochondria Isolation
2.4. qRT-PCR
2.5. β-Galactosidase Liquid Assay
2.6. Mass Spectrometry Analysis
3. Results and Discussion
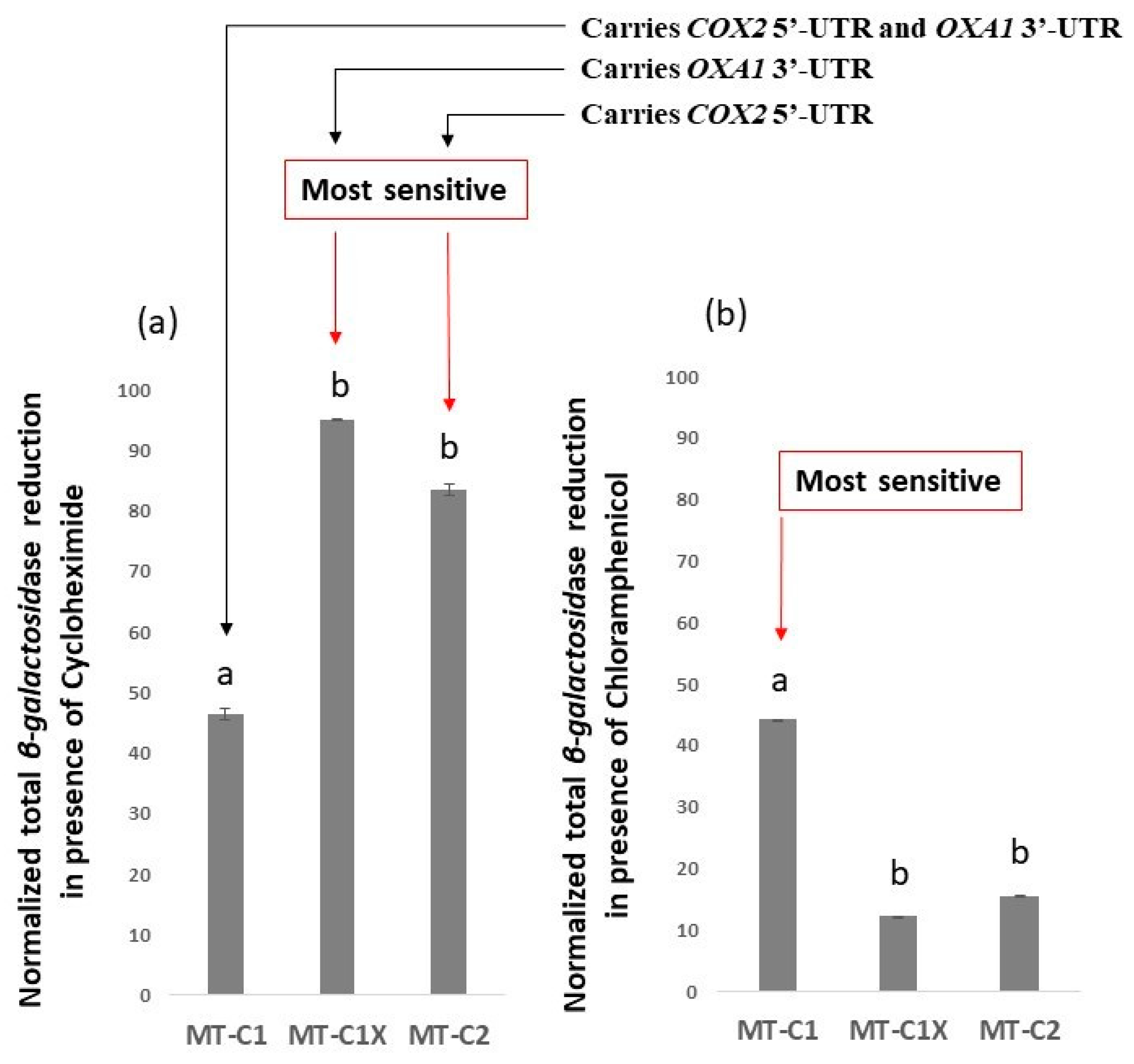
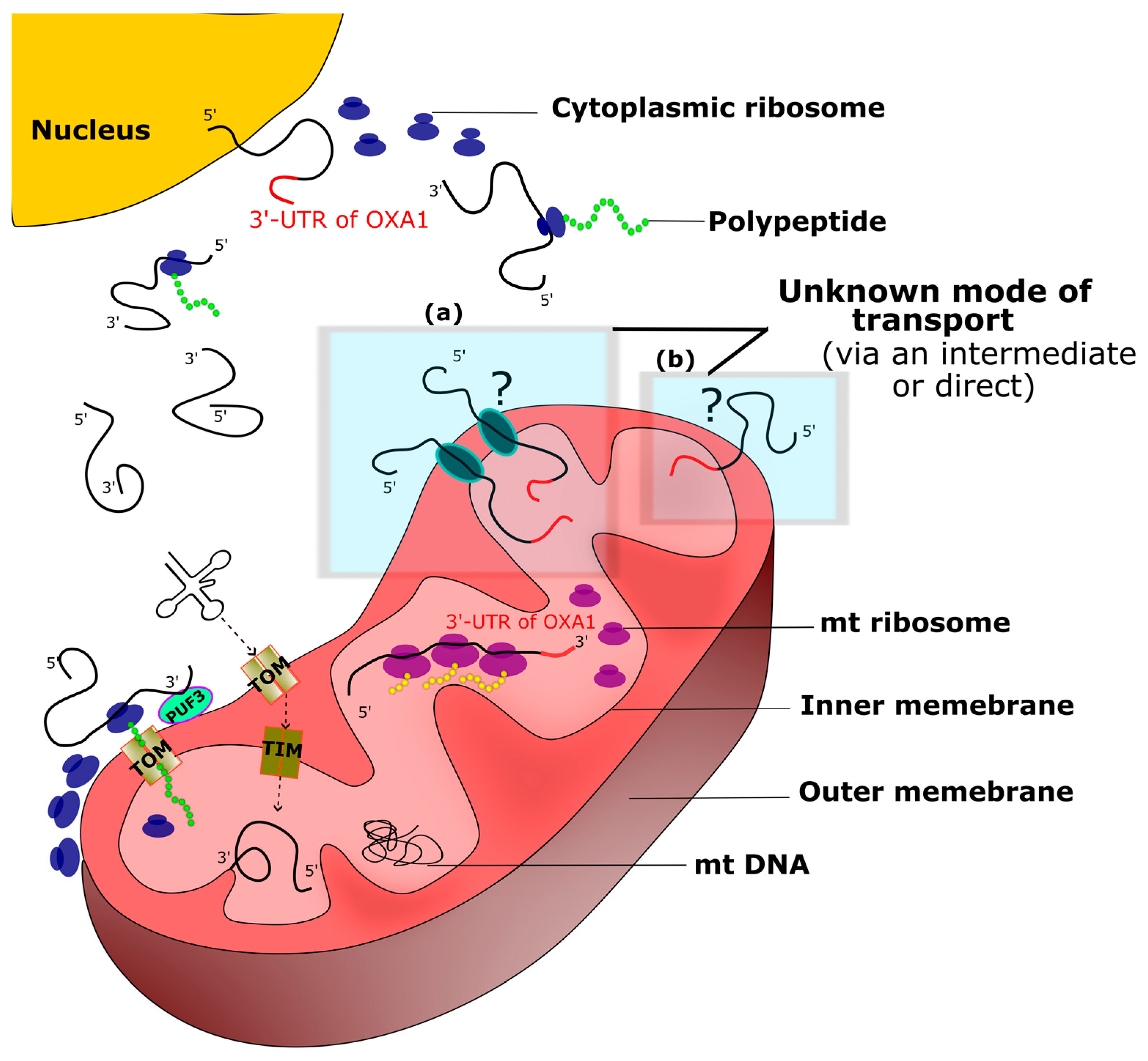
3.1. OXA1 Gene 3′-UTR Directs β-Galactosidase mRNA to the Vicinity of Mitochondria
3.2. Observed β-Galactosidase Activity Contains Signatures of Mitochondrial Translation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fox, T.D. Mitochondrial protein synthesis, import, and assembly. Genetics 2012, 192, 1203–1234. [Google Scholar] [CrossRef] [Green Version]
- Kummer, E.; Ban, N. Mechanisms and regulation of protein synthesis in mitochondria. Nat. Rev. Mol. Cell Biol. 2021, 22, 307–325. [Google Scholar] [CrossRef]
- Falkenberg, M.; Larsson, N.; Gustafsson, C. DNA Replication and Transcription in Mammalian Mitochondria. Annu. Rev. 2007, 76, 679–699. [Google Scholar] [CrossRef]
- Herrmann, J.M.; Woellhaf, M.W.; Bonnefoy, N. Control of protein synthesis in yeast mitochondria: The concept of translational activators. BBA-Mol. Cell Res. 2013, 1833, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Kepp, O.; Kroemer, G. Mitochondrial Regulation of Cell Death: A Phylogenetically Conserved Control. Micorbial Cell 2016, 3, 101–108. Available online: https://microbialcell.com/researcharticles/mitochondrial-regulation-of-cell-death-a-phylogenetically-conserved-control/ (accessed on 21 December 2019). [CrossRef] [Green Version]
- Schapira, A.H.V. Mitochondrial disease. Lancet 2006, 368, 70–82. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Chandel, N.S. Targeting mitochondria metabolism for cancer therapy. Nat. Chem. Biol. 2015, 11, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the Regulation of Innate and Adaptive Immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Shimada, E.; Zhang, J.; Hong, J.S.; Smith, G.M.; Teitell, M.A. Correcting human mitochondrial mutations with targeted RNA import. Proc. Natl. Acad. Sci. USA 2012, 109, 4840–4845. Available online: https://www.pnas.org/content/early/2012/03/07/1116792109 (accessed on 21 December 2019). [CrossRef] [PubMed] [Green Version]
- Brand, R.M.; Wipf, P.; Durham, A.; Epperly, M.W.; Greenberger, J.S.; Falo, L.D., Jr. Targeting Mitochondrial Oxidative Stress to Mitigate UV-Induced Skin Damage. Front. Pharmacol. 2018, 9, 920. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6110189/#!po=46.7742 (accessed on 21 December 2019). [CrossRef] [PubMed]
- Osman, C.; Noriega, T.R.; Okreglak, V.; Fung, J.C.; Walter, P. Integrity of the Yeast Mitochondrial Genome, But Not Its Distribution and Inheritance, Relies on Mitochondrial Fission and Fusion. Proc. Natl. Acad. Sci. USA 2015, 112, E947–E956. Available online: https://www.pnas.org/content/112/9/E947 (accessed on 21 December 2019). [CrossRef] [PubMed] [Green Version]
- Kren, B.T.; Wong, P.Y.; Sarver, A.; Zhang, X.; Zeng, Y.; Steer, C.J. microRNAs identified in highly purified liver-derived mitochondria may play a role in apoptosis. RNA Biol. 2009, 6, 65–72. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19106625 (accessed on 21 December 2019). [CrossRef] [PubMed] [Green Version]
- Schneider, A. Mitochondrial tRNA Import and Its Consequences for Mitochondrial Translation. Annu. Rev. Biochem. 2011, 80, 1033–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiyar, R.; Gagneur, J.; Steinmetz, L. Identification of mitochondrial disease genes through integrative analysis of multiple datasets. Methods Mol. Biol. 2008, 46, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Herrmann, J.M. Co-Translational Membrane Insertion of Mitochondrially Encoded Proteins. BBA-Mol. Cell Res. 2010, 1803, 767–775. Available online: https://www.sciencedirect.com/science/article/pii/S0167488909002924?via%3Dihub (accessed on 22 December 2019). [CrossRef] [PubMed] [Green Version]
- Gadir, N.; Haim-Vilmovsky, L.; Kraut-Cohen, J.; Gerst, J.E. Localization of mRNAs Coding for Mitochondrial Proteins in the Yeast Saccharomyces cerevisiae. Rna 2011, 17, 1551–1565. Available online: https://rnajournal.cshlp.org/content/17/8/1551.long (accessed on 21 December 2022). [CrossRef] [Green Version]
- Krogan, N.J.; Kim, M.; Tong, A.; Golshani, A.; Cagney, G.; Canadien, V.; Richards, D.P.; Beattie, B.K.; Emili, A.; Boone, C.; et al. Methylation of Histone H3 by Set2 in Saccharomyces cerevisiae Is Linked to Transcriptional Elongation by RNA Polymerase II. Mol. Cell. Biol. 2003, 23, 4207–4218. [Google Scholar] [CrossRef] [Green Version]
- Alamgir, M.; Jessulat, M.; Azizi, A.; Golshani, A. Chemical-genetic profile analysis of five inhibitory compounds in yeast. BMC Chem. Biol. 2010, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Weber, P.J.A.; Weber, G.; Eckerskorn, C. Isolation of Organelles and Prefractionation of Protein Extracts Using Free-Flow Electrophoresis. Curr. Protoc. Protein Sci. 2003, 32, 22.5.1–22.5.21. [Google Scholar] [CrossRef]
- Samanfar, B.; Omidi, K.; Hooshyar, M.; Laliberte, B.; Alamgir, M.; Seal, A.J.; Ahmed-Muhsin, E.; Viteri, D.F.; Said, K.; Chalabian, F.; et al. Large-scale investigation of oxygen response mutants in Saccharomyces cerevisiae. Mol. Biosyst. 2013, 9, 1351–1359. [Google Scholar] [CrossRef]
- Sylvestre, J.; Margeot, A.; Jacq, C.; Dujardin, G. The Role of the 3′ Untranslated Region in mRNA Sorting to the Vicinity of Mitochondria Is Conserved from Yeast to Human Cells. Mol. Biol. Cell 2003, 14, 3848–3856. Available online: ncbi.nlm.nih.gov/pubmed/12972568 (accessed on 21 December 2019). [CrossRef] [Green Version]
- Vardi-Oknin, D.; Arava, Y. Characterization of Factors Involved in Localized Translation Near Mitochondria by Ribosome-Proximity Labeling. Front. Cell Dev. Biol. 2019, 7, 305. [Google Scholar] [CrossRef] [Green Version]
- Laso, M.R.V.; Zhus, D.; Sagliocco, F.; Brown, A.J.P.; Mick, F.; Mccarthyl, J.E.G. Inhibition of Translational Initiation in the Yeast Saccharomyces Position of Hairpin cereuisiae as a Function of the Stability and Structures in the mRNA Leader. Biol. Chem. 1993, 268, 6453–6462. Available online: http://www.jbc.org/content/268/9/6453.abstract (accessed on 21 December 2019). [CrossRef]
- Dunstan, H.M.; Green-willms, N.S.; Fox, T.D. In Vivo analysis of Saccharomyces cerevisiae COX2 mRNA 5′-untraslated leader functions in Mitochondrial translation initiation and translational activation. Genet. Soc. Am. 1997, 147, 87–100. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1208125/ (accessed on 21 December 2019).
- Green-willms, N.S.; Butler, C.A.; Dunstan, H.M.; Fox, T.D. Pet111p, an Inner Membrane-Bound Translational Activator That Limits Expression of the Saccharomyces cerevisiae Mitochondrial Gene COX2. Biol. Chem. 2001, 276, 6392–6397. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11106667 (accessed on 21 December 2019). [CrossRef] [PubMed] [Green Version]
- Samanfar, B.; Tan, L.H.; Shostak, K.; Chalabian, F.; Wu, Z.; Alamgir, M.; Sunba, N.; Burnside, D.; Omidi, K.; Hooshyar, M.; et al. A global investigation of gene deletion strains that affect premature stop codon bypass in yeast, Saccharomyces cerevisiae. Mol. Biosyst. 2014, 10, 916–924. [Google Scholar] [CrossRef]
- Jeandard, D.; Smirnova, A.; Tarassov, I.; Barrey, E.; Smirnov, A.; Entelis, N. Import of non-coding RNAs into Human mitochondria: A critical review and emerging approaches. Cells 2019, 8, 286. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Aix, C.; García-García, M.; Aranda, M.A.; Sánchez-Pina, M.A. Melon necrotic spot virus Replication Occurs in Association with Altered Mitochondria. Mol. Plant Microbe Interact 2015, 28, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Nibert, M.L.; Debat, H.J.; Manny, A.R.; Grigoriev, I.V.; De Fine Licht, H.H. Mitovirus and mitochondrial coding sequences from basal fungus entomophthora muscae. Viruses 2019, 11, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, I.; Hillman, G.C. Chapter Six—The Family Narnaviridae: Simplest of RNA Viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar] [CrossRef]
- Hillman, B.I.; Annisa, A.; Suzuki, N. Chapter five-Viruses of Plant-Interacting Fungi. Adv. Virus Res. 2018, 100, 99–116. [Google Scholar] [PubMed]
- Mazur, M.; Kmita, H.; Wojtkowska, M. The diversity of the mitochondrial outer membrane protein import channels: Emerging targets for modulation. Molecules 2021, 26, 4087. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H.-W.; Oktay, Y.; Zhang, J.; Allen, E.L.; Smith, G.M.; Fan, K.C.; Hong, J.S.; French, S.W.; Mccaffery, J.M.; et al. PNPASE Regulates RNA Import into Mitochondria. Cell 2010, 142, 456–467. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Basu, S.; Home, P.; Dhar, G.; Adhya, S. Necessary and sufficient factors for the import of transfer RNA into the kinetoplast mitochondrion. EMBO Rep. 2007, 8, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Alston, C.L.; Rocha, M.C.; Lax, N.Z.; Turnbull, D.M.; Taylor, R.W. The genetics and pathology of mitochondrial disease. Pathology 2017, 241, 236–250. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27659608 (accessed on 14 July 2021). [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Peptide Count | |||
|---|---|---|---|
| MT-C1 | MT-C1X | MT-C2 | |
| Mitochondria Protein ID | |||
| TTFMSRK | 11 | --- | --- |
| DQFTRAPTDNDMGVSEATR | 43 | --- | 49 |
| QTTTPTR | 45 | 75 | 67 |
| RWQFNR | 154 | --- | 53 |
| HQQQFFQFRTSGQTMEVTSEYTFR | 236 | 33 | 31 |
| TPHPATTEAK | 21 | --- | --- |
| VDEDQPFPAVPKWSMK | 37 | --- | 13 |
| VTRMVQR | 53 | 58 | 75 |
| WTPAMSERVTR | 86 | 11 | 55 |
| TWSAEIPNTYR | 319 | --- | 191 |
| FNDDFSRAVTEAEVQMCGETRDYTR | 11 | --- | 3 |
| TAVMVTR | 17 | 36 | 63 |
| MTMMTDSTAVVTQRR | 52 | --- | 23 |
| Cytoplasmic protein ID | |||
| GDFQFNISR | --- | 41 | --- |
| QSGFLSQMWIGDKK | --- | 34 | --- |
| QLLTPLR | 44 | 46 | 97 |
| MVQRDR | 45 | 75 | 67 |
| AGENRLAVMVLR | 30 | 40 | 37 |
| LAVMVLRWSDGSYLEDQDMWRMSGIFR | 34 | 3 | 27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajikarimlou, M.; Hooshyar, M.; Sunba, N.; Nazemof, N.; Moutaoufik, M.T.; Phanse, S.; Said, K.B.; Babu, M.; Holcik, M.; Samanfar, B.; et al. A Correlation between 3′-UTR of OXA1 Gene and Yeast Mitochondrial Translation. J. Fungi 2023, 9, 445. https://doi.org/10.3390/jof9040445
Hajikarimlou M, Hooshyar M, Sunba N, Nazemof N, Moutaoufik MT, Phanse S, Said KB, Babu M, Holcik M, Samanfar B, et al. A Correlation between 3′-UTR of OXA1 Gene and Yeast Mitochondrial Translation. Journal of Fungi. 2023; 9(4):445. https://doi.org/10.3390/jof9040445
Chicago/Turabian StyleHajikarimlou, Maryam, Mohsen Hooshyar, Noor Sunba, Nazila Nazemof, Mohamed Taha Moutaoufik, Sadhena Phanse, Kamaledin B. Said, Mohan Babu, Martin Holcik, Bahram Samanfar, and et al. 2023. "A Correlation between 3′-UTR of OXA1 Gene and Yeast Mitochondrial Translation" Journal of Fungi 9, no. 4: 445. https://doi.org/10.3390/jof9040445