
First Report on Colletotrichum fructicola Causing Anthracnose in Chinese Sorghum and Its Management Using Phytochemicals

Abstract
:1. Introduction
2. Materials and Methods
2.1. Diseased Leaf Collection, Fungus Isolation, and Phytochemicals
2.2. Pathogen Identification
2.3. Pathogenicity Assays
2.4. Antimicrobial Activity of Phytochemicals against Mycelial Growth
2.5. Evaluations of Field Trials of Phytochemicals to Control the Disease Caused by C. fructicola
2.6. Statistical Analyses
3. Results
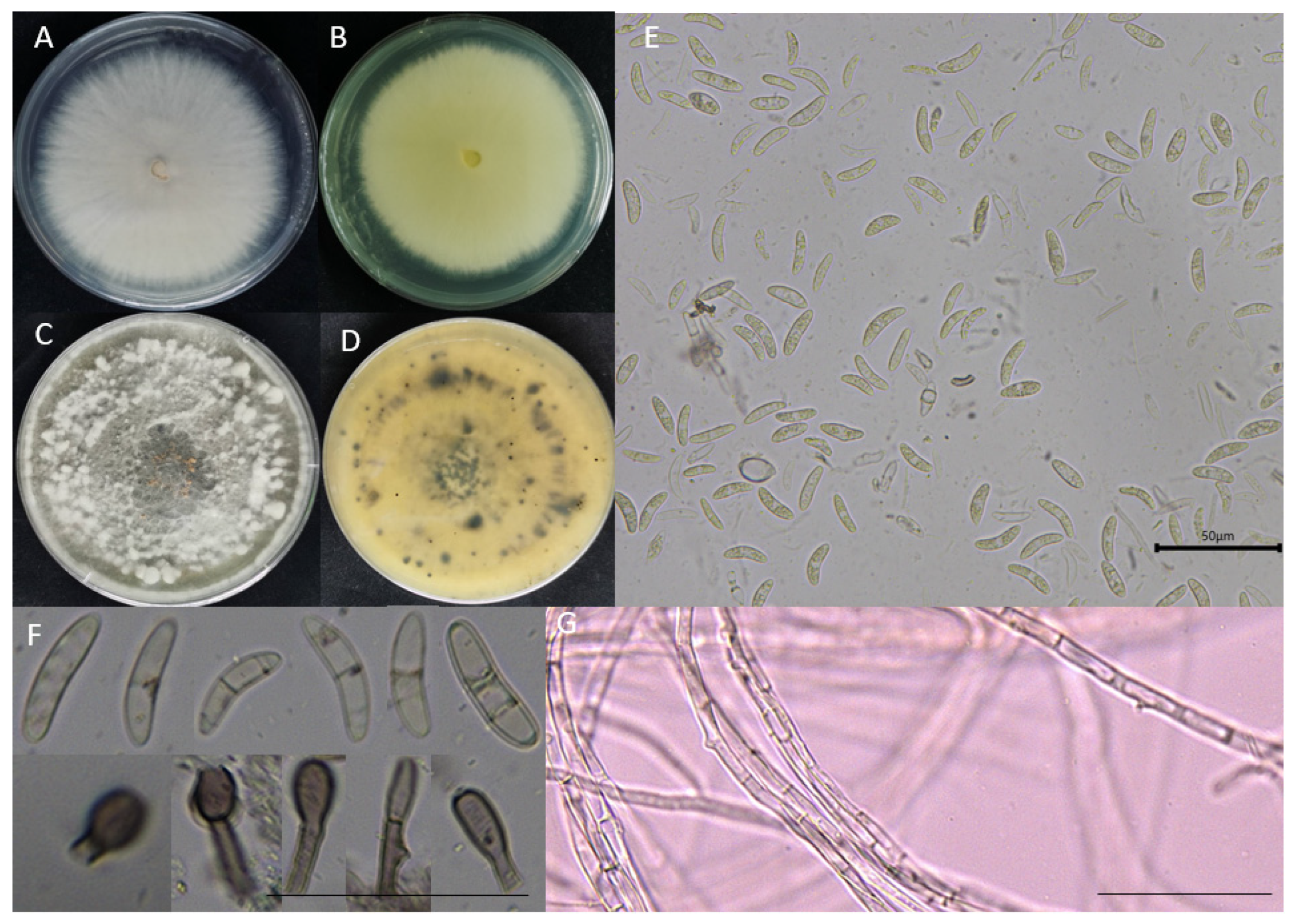
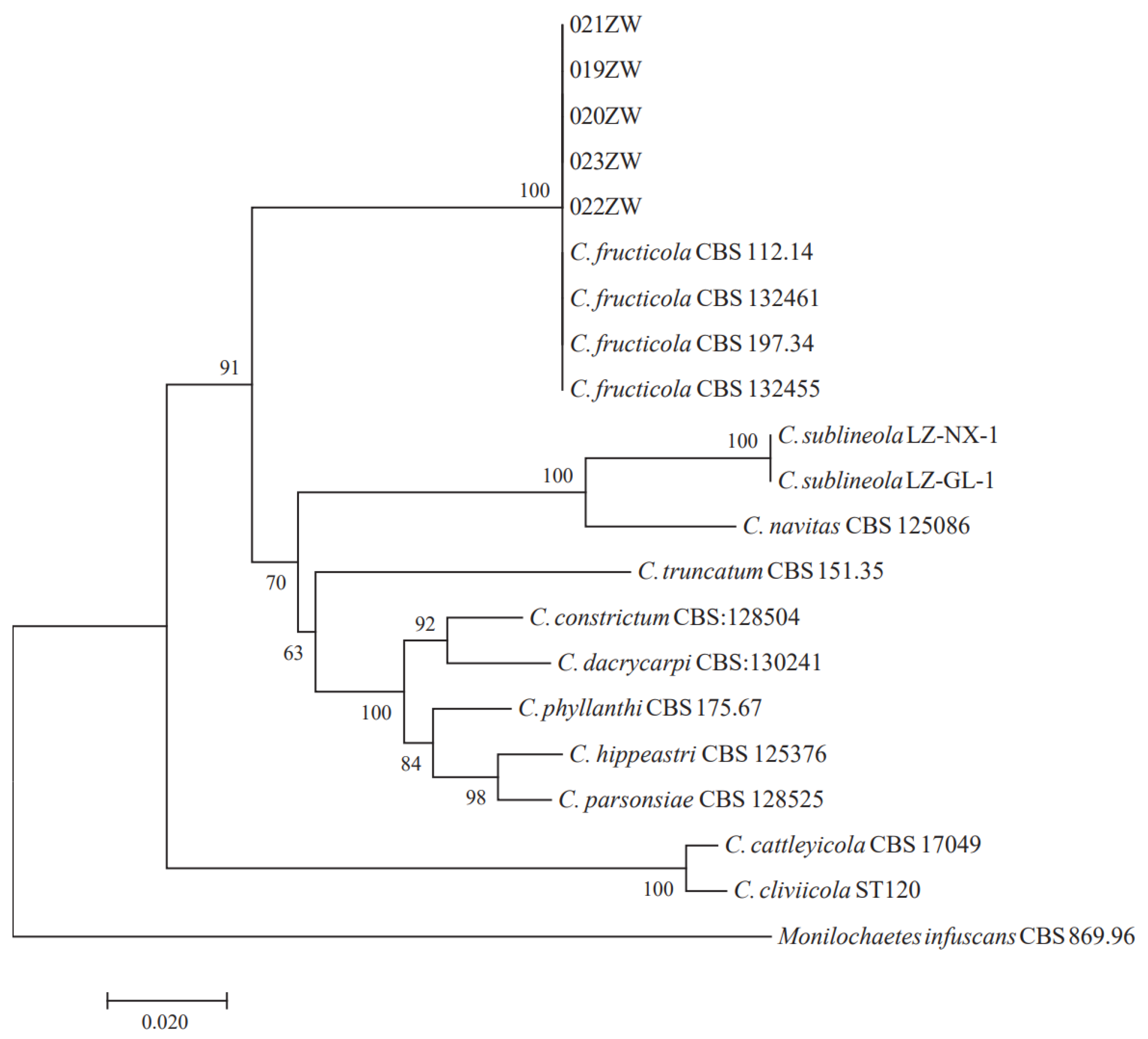
3.1. Isolation and Identification of Strain 022ZW from Sorghum Leaves
3.2. Phytochemical Sensitivity of Isolate 022ZW
3.3. Controlling Sorghum Leaf Spots Caused by C. fructicola Using Phytochemicals in the Field
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gerrano, A.S.; Labuschagne, M.T.; van Biljon, A.; Shargie, N.G. Quantification of Mineral Composition and Total Protein Content in Sorghum [Sorghum Bicolor (L.) Moench] Genotypes. Cereal Res. Commun. 2016, 44, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Prasad, V.B.R.; Govindaraj, M.; Djanaguiraman, M.; Djalovic, I.; Shailani, A.; Rawat, N.; Singla-Pareek, S.L.; Pareek, A.; Prasad, P.V.V. Drought and High Temperature Stress in Sorghum: Physiological, Genetic, and Molecular Insights and Breeding Approaches. Int. J. Mol. Sci. 2021, 22, 9826. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Qiu, H.B.; Long, H.J.; Zhao, W.; Hu, A.L. First Report of Leaf Spot on Sorghum caused by Pestalotiopsis trachycarpicola in china. Plant Pathol. 2021, 103, 1043–1044. [Google Scholar] [CrossRef]
- Yang, S.Y.; Feng, M.C.; Lu, J.; Zhang, H.Z.; Fan, J.; Zhao, W.; Hu, A. Bioactivity of 8 Plant Extracts against Pathogen of Sorghum Leaf Spot. World J. Agric. Med. 2021, 43, 53–57. [Google Scholar]
- Zhao, Y.Q.; Yu, H.R.; Shi, K.; Zhang, D.M.; Zhu, P.T.; Cai, C.; Jia, A.M. Pathogen Identification of Sorghum Alternaria alternaria Leaf Spot Disease. Chin. J. Plant Pathol. 2017, 47, 282–285. [Google Scholar]
- Jonglaekha, N.; Kongjit, J. Red Leaf Spot of Sorghum Caused by Curvularia clavata Jain, A New Disease in Thailand; Thailand National Corn and Sorghum Program 1981 Annual Report; Thailand National Corn and Sorghum Program: Bangkok, Thailand, 1981; pp. 174–178. [Google Scholar]
- Wang, C.L. The Occurrence and Prevention of major Diseases of Sorghum in Heilongjiang Province. Chin. Mod. Agric. Sci. Technol. 2018, 119+123. [Google Scholar]
- Klittich, C.; Leslie, J.F.; Nelson, P.E.; Marasas, W.F.O. Fusarium thapsinum (Gibberella thapsina): A new Species in Section Liseola from Sorghum. Mycologia 1997, 89, 643–652. [Google Scholar] [CrossRef]
- Cota, L.V.; Costa, R.V.; Silva, D.D.; Parreira, D.F.; Lana, U.G.P.; Casela, C.R. First Report of Pathogenicity of Pantoea ananatis in Sorghum (Sorghum Bicolor) in Brazil. Australas. Plant Dis. Notes 2010, 5, 120–122. [Google Scholar] [CrossRef]
- Lim, S.; Yoon, Y.; Jang, Y.W.; Bae, D.H.; Kim, B.S.; Maharjan, R.; Yi, H.; Bae, S.; Lee, Y.; Lee, C.; et al. First Report of Maize Yellow Mosaic Virus Infecting Panicum miliaceum and Sorghum bicolor in South Korea. Plant Dis. 2018, 102, 689-689. [Google Scholar] [CrossRef]
- Evallo, E.; Taguiam, J.D.; Bengoa, J. First report of Colletotrichum fructicola, causing Anthracnose of Hylocereus Plants, in the Philippines. Czech Mycol. 2021, 73, 79–90. [Google Scholar] [CrossRef]
- Narmadhavathy, S.; Nayar, K.; Gokulapalan, C.; Geetha, D. First Report of Leaf Spot Disease of Culinary Melon caused by Colletotrichum fructicola in India. Indian Phytopathol. 2016, 69, 318–319. [Google Scholar]
- Lin, S.R.; Yu, S.Y.; Chang, T.D.; Lin, Y.J.; Wen, C.J.; Lin, Y.H. First Report of Anthracnose Caused by Colletotrichum fructicola on Tea in Taiwan. Plant Dis. 2021, 105, 710-710. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.H.; Li, D.W.; Duan, C.H.; Liu, X.H.; Niu, S.G.; Hou, L.Q.; Wu, X.Q. First Report of Walnut Anthracnose Caused by Colletotrichum fructicola in China. Plant Dis. 2018, 102, 247-247. [Google Scholar] [CrossRef]
- Jiang, G.H.; Jiang, A.M.; Fan, C.L.; Wei, J.G.; Ren, L.Y.; Luo, J.T. First Report of Anthracnose on Kadsura coccinea Caused by Colletotrichum fructicola in China. Plant Dis. 2022, 106, 1757. [Google Scholar] [CrossRef]
- Wang, S.M.; Huang, J.; Zheng, M.; Wang, Y.; Yuan, Q.; Gao, Q.; Zhou, H. First Report of Anthracnose on Bletilla striata Caused by Colletotrichum fructicola in China. Plant Dis. 2022, 106, 756. [Google Scholar] [CrossRef]
- Liu, Q.; Qiao, K.; Zhang, S. Potential of a Small Molecule Carvacrol in Management of Vegetable Diseases. Molecules 2019, 24, 1932. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.; Chen, J.; Shi, Z. The Fungicidal Activity of Thymol against Fusarium graminearum via Inducing Lipid Peroxidation and Disrupting Ergosterol Biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Yan, L.; Zhang, M.; Chang, X.; Zhu, D.; Wei, D.; Naeem, M.; Song, C.; Wu, X.; Liu, T.; et al. Changes in the Density and Composition of Rhizosphere Pathogenic Fusarium and Beneficial Trichoderma Contributing to Reduced Root Rot of Intercropped Soybean. Pathogens 2022, 11, 478. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Molecular Phylogeny of the Higher and Lower Taxonomy of the Fusarium Genus and Differences in the Evolutionary Histories of Multiple Genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, X.; Sun, B.; Sun, L. The Involvement of Thiamine Uptake in the Virulence of Edwardsiella piscicida. Pathogens 2022, 11, 464. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and Molecular Characterization of the Glyceraldehyde-3-phosphate Dehydrogenase-encoding Gene and cDNA from the Plant Pathogenic Fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of Primer sets designed for use with the PCR to Amplify conserved Genes from Filamentous scomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Wu, X.; Dai, Y.; Yin, X.; Zhao, Z.; Zhang, Z.; Li, W.; He, L.; Long, Y. Sensitivity Testing of Natural Antifungal Agents on Fusarium fujikuroi to Investigate the Potential for Sustainable Control of Kiwifruit Leaf Spot Disease. J. Fungi 2022, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Mao, W.; Lu, F.; Li, T.; Wang, J.; Duan, Y.; Zhou, M. In vitro fungicidal Activity and in Planta Control Efficacy of Coumoxystrobin against Magnaporthe oryzae. Pestic. Biochem. Physiol. 2020, 162, 78–85. [Google Scholar] [CrossRef]
- Mo, F.; Hu, X.; Ding, Y.; Li, R.; Li, M. Naturally Produced Magnolol can significantly damage the Plasma Membrane of Rhizoctonia solani. Pestic. Biochem. Physiol. 2021, 178, 104942. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, R.; Vicente, D.L.; Paula, C.; Victoria, G.; Mario, R.; Devani, A.P.; Castagnaro, L.; Ploper, D. Evaluation of the Efficacy and Application timing of different Fungicides for Management of Soybean Foliar Diseases in Northwestern Argentina. Crop Prot. 2019, 124, 104844. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of Development Descriptions for Soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Li, W.; Long, Y.; Mo, F.; Shu, R.; Yin, X.; Wu, X.; Zhang, R.; Zhang, Z.; He, L.; Chen, T.; et al. Antifungal Activity and Biocontrol Mechanism of Fusicolla violacea J-1 against Soft Rot in Kiwifruit Caused by Alternaria alternata. J. Fungi 2021, 7, 937. [Google Scholar] [CrossRef]
- Zhang, L.; Song, L.; Xu, X.; Zou, X.; Duan, K.; Gao, Q. Characterization and Fungicide Sensitivity of Colletotrichum Species Causing Strawberry Anthracnose in Eastern China. Plant Dis. 2020, 104, 1960–1968. [Google Scholar] [CrossRef]
- Dahlberg, J.; Berenji, J.; Sikora, V.; Latković, D. Assessing Sorghum [Sorghum bicolor (L) Moench] Germplasm for new Traits: Food, Fuels & unique Uses. Maydica 2012, 56, 1750. [Google Scholar]
- McCormick, R.F.; Truong, S.K.; Sreedasyam, A.; Jenkins, J.; Shu, S.; Sims, D.; Kennedy, M.; Amirebrahimi, M.; Weers, B.D.; McKinley, B.; et al. The Sorghum bicolor Reference Genome: Improved assembly, Gene Annotations, a Transcriptome Atlas, and signatures of Genome Organization. Plant J. 2018, 93, 338–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, B.D. Sorghum Value Chain for Food and Fodder Security. In Breeding Sorghum for Diverse End Uses; Woodhead Publishing: Sawston, UK, 2019; pp. 409–419. [Google Scholar]
- Li, Q.; Bu, J.; Shu, J.; Yu, Z.; Tang, L.; Huang, S.; Guo, T.; Mo, J.; Luo, S.; Solangi, G.S.; et al. Colletotrichum Species associated with Mango in Southern China. Sci. Rep. 2019, 9, 18891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Luo, C.-X.; Wu, H.-J.; Peng, B.; Kang, B.-S.; Liu, L.-M.; Zhang, M.; Gu, Q.-S. Colletotrichum Species Associated with Anthracnose Disease of Watermelon (Citrullus lanatus) in China. J. Fungi 2022, 8, 790. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fu, D.; Wang, W.; Gleason, M.L.; Zhang, R.; Liang, X.; Sun, G. Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China. J. Fungi 2022, 8, 740. [Google Scholar] [CrossRef]
- Jing, J.J.; Hong, Y.Z.; Li, H.; Wang, Z.; Chen, Y.; Hong, N.; Wang, G.; Chofong, G.N.; Xu, W. Identification and Characterization of Colletotrichum fructicola causing Black Spots on Young Fruits related to bitter Rot of Pear (Pyrus bretschneideri Rehd.) in China. Crop Prot. 2014, 58, 41–48. [Google Scholar] [CrossRef]
- Saxena, A.; Raghuwanshi, R.; Gupta, V.K.; Singh, H.B. Chilli Anthracnose: The Epidemiology and Management. Front. Microbiol. 2016, 7, 1527. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu, H.; Hu, M.; Wu, J.; Zhang, C. Fungal Pathogens Associated with Strawberry Crown Rot Disease in China. J. Fungi 2022, 8, 1161. [Google Scholar] [CrossRef]
- Slippers, B.; Stenlid, J.; Wingfield, M.J. Emerging pathogens: Fungal Host Jumps following Anthropogenic Introduction. Trends Ecol. Evol. 2005, 20, 420–421. [Google Scholar] [CrossRef]
- Dubey, S.C.; Singh, B. Integrated Management of major Diseases of Mungbean by Seed Treatment and Foliar Application of Insecticide, Fungicides and Bioagent. Crop Prot. 2013, 47, 55–60. [Google Scholar] [CrossRef]
- Zhong, S.; Miao, J.; Liu, X.; Zhang, G. Characterization of Colletotrichum spp. Sensitivity to Carbendazim for Isolates Causing Strawberry Anthracnose in China. Plant Dis. 2021, 105, 87–95. [Google Scholar] [CrossRef]
- Xue, Y.; Yang, Z.R.; Huang, X.B.; Xu, J.G. Properly handle the “3R” Problem to ensure the Quality and Safety of Agricultural Products. Hubei Prov. Plant Prot. 2017, 60–62. [Google Scholar]
- Usman, H.M.; Tan, Q.; Karim, M.M.; Adnan, M.; Yin, W.X.; Zhu, F.X.; Luo, C.X. Sensitivity of Colletotrichum fructicola and Colletotrichum siamense of Peach in China to Multiple Classes of Fungicides and Characterization of Pyraclostrobin-Resistant Isolates. Plant Dis. 2021, 105, 3459–3465. [Google Scholar] [CrossRef]
- Halko, R.; Sanz, C.P.; Ferrera, Z.S.; Rodriguez, J.J.S. Determination of Benzimidazole Fungicides in Soil Samples using microwave-assisted Micellar Extraction and Liquid Chromatography withFluorescence Detection. J. AOAC Int. 2006, 89, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Miller, M.G. The Role of the Benomyl Metabolite Carbendazim in benomyl-induced testicular Toxicity. Toxicol. Appl. Pharmacol. 1997, 142, 401–410. [Google Scholar] [CrossRef]
- Chechi, A.; Stahlecker, J.; Dowling, M.E.; Schnabel, G. Diversity in Species Composition and Fungicide Resistance Profiles in Colletotrichum Isolates from Apples. Pestic. Biochem. Physiol. 2019, 158, 18–24. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a Reduced Reliance on conventional Pesticides in European Agriculture. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef] [Green Version]
- Qu, T.; Gao, S.; Li, J.; Hao, J.J.; Ji, P. Synthesis and antifungal Activity of 2-allylphenol Derivatives against fungal Plant Pathogens. Pestic. Biochem. Physiol. 2017, 135, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Oufensou, S.; Scherm, B.; Pani, G.; Balmas, V.; Fabbri, D.; Dettori, M.A.; Carta, P.; Malbrán, I.; Migheli, Q.; Delogu, G. Honokiol, Magnolol and Its Monoacetyl Derivative show strong anti-fungal Effect on Fusarium Isolates of clinical Relevance. PLoS ONE 2019, 14, e0221249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Bi, Y.; Xue, H.; Wang, Y.; Zong, Y.; Prusky, D. Antifungal activity of cinnamaldehyde against Fusarium sambucinum involves Inhibition of Ergosterol Biosynthesis. J. Appl. Microbiol. 2020, 129, 256–265. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Sequence | Primer | Primer Sequence (5′→3′) |
|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG |
| ITS4 | TCCTCCGCTTATTGATATGC | |
| GAPDH [22] | GDR | GGGTGGAGTCGTACTTGAGCATGT |
| GDF | GCCGTCAACGACCCCTTCATTGA | |
| TUB2 [23] | Bt2a | GGTAACCAAATCGGTGCTGCTTTC |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC |
| Species | Strain Accession | GenBank Accession | ||
|---|---|---|---|---|
| ITS | GAPDH | TUB2 | ||
| C. fructicola C. cattleyicola | 022ZW CBS 17049 | OP523978 MG600758 | OP539309 MG600819 | OP539308 MG601025 |
| C. cliviicola | ST120 | MH291214 | MH291258 | MH458027 |
| C. navitas | CBS 125086 | JQ005769 | – | JQ005853 |
| C. sublineola | LZ-NX-1 | MK881657 | MK881674 | MK881725 |
| C. cliviicola | LZ-JY-1 | MK881658 | MK881675 | MK881726 |
| C. sublineola | LZ-GL-1 | MK881659 | MK881676 | MK881727 |
| C. fructicola | CBS 112.14 | KC566786 | KC566640 | KC566208 |
| C. fructicola | CBS 132461 | KC566784 | KC566638 | KC566206 |
| C. fructicola | CBS 197.34 | KC566789 | KC566643 | KC566211 |
| C. fructicola | CBS 132455 | KC566788 | KC566642 | KC566210 |
| C. cymbidiicola | CBS 128504 | JQ005238 | JQ005325 | JQ005672 |
| C. cymbidiicola | CBS 130241 | JQ005236 | JQ005323 | JQ005670 |
| C. hippeastri | CBS 125376 | JQ005231 | JQ005318 | JQ005665 |
| C. phyllanthi | CBS 175.67 | JQ005221 | JQ005308 | JQ005655 |
| C. torulosum | CBS 151.35 | GU227862 | GU228254 | GU228156 |
| C. parsonsiae | CBS 128525 | JQ005233 | JQ005320 | JQ005667 |
| Monilochaetes infuscans | CBS 869.96 | JQ005780 | JX546612 | JQ005864 |
| Phytochemicals | Concentration (µg/mL) | Regression Equation | EC50 (µg/mL) | Coefficient of Determination (R2) | 95% Confidence Interval |
|---|---|---|---|---|---|
| Honokiol | 400, 200, 100, 50, 25 | y = 2.9817 + 1.5100x | 21.70 ± 0.81 | 0.9904 | 13.68–34.44 |
| Magnolol | 300, 200, 100, 50, 25 | y = 2.5091 + 1.8038x | 24.04 ± 0.49 | 0.9980 | 15.41–37.48 |
| Carvacrol | 150, 100, 50, 20, 10 | y = 2.1503 + 1.9100x | 31.04 ± 0.89 | 0.9256 | 18.41–52.34 |
| Thymol | 500, 200, 100, 50, 25 | y = 2.6682 + 1.5495x | 31.97 ± 0.51 | 0.9744 | 17.64–57.96 |
| Citral | 500, 200, 100, 50, 25 | y = 1.0774 + 1.7119x | 41.17 ± 0.69 | 0.9129 | 28.53–89.76 |
| Citronellol | 60, 30, 20, 10, 5 | y = 1.9081 + 1.7525x | 58.11 ± 0.28 | 0.9835 | 36.09–93.56 |
| Geraniol | 150, 100, 50, 20, 10 | y = 3.4124 + 0.8930x | 59.96 ± 1.21 | 0.9887 | 25.31–142.01 |
| Eugenol | 400, 200, 100, 50, 25 | y = 3.1670 + 0.9392x | 89.47 ± 0.81 | 0.9835 | 18.77–426.31 |
| Citronellal | 80, 40, 20, 10, 5 | y = 1.3826 + 1.4634x | 296.39 ± 0.11 | 0.9349 | 133.49–658.07 |
| Cinnamaldehyde | 500, 200, 100, 50, 25 | y = 1.8266 + 1.1433x | 596.69 ±1.10 | 0.9639 | 214.57–1659.30 |
| Resveratrol | 500, 200, 100, 50, 25 | y = 2.9644 + 0.6950x | 849.37 ± 0.30 | 0.9930 | 637.04–1264.24 |
| Phytochemical Treatment | Year | |||||
|---|---|---|---|---|---|---|
| 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | |
| Sorghum Yield (kg/16 m2) | Severity (%) | Severity (%) | Control Percentage % | Control Percentage % | ||
| Honokiol | 13.33 ± 0.16 A | 14.05 ± 0.41 A | 6.61 F | 5.66 E | 70.45% | 74.54% |
| Magnolol | 13.41 ± 0.21 A | 14.19 ± 0.11 AB | 6.04 EF | 6.01 DE | 72.95% | 72.96% |
| Carvacrol | 12.44 ± 0.35 B | 13.03 ± 0.12 BC | 8.15 DE | 6.97 CED | 63.52% | 68.65% |
| Thymol | 12.23 ± 0.13 B | 12.83 ± 0.72 CD | 9.23 CD | 7.38 CD | 58.71% | 66.80% |
| Citral | 10.95 ± 0.17 C | 11.71 ± 0.22 DE | 9.68 CD | 7.99 C | 56.69% | 64.06% |
| Citronellol | 11.41 ± 0.37 C | 11.93 ± 0.35 CDE | 10.52 C | 7.51 CD | 52.91% | 55.83% |
| Geraniol | 11.02 ± 0.29 C | 11.54 ± 0.43 EF | 10.95 C | 7.56 CD | 51.02% | 53.71% |
| Eugenol | 10.05 ± 0.46 D | 10.51 ± 0.22 F | 16.13 B | 11.37 B | 27.82% | 36.93% |
| Control | 8.23 ± 0.29 E | 8.55 ± 0.21 G | 22.35 A | 22.23 A | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Hu, A.; Ren, M.; Wei, G.; Xu, H. First Report on Colletotrichum fructicola Causing Anthracnose in Chinese Sorghum and Its Management Using Phytochemicals. J. Fungi 2023, 9, 279. https://doi.org/10.3390/jof9020279
Zhao W, Hu A, Ren M, Wei G, Xu H. First Report on Colletotrichum fructicola Causing Anthracnose in Chinese Sorghum and Its Management Using Phytochemicals. Journal of Fungi. 2023; 9(2):279. https://doi.org/10.3390/jof9020279
Chicago/Turabian StyleZhao, Wei, Anlong Hu, Mingjian Ren, Guoyu Wei, and Huayang Xu. 2023. "First Report on Colletotrichum fructicola Causing Anthracnose in Chinese Sorghum and Its Management Using Phytochemicals" Journal of Fungi 9, no. 2: 279. https://doi.org/10.3390/jof9020279