
Proposal of Four New Aureobasidium Species for Exopolysaccharide Production


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Strain Isolation
2.2. DNA Isolation, PCR Amplification and Sequencing
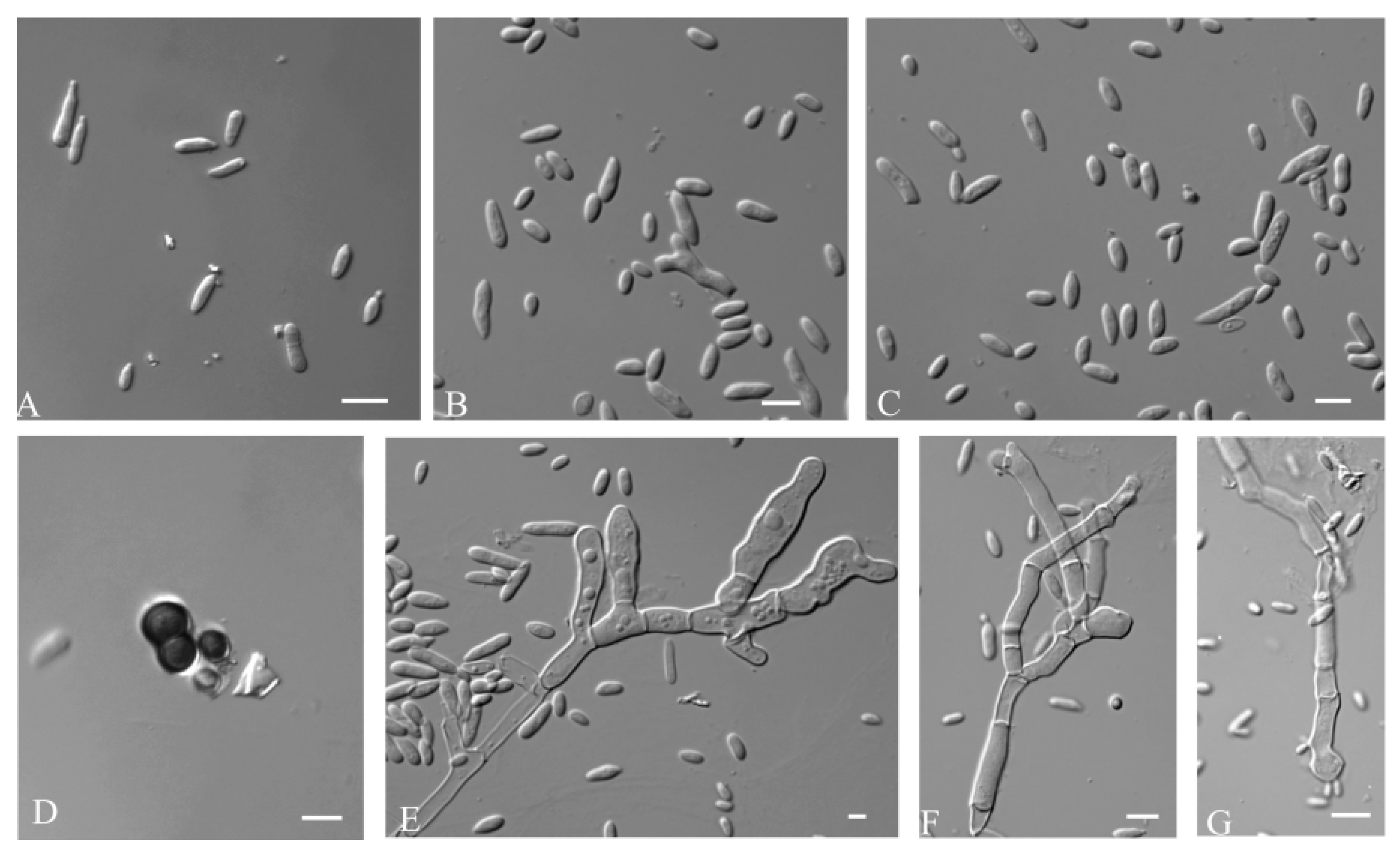
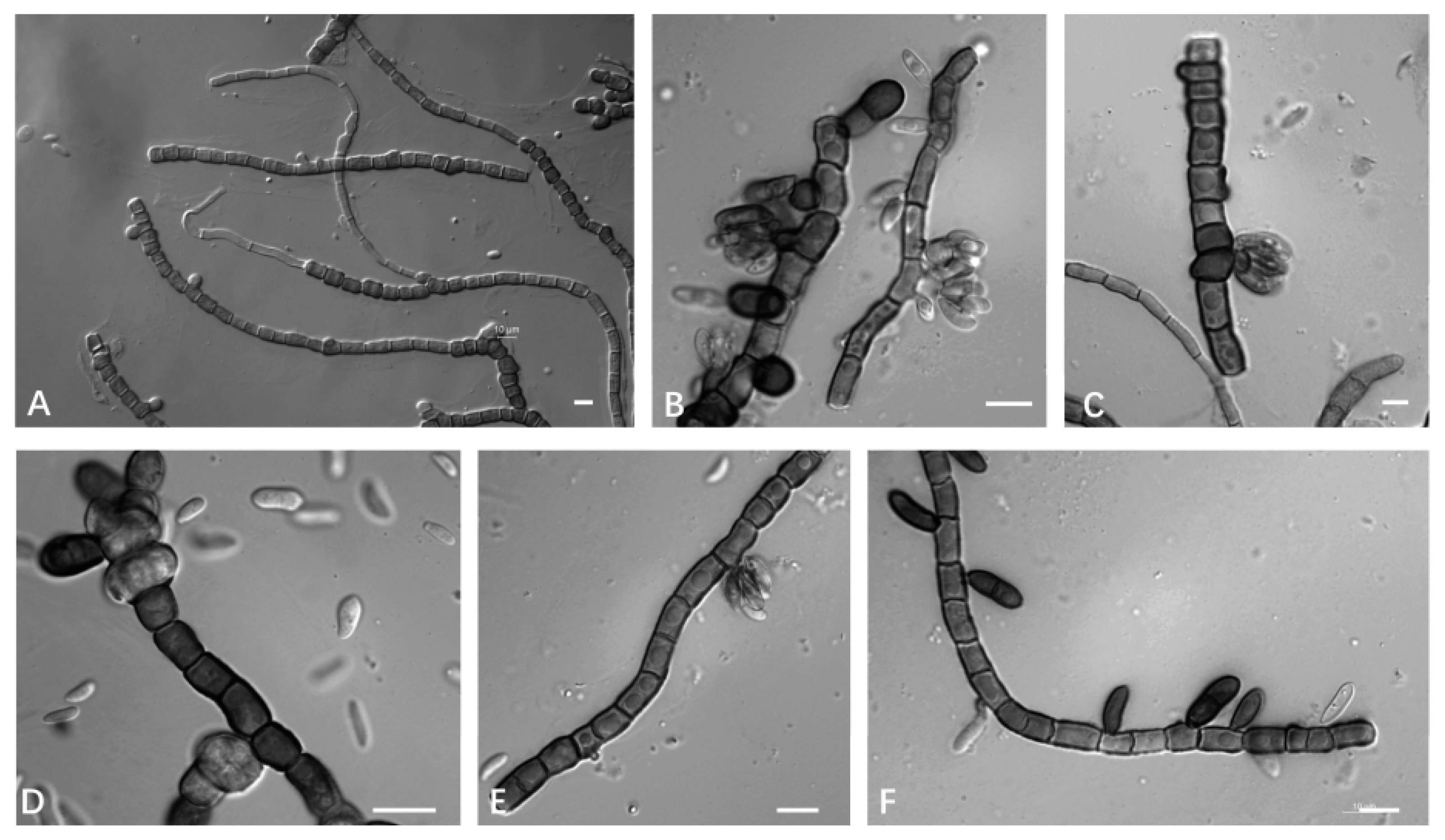
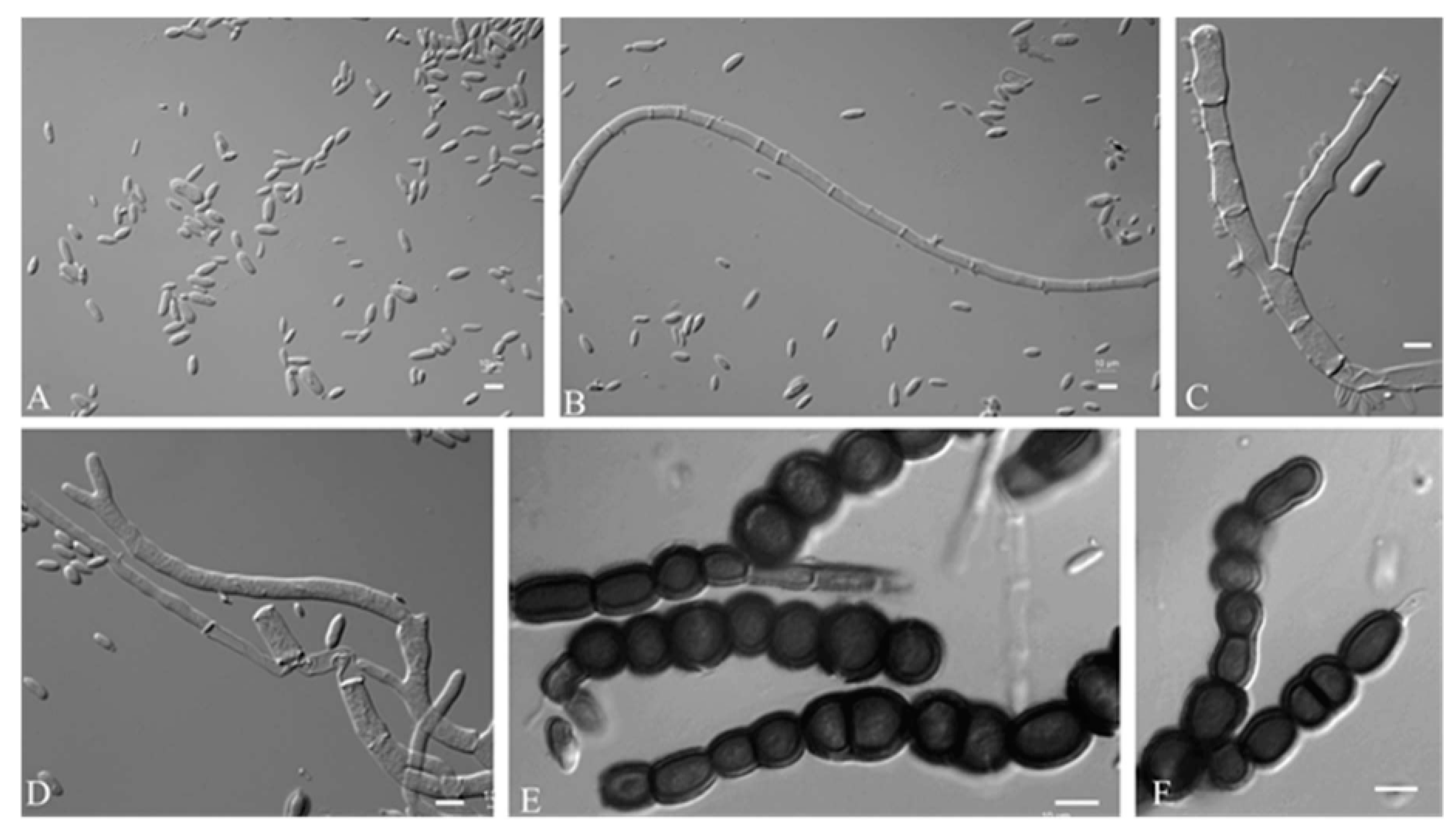
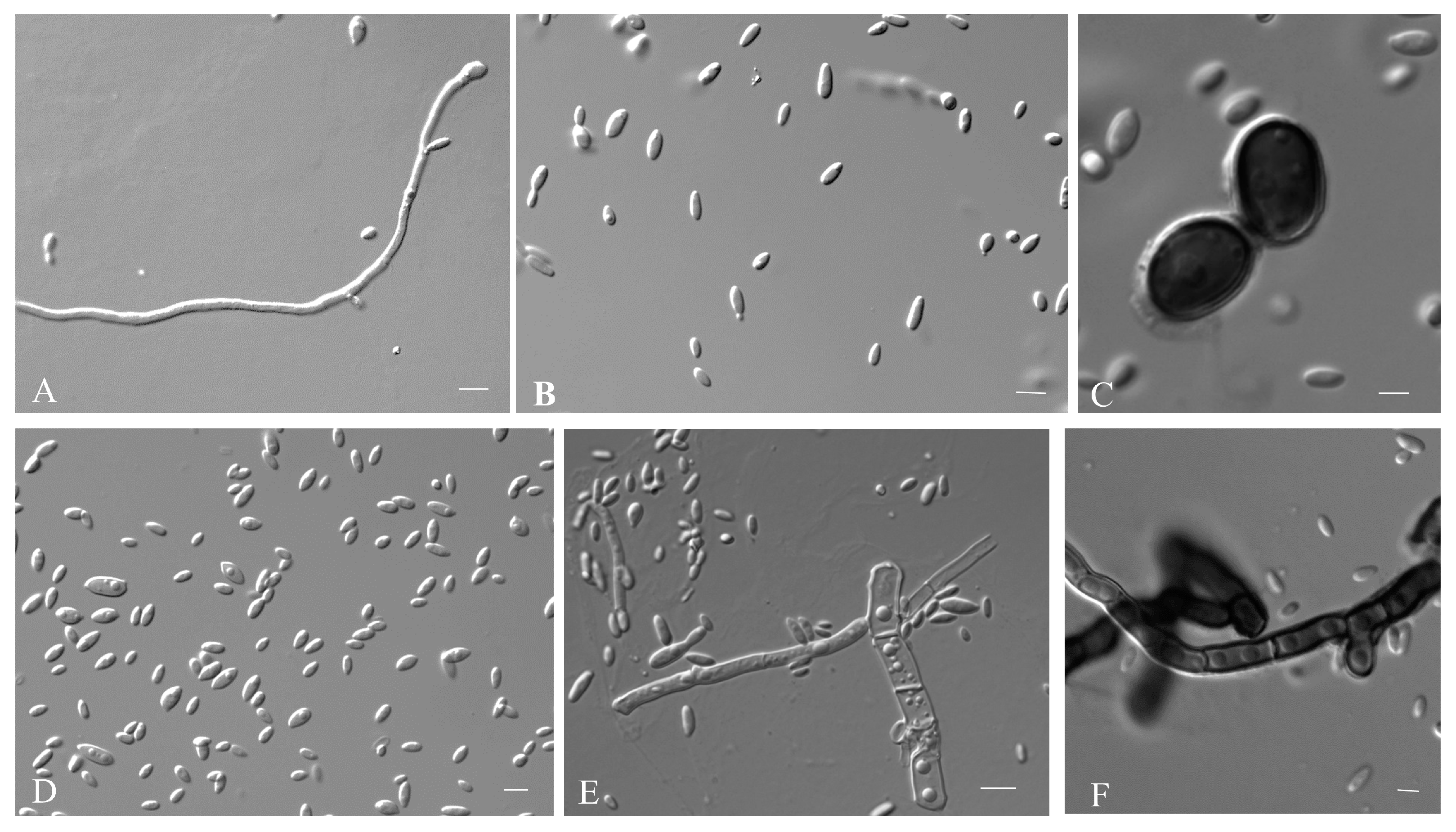
2.3. Observation of Morphology
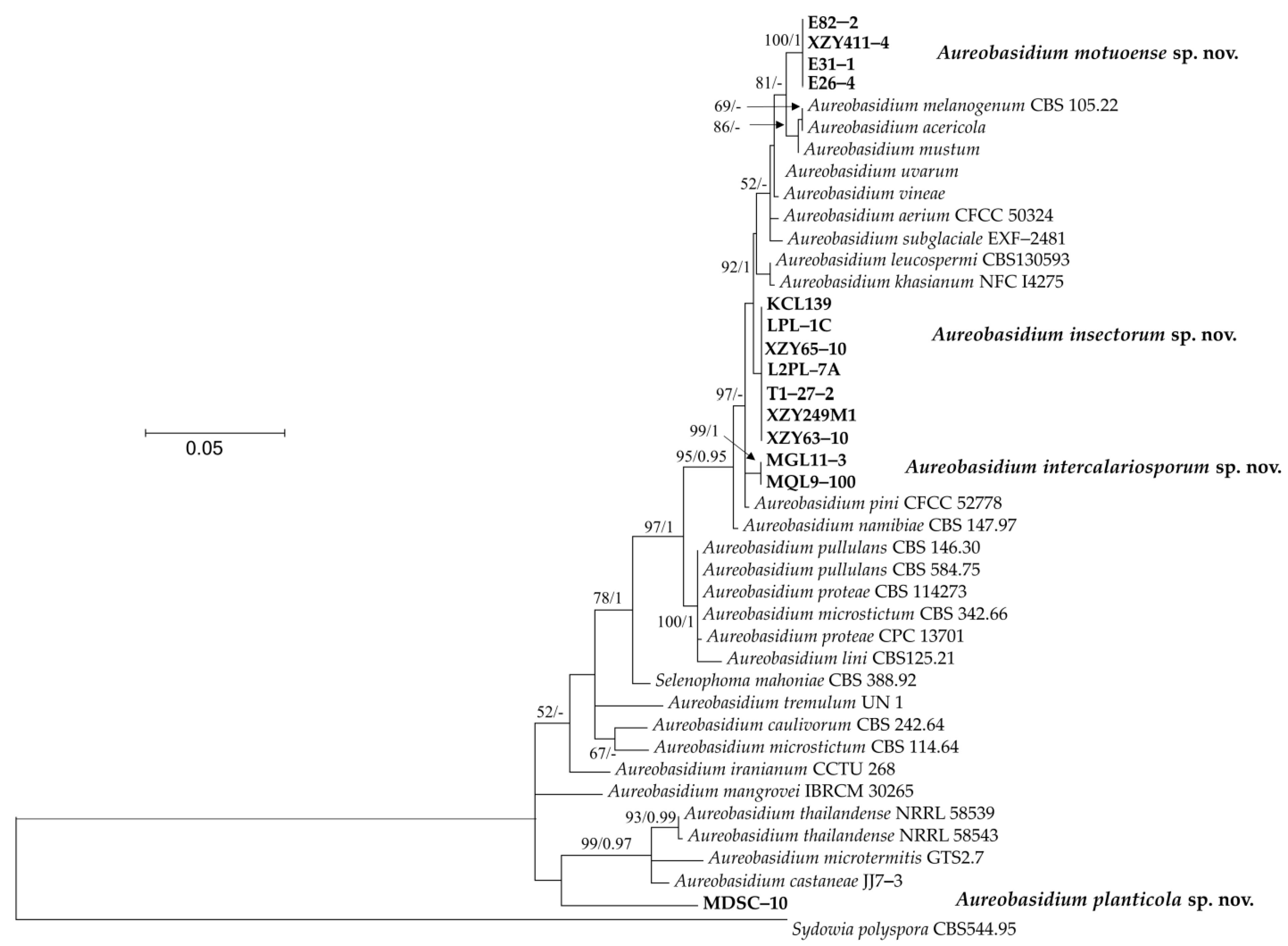
2.4. Phylogenetic Analyses
2.5. Exopolysaccharides Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Date | Location | Latitude and Longitude | Source | GenBank No. | References | |
|---|---|---|---|---|---|---|---|---|
| ITS | D1/D2 | |||||||
| Aureobasidium acericola | CDH 2020−10 | June 2020 | South Korea | 37°45′49.50″ N, 127°11′3.8″ E | Acer pseudosieboldianum | MT863788 | MT863787 | [10] |
| Aureobasidium aerium | CFCC 50324 | April 2015 | Sennon, Beijing, China | NA | air | ON007058 | ON007081 | [11] |
| Aureobasidium castanea | CFCC 54591 * | November 2021 | Jinjing Town, Changsha Hunan, China | 28°58′52″ N, 113°34′38″ E | Castanea heryi | NR_177551 | MW364275 | [12] |
| Aureobasidium caulivorum | CBS 242.64 | NA | Oregon, America | NA | Trifolium incarnatum | FJ150871 | FJ150944 | [39] |
| Aureobasidium insectorum sp. nov. | KCL139 | September 2021 | Zhangjiakou, Hebei, China | 39°30′ N, 113°50′ E | spittle insects | OP856707 | OP857208 | This study |
| LPL−1C | September 2022 | Zhoushan, Zhejiang, China | 29°53′28.86″ N, 122°24′59.35″ E | leaf | OP856705 | OP857207 | This study | |
| XZY65−10 | October 2019 | Shannan City, Tibet, China | 29°14′9.68″ N, 91°45′59.50″ E | leaf | OP856706 | OP857206 | This study | |
| L2PL−7A | September 2022 | Zhoushan, Zhejiang, China | 29°53′28.86″ N, 122°24′59.35″ E | leaf | OP856715 | OP857216 | This study | |
| T1−27−2 | November 2021 | Motuo County, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | leaf | OP856714 | OP857215 | This study | |
| XZY249M1 | October 2019 | Nyingchi City, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | deadwood | OP856713 | OP857214 | This study | |
| XZY63−10 | October 2019 | Shannan City, Tibet, China | 29°14′9.68″ N, 91°45′59.50″ E | leaf | OP856712 | OP857213 | This study | |
| Aureobasidium intercalariosporum sp. nov. | MGL11−3 | September 2022 | Zhoushan, Zhejiang, China | 29°53′28.86″ N, 122°24′59.35″ E | leaf | OP856703 | OP857204 | This study |
| MQL9−100 | September 2022 | Zhoushan, Zhejiang, China | 29°53′28.86″ N, 122°24′59.35″ E | leaf | OP856703 | OP857205 | This study | |
| Aureobasidium iranianum | CCTU 268 | June 2009 | Southern parts of Iran | NA | bamboo stems | NR_137598 | NG_057049 | [13] |
| Aureobasidium khasianum | NFCCI 4275 | December 2016 | Meghalaya, India | NA | litter samples | MH188305 | MH188306 | [40] |
| Aureobasidium leucospermi | CBS 130593 | April 2008 | South Africa | NA | leaves and stems of Proteaceae with cankers or leaf spots | NR_156246 | MH877257 | [14] |
| Aureobasidium lini | CBS 125.21T | NA | UK | NA | Linum usitatissimum | FJ150897 | FJ150946 | [8] |
| Aureobasidium mangrovei | IBRCM 30265T | January 2016 | Qeshm Island, Iran | 26°47′ N, 55°45′ E | mangrove trees (Avicennia marina) | NR_174637 | NG_078639 | [15] |
| Aureobasidium melanogenum | CBS 105.22 | NA | NA | NA | leaf | NR_159598 | NG_056960 | [8] |
| Aureobasidium microstictum | CBS 342.66 | NA | Germany | NA | dying or dead leaves | KT693743 | FJ150945 | [8] |
| Aureobasidium microstictum | CBS 114.64 | NA | Wageningen, The Netherlands | NA | Hemerocallis sp. | KT693744 | KT693986 | [8] |
| Aureobasidium microtermitis | NA | NA | NA | NA | NA | MW276135 | MW276136 | NA |
| Aureobasidium motuoense sp. nov. | E82−2 | October 2019 | Motuo County, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | soil | OP856702 | OP857203 | This study |
| XZY411−4 | August 2019 | Motuo County, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | leaf | OP856710 | OP857211 | This study | |
| E31−1 | October 2019 | Motuo County, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | soil | OP856709 | OP857210 | This study | |
| E26−4 | October 2019 | Motuo County, Tibet, China | 29°19′37.128″ N, 95°19′53.76″ E | soil | OP856708 | OP857209 | This study | |
| Aureobasidium mustum | AWRI 4233 CO−2020 | NA | South Australia | NA | grape juice | NA | NA | [17] |
| Aureobasidium namibiae | CBS 147.97 | 1997 | Namib Desert, Namibia | NA | dolomitic marble | FJ150875 | FJ150937 | [8] |
| Aureobasidium pini | CFCC 52778 | May 2018 | Miyun District, Beijing, China | 40°41′18″ N, 116°55′21″ E | pine needles covered with mycelium | MK184533 | MK184535 | [18] |
| Aureobasidium planticola sp. nov. | MDSC−10 | September 2022 | Zhoushan, Zhejiang, China | 29°53′28.86″ N, 122°24′59.35″ E | leaf | OP856711 | OP857212 | This study |
| Aureobasidium proteae | CBS 114273 | February 2006 | Netherlands | NA | Protea sp. | JN712491 | JN712557 | [15] |
| Aureobasidium proteae | CPC 13701 | July 1998 | Hilly Lands Farm, Somerset West, South Africa | NA | Protea cv. ‘Sylvia’ | JN712490 | JN712556 | [15] |
| Aureobasidium pullulans | CBS 584.75 | 1974 | France | NA | fruit of Vitis vinifera | FJ150906 | FJ150942 | [8] |
| Aureobasidium pullulans | CBS 146.30 | NA | Germany, Ohlsdorf near Hamburg | NA | slime flux of Quercus sp. | FJ150902 | FJ150916 | [8] |
| Aureobasidium subglaciale | EXF−2481 | June and August 2001 | Norway, Svalbard, Kongsvegen | 79° N, 12° E | subglacial ice from seawater | FJ150895 | FJ150913 | [8] |
| Aureobasidium thailandense | NRRL 58539T | 2006 | Nakhonratchasima, Thailand | NA | leaf of Cerbera odollum | JX462674 | JX462674 | [19] |
| Aureobasidium thailandense | NRRL 58543 | 2006 | Prachuapkhirikhan, Thailand | NA | wood surface | JX462675 | JX462675 | [19] |
| Aureobasidium tremulum | UN 1 | NA | NA | NA | NA | MK503657 | MK503660 | NA |
| Aureobasidium uvarum | AWRI 4620 CO−2020 | NA | NA | NA | NA | NA | NA | [17] |
| Aureobasidium vineae | AWRI4619 CO−2020 | NA | NA | NA | NA | NA | NA | [17] |
| Selenophoma mahoniae | CBS 388.92 | NA | Colorado, America | NA | Mahonia repens, leaf | FJ150872 | FJ150943 | [8] |
| Sydowia polyspora | CBS 750.71 | September 1969 | Quebec, Lac Normand, Canada | NA | Pinus strobus, twig | MH872085 | MH872085 | [41] |
3. Results
3.1. Phylogeny
3.2. Taxonomy
3.2.1. Aureobasidium insectorum Q.M. Wang, F. Wu & M.M. Wang sp. nov.
3.2.2. Aureobasidium planticola Q.M. Wang, F. Wu & M.M. Wang sp. nov.
3.2.3. Aureobasidium motuoense Q.M. Wang, F. Wu & M.M. Wang sp. nov.
3.2.4. Aureobasidium intercalariosporum Q.M. Wang, F. Wu & M.M. Wang sp. nov.
3.3. Exopolysaccharides Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hymphries, Z.; Seifert, K.A.; Hirooka, Y.; Visagie, M. A new family and genus in Dothideales for Aureobasidium-like species isolated from house dust. IMA Fungus 2017, 8, 299–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thambugala, K.M.; Ariyawansa, H.A.; Li, Y.M.; Boonmee, S.; Hongsanan, S.; Tian, Q.; Singtripop, C.; Bhat, D.J.; Camporesi, E.; Jayawardena, R.; et al. Dothideales. Fungal Divers. 2014, 68, 105–158. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Dai, D.Q.; D’souza, M.J.; Diederich, P.; Dissanayake, A.; et al. Naming and outline of Dothideomycetes–2014 including proposals for the protection or suppression of generic names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Hermanides-Nijhof, E.J. Aureobasidium and allied genera. Stud. Mycol. 1977, 15, 141–177. [Google Scholar]
- De Hoog, G.S.; Yurlova, N.A. Conidiogenesis, nutritional physiology and taxonomy of Aureobasidium and Hormonema. Antonie Van Leeuwenhoek 1994, 65, 41–54. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Zalar, P.; Urzi, C.; De Leo, F.; Yurlova, N.A.; Sterflinger, K. Relationships of dothideaceous black yeasts and meristematic fungi based on 5.8 S and ITS2 rDNA sequence comparison. Stud. Mycol. 1999, 43, 31–37. [Google Scholar]
- Yurlova, N.A.; Uijthof, J.M.J.; De Hoog, G.S. Distinction of species in Aureobasidium and related genera by PCR-ribotyping. Antonie Van Leeuwenhoek 1996, 69, 323. [Google Scholar] [CrossRef]
- Zalar, P.; Gostinčar, C.; De Hoog, G.S.; Uršič, V.; Sudhadham, M.; Gunde-Cimerman, N. Redefinition of Aureobasidium pullulans and its varieties. Stud. Mycol. 2008, 61, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome sequencing of four Aureobasidium pullulans varieties: Biotechnological potential, stress tolerance, and description of new species. BMC Genom. 2014, 15, 549. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Cho, S.E.; Oh, J.Y.; Cho, E.J.; Kwon, S. A novel species of Aureobasidium (Dothioraceae) recovered from Acer pseudosieboldianum in Korea. J. Asia-Pac. Biodivers. 2021, 14, 657–661. [Google Scholar] [CrossRef]
- Wang, C.B.; Jiang, N.; Zhu, Y.Q.; Xue, H.; Li, Y. Aureobasidium aerium (Saccotheciaceae, Dothideales), a new yeast-like fungus from the air in Beijing, China. Phytotaxa 2022, 544, 185–192. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, X.; Tian, C. Identification and Characterization of Leaf-Inhabiting Fungi from Castanea Plantations in China. J. Fungi 2021, 18, 64. [Google Scholar] [CrossRef]
- Arzanlou, M. Aureobasidium iranianum, a new species on bamboo from Iran. Mycosphere 2012, 3, 404–408. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Swart, L.; Denman, S.; Taylor, J.E.; Bezuidenhout, C.M.; Palm, M.E.; Marincowitz, S.; Groenewald, J.Z. Fungal pathogens of Proteaceae. Persoonia 2011, 27, 20–45. [Google Scholar] [CrossRef] [Green Version]
- Nasr, S.; Mohammadimehr, M.; Geranpayeh Vaghei, M.; Amoozegar, M.A.; Shahzadeh Fazeli, S.A. Aureobasidium mangrovei sp. nov., an ascomycetous species recovered from Hara protected forests in the Persian Gulf, Iran. Antonie Van Leeuwenhoek 2018, 111, 1697–1705. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.; Jansen, G.M.; et al. Fungal Planet description sheets. Persoonia 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Onetto, C.A.; Schmidt, S.A.; Roach, M.J.; Borneman, A.R. Comparative genome analysis proposes three new Aureobasidium species isolated from grape juice. FEMS Yeast Res. 2020, 20, 52. [Google Scholar] [CrossRef]
- Jiang, N.; Liang, Y.M.; Tian, C.M. Aureobasidium pini sp. nov. from pine needle in China. Phytotaxa 2019, 402, 10. [Google Scholar] [CrossRef]
- Peterson, S.; Manitchotpisit, P.; Leathers, T. Aureobasidium thailandense sp. nov. isolated from leaves and wooden surfaces. Int. J. Syst. Evol. Microbiol. 2012, 63, 790–795. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets. Persoonia 2019, 44, 291–473. [Google Scholar] [CrossRef]
- Prasongsuk, S.; Lotrakul, P.; Ali, I.; Bankeeree, W.; Punnapayak, H. The current status of Aureobasidium pullulans in biotechnology. Folia Microbiol. 2018, 63, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Manitchotpisit, P.; Leathers, T.D.; Peterson, S.W.; Kurtzman, C.P.; Li, X.L.; Eveleigh, D.E.; Lotrakul, P.; Prasongsuk, S.; Dunlap, C.A.; Vermillion, K.E.; et al. Multilocus phylogenetic analyses, pullulan production and xylanase activity of tropical isolates of Aureobasidium pullulans. Mycol. Res. 2009, 113, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Kutleša, M.; Mlinarić-Missoni, E.; Hatvani, L.; Voncina, D.; Simon, S.; Lepur, D.; Baršić, B. Chronic fungal meningitis caused by Aureobasidium proteae. Diagn. Microbiol. Infect. Dis. 2012, 73, 271–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leathers, T.D. Biotechnological production and applications of pullulan. Appl. Microbiol. Biotechnol. 2002, 62, 468–473. [Google Scholar] [CrossRef]
- Cheng, K.C.; Demirci, A.; Catchmark, J.M. Pullulan: Biosynthesis, production, and applications. Appl. Microbiol. Biotechnol. 2011, 92, 29–44. [Google Scholar] [CrossRef]
- Manitchotpisit, P.; Skory, C.D.; Peterson, S.W.; Price, N.P.J.; Vermillion, K.E.; Leathers, T.D. Poly (β-L-malic acid) production by diverse phylogenetic clades of Aureobasidium pullulans. J. Ind. Microbiol. Biotechnol. 2012, 39, 125–132. [Google Scholar] [CrossRef]
- Muthusamy, S.; Anandharaj, S.J.; Kumar, P.S. Microbial pullulan for food, biomedicine, cosmetic, and water treatment: A review. Environ. Chem. Lett. 2022, 20, 3199–3234. [Google Scholar] [CrossRef]
- Kang, X.X.; Jia, S.L.; Wei, X.; Zhang, M.; Liu, G.L.; Hu, Z.; Chi, Z.; Chi, Z.M. Liamocins biosynthesis, its regulation in Aureobasidium spp., and their bioactivities. Crit. Rev. Biotechnol. 2022, 42, 93–105. [Google Scholar] [CrossRef]
- Suzuki, T.; Kusano, K.; Kondo, N.; Nishikawa, K.; Kuge, T.; Ohno, N. Biological Activity of High-Purity β-1,3-1,6-Glucan Derived from the Black Yeast Aureobasidium pullulans: A Literature Review. Nutrients 2021, 13, 242. [Google Scholar] [CrossRef]
- Xin, Z.; Chen, J. A high throughput DNA extraction method with high yield and quality. Plant Methods 2012, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification, and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Lin, D.; Wu, L.C.; Rinaldi, M.G.; Lehmann, P.F. Three distinct genotypes within Candida parapsilosis from clinical sources. J. Clin. Microbiol. 1995, 33, 1815–1821. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5′ end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; Begerow, D.; Groenewald, M.; Liu, X.; Theelen, B.; Bai, F.Y.; Boekhout, T. Multigene phylogeny and taxonomic revision of yeasts and related fungi in the Ustilaginomycotina. Stud. Mycol. 2015, 81, 55–83. [Google Scholar] [CrossRef] [Green Version]
- Haghighatpanah, N.; Mirzaee, H.; Khodaiyan, F.; Kennedy, J.F.; Aghakhani, A.; Hosseini, S.S.; Jahanbin, K. Optimization and characterization of pullulan produced by a newly identified strain of Aureobasidium pullulans. Int. J. Biol. Macromol. 2020, 152, 305–313. [Google Scholar] [CrossRef]
- Buksa, K.; Kowalczyk, M.; Boreczek, J. Extraction, purification and characterisation of exopolysaccharides produced by newly isolated lactic acid bacteria strains and the examination of their influence on resistant starch for-mation. Food Chem. 2021, 362, 130221. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Prabhugaonkar, A.; Jalmi, P. Aureobasidium khasianum (Aureobasidiaceae) a novel species with distinct morphology. Phytotaxa 2018, 374, 257. [Google Scholar] [CrossRef]
- Verkley, G.J.; Starink-Willemse, M.; van Iperen, A.; Abeln, E.C. Phylogenetic analyses of Septoria species based on the ITS and LSU-D2 regions of nuclear ribosomal DNA. Mycologia 2004, 96, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boundy-Mills, K.; Čadež, N.; Endoh, R.; Jindamorakot, S.; Pohl-Albertyn, C.; Rosa, C.A.; Turchetti, B.; Yurkov, A. Census of yeasts isolated from natural ecosystem and conserved in worldwide collections. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Berlin, Germany, 2017; pp. 455–476. [Google Scholar]
- Blackwell, M. Yeasts in Insects and Other Invertebrates. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Berlin, Germany, 2017; pp. 397–433. [Google Scholar] [CrossRef]
- Boekhout, T.; Amend, A.S.; EI Baidouri, F.; Gabaldón, T.; GemI, J.; Mittelbach, M.; Rober, V.; Tan, C.S.; Turchetti, B.; Vu, D.; et al. Trends in yeast diversity discovery. Fungal Divers. 2022, 114, 491–537. [Google Scholar] [CrossRef]
- Boekhout, T. Biodiversity: Gut feeling for yeasts. Nature 2005, 434, 449–451. [Google Scholar] [CrossRef] [PubMed]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect-yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Cheng, C.; Feng, J.; Song, X.; Lin, M.; Yang, S.T. Biosynthesis of polymalic acid in fermentation: Advances and prospects for industrial application. Crit. Rev. Biotechnol. 2019, 39, 408–421. [Google Scholar] [CrossRef]






| Strain | Species | Fermentation Liquid Color | Exopolysaccharides Yield (g/L) | Average Weight (g/L) |
|---|---|---|---|---|
| PTSL5−5 | A. thailandense | Light yellow | 8.47 | 6.17 |
| PTSL4−6 | A. thailandense | Light yellow | 8.09 | |
| PTSL5−3 | A. thailandense | Light yellow | 2.71 | |
| PTSL11−5 | A. thailandense | Light yellow | 5.42 | |
| PTSL9−106 | A. melanogenum | Pink | 1.53 | 39.06 |
| PTSL19−101 | A. melanogenum | Light yellow | 32.53 | |
| PTSL6−101 | A. melanogenum | Light yellow | 34.50 | |
| PTSL19−107 | A. melanogenum | Yellow | 41.84 | |
| PTSL19−104 | A. melanogenum | Yellow | 54.58 | |
| PTSL20−102 | A. melanogenum | Light yellow | 48.13 | |
| PTSL20−104 | A. melanogenum | Yellow | 52.33 | |
| PTSL19−104 | A. melanogenum | Light yellow | 45.71 | |
| PTSL19−104 | A. melanogenum | Light yellow | 45.71 | |
| PTSL17−4 | A. melanogenum | Light yellow | 34.36 | |
| PTSL9−100 | A. melanogenum | Light yellow | 38.45 | |
| LF75−2 | A. leucospermi | Light yellow | 0.92 | 17.24 |
| SXY35−16 | A. leucospermi | Light yellow | 28.94 | |
| SXY35−15 | A. leucospermi | Light yellow | 23.37 | |
| LF45−2 | A. leucospermi | Light yellow | 15.75 | |
| LPL−7A | A. insectorum | Light yellow | 27.67 | 14.7 |
| KCL139 | A. insectorum | Dark yellow | 8.64 | |
| XZY65−10 | A. insectorum | Dark yellow | 7.80 | |
| E26−4 | A. motuoense | Yellow | 15.74 | 26.57 |
| E31−1 | A. motuoense | Dark yellow | 21.39 | |
| XZY411−4 | A. motuoense | Dark yellow | 31.72 | |
| E82−2 | A. motuoense | Dark yellow | 37.43 | |
| MGL11−3 | A. intercalariosporum | Light yellow | 29.43 | 31.79 |
| MQL9−100 | A. intercalariosporum | Light yellow | 34.15 | |
| MDSC−10 | A. planticola | Black | 2.10 | 2.1 |
| Aureobasidium planticola | Aureobasidium intercalariosporum | Aureobasidium motuoense | Aureobasidium insectorum | |
|---|---|---|---|---|
| PDA | 22–25 | 32–36 | 34–38 | 32–35 |
| M40Y | 33–40 | 35–40 | 41–43 | 32–34 |
| M60Y | 27–34 | 29–34 | 36–40 | 25–27 |
| MEA + 5% NaCl | 14–15 | 14–17 | 13–15 | 9–12 |
| MEA + 10% NaCl | 7–8 | 8–11 | 10–11 | 8–8 |
| MEA + 15% NaCl | 0 | 9–13 | 5–8 | 8–9 |
| MEA + 20% NaCl | 0 | 0 | 0 | 0 |
| MEA at 4 °C | 0 | 0 | 0 | 5–5 |
| MEA at 17 °C | 10–13 | 13–15 | 8–13 | 12–15 |
| MEA at 28 °C | 27–27 | 24–25 | 32–35 | 28–28 |
| MEA at 30 °C | 11–10 | 13–14 | 38–44 | 7–8 |
| MEA at 37 °C | 0 | 0 | 7–8 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Feng, Z.; Wang, M.; Wang, Q. Proposal of Four New Aureobasidium Species for Exopolysaccharide Production. J. Fungi 2023, 9, 447. https://doi.org/10.3390/jof9040447
Wu F, Feng Z, Wang M, Wang Q. Proposal of Four New Aureobasidium Species for Exopolysaccharide Production. Journal of Fungi. 2023; 9(4):447. https://doi.org/10.3390/jof9040447
Chicago/Turabian StyleWu, Feng, Zixuan Feng, Manman Wang, and Qiming Wang. 2023. "Proposal of Four New Aureobasidium Species for Exopolysaccharide Production" Journal of Fungi 9, no. 4: 447. https://doi.org/10.3390/jof9040447